
D.O. Okach
Department of Botany, Maseno University, P.O. Box 333, Maseno, Kenya
O. Amuka
Department of Applied Plant Science, Maseno University, Private Bag, Maseno, Kenya







ABSTRACT
Kisumu County has very minimal assessment of the local flora, with only scanty information from protected areas such as Ndere Island. There is no information on the roadside flora, yet road reserves are better conduit for biodiversity conservation due to limited human interference. This study assessed the role of a road reserve in conservation of floral diversity within Kisumu County. Species sampling was by transect method while percentage cover was by point quadrat method for grasses and herbs and total head count method for trees. Species identification was based on morphological attributes according to ICBN. A total of 133 plants were sampled from 45 families. The therophytes (19.8%) dominated in terms of cover, while, the microphanerophytes (18 species) dominated in species abundance. Fabaceae (20 species) had the highest number of species, though; Poaceae had the highest ground cover (15.7%).
PDF Abstract XML References Citation
Received: April 11, 2015;
Accepted: June 20, 2015;
Published: July 04, 2015
How to cite this article
D.O. Okach and O. Amuka, 2015. Plant Species Diversity along Road Reserve in Kisumu County- Kenya. Ecologia, 5: 8-21.
DOI: 10.3923/ecologia.2015.8.21
URL: https://scialert.net/abstract/?doi=ecologia.2015.8.21
DOI: 10.3923/ecologia.2015.8.21
URL: https://scialert.net/abstract/?doi=ecologia.2015.8.21
INTRODUCTION
The distribution and abundance of invasive species can be influenced by the presence of corridors and habitat suitability which facilitate dispersal of propagules. Species invasion usually result from environmental and anthropogenic factors such as disturbance, proximity to roads, nutrient availability, topography and habitat fragmentation (Brothers and Spingarn, 1992; Cadenasso and Pickett, 2001; Gelbard and Belnap, 2003). Roads also influence the spread and growth of species by serving as corridors for movement as well as providing habitat for establishment of propagules (Christen and Matlack, 2009). Propagules of invasive species may be accidentally spread during road construction and maintenance. Road graders and other maintenance equipments used during the operation can act like plows, pushing seeds along the road (Gelbard and Belnap, 2003; Ferguson et al., 2002; Hansen and Clevenger, 2005; Spooner, 2005). Some studies have also shown that plant propagules can be transported on vehicles (Forman et al., 2002; Pickering and Mount, 2010). Non native species are more likely to occur along roadsides and their probability of establishing in the interior is generally lower (Ferguson et al., 2002; Hansen and Clevenger, 2005). Culvert outwashes usually facilitate the movement of propagules off the road and roadways often serve as dispersal vectors within a region at both landscape and local scale (Flory and Clay, 2006).
Roadsides have different environmental conditions in terms of altered light availability, soil texture, compaction and chemistry; water regimes such as increased water runoff and repeated distribution for maintenance and off road driving when compared to adjacent native vegetation (Gelbard and Belnap, 2003; Hansen and Clevenger, 2005). Non native species may respond better than the indigenous species to the alien substrates altered disturbance, water and nutrient regimes found on roadsides. Upon their establishment on the roadside, they begin spreading interior over time (Pauchard and Alaback, 2004).
There are fewer publications in Kisumu County on assessment of the local flora (Agnew and Agnew, 1994; Arwa, 2005; Beentje, 1994; Kokwaro, 1993). There, however seems to be no information on floral diversity a long road reserves either protected or accidentally conserved. Most of the documented studies on species diversity available are on government protected sites such as Kakamega Forest and Mau Forest in the neighbouring Counties (KIFCON., 1994; Onyango et al., 2004). This study assessed the floral diversity along a road reserve within Kisumu County-Kenya as a way of recognizing their role in conservation of plant species. As a consequence, we developed a checklist on plant species diversity along Kisumu-Busia road reserve within Kisumu County with emphasis on plant habits, familial and species compositions.
MATERIALS AND METHODS
Study area: The study sites are located along the Kisumu-Busia Highway but within Kisumu County (0.1000° S, 34.7500° E) (Ojany and Ogendo, 1973) (Fig. 1), Kenya. The study was conducted in 7 blocks of land (sites) covering a total area of 60, 000 m2 along the highway from (00°06’00” S, 34°45’00” E) to (00°02’00” N, 34°35’00” E). The largest block was approximately 18, 000 m2, while the smallest was about 5, 000 m2. Since, all the study sites were on the road reserve (12 m from the highway), human activities were very limited due to security concerns and also given that all the reserves in Kenya are owned by the government. Consequently, the blocks represent abandoned pieces of land where only minimal cattle grazing and firewood collection take place occasionally. The study sites were characterized by large rock out-crops and road construction debris that supported plant growth. This gave the sites an elevation of approximately 30 m above the highway level (1131-1501 m) from Kisumu town to the County Boundary Northwards.
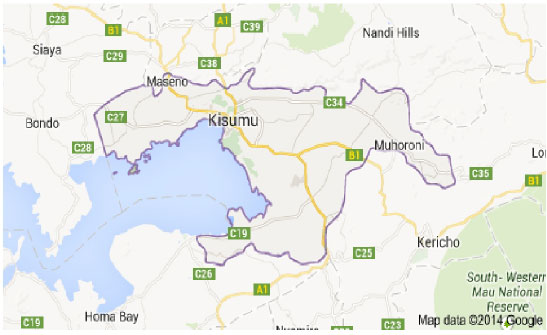 | |
| Fig. 1: | Map of Kasumu country showing the road under study (marked B1) between Kisumu and Maseno |
The soils were sandy with overlapping ferralsols from Mount Elgon eruptions in certain parts. The sites were rich in floral composition, what could likely be attributed to the limited human access to the areas prompted by their elevation, thus the accidental conservation. The study sites were commonly dominated by plant species such as Tithonia diversifolia (Helmsl) A. Gray and Calamintha nepeta (L) Savi by the road side and tree species such as Albizia coriara Welw. Ex. Oliv. and Markhamia lutea (Benth.) K. Schuman at the elevation peaks. The rock surfaces and tree barks were covered by species of moss such as: Campylopus arctocarpus (Hornsch.) Mitt, Epipordium becarii (Mull. Hal). Ex Venturi and Rhynchostegium comorae (Mull. Hal.) A. Jaeger among others. The areas adjacent to both roads were dominated by invasive species such as Ipomoea hildelbrandtii Vatke, Calamintha nepeta (L) Savi, Tithonia diversifolia (Helmsl) A. Gray and Lantana camara L. which could have developed as a result of accidental movement of plant propagules by road construction machines (Christen and Matlack, 2009).
Sampling method: The sites were stratified into three categories (1) Foot hill, (2) Escarpment and (3) Hill top. Sampling for plant species diversity was by transect method while percentage cover was done using point frame quadrat method for grasses and herbaceous species. Total head count method was also used to estimate percentage cover for trees. Five transects, 2 at the foot hill, 1 at the escarpment and 2 at the hill top, each measuring 2×20 m were randomly laid in each block along the highway to estimate species diversity. The invasive species were identified according to the (GISIN., 2008) invasive species list. Plant life form spectrum analysis was based on and (Govaerts et al., 2000) systems. Data on percentage composition was collected using 5 quadrats measuring 2×2 m from all transects within a block. Samples were identified based on their morphological attributes according to ICBN and then deposited to the East Africa Herbarium, in The National Museum of Kenya.
RESULTS
From the study, a total of 133 plant species were sampled belonging to 45 different plant families (Table 1). Most species were erect while some were climbers and scramblers with a few lianas. Twenty invasive species were reported from the study as the rest (93) were non invasive (Table 1). The ten most species rich families included: Fabaceae (20 species), Euphorbiaceae (10 species), Asteraceae (9 species), Malvaceae (9 species), Verbenaceae (8 species), Rubiaceae (7 species), Lamiaceae (5 species), Poaceae (5 species), Vitaceae (5 species) and Apocynaceae (5 species) (Table 2). Families such as Acanthaceae, Cyperaceae, Moraceae and Solanaceae among others (Table 2) reported one species each and were considered as the least species rich families. The families with the highest percentage ground cover included; Poaceae (15.7%), Asteraceae (10.087%), verbenaceae (9.389%) and Lamiaceae (9.2%) as shown in Table 2. Families such as, Orchidaceae (0.139%) and Ranunculaceae (0.139%) recorded the lowest percentage ground cover (Table 2). In terms of the life forms, the therophytes (19.8%) had the highest percentage composition followed by the microphanerophytes (18.8%), nanophanerophytes (18.4%), while, the epiphytes (0.1%) recorded the lowest composition (Fig. 2). On the other hand, the microphanerophytes had the largest number of families (17 families), followed by the mesophanerophytes (13 families), the epiphytes (1) had the least number of species (Fig. 3).
| Table 1: | Checklist of plant species sampled |
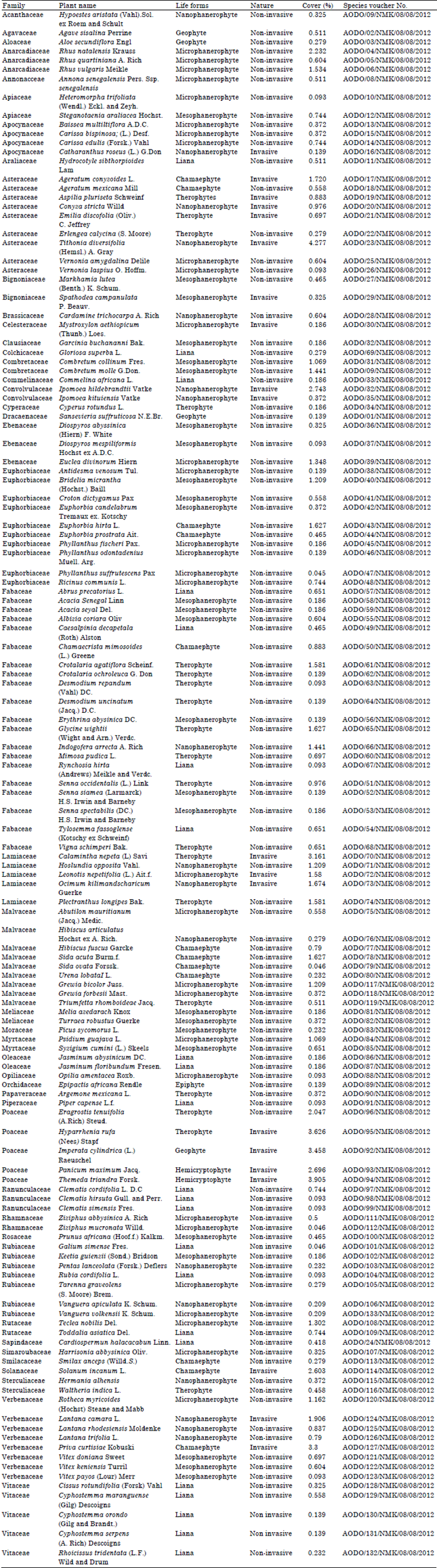 | |
The hemicryptophytes (1 family) and epiphytes (1 family) recorded the lowest number of families in their categories (Fig. 3). The ten most dominant plant species in the area included: Tithonia diversifolia (Hemsl.) A. Gray (4.28%), Themeda triandra Forsk (3.9%), Hyparrhenia rufa (Nees) Stapf (3.63%), Imperata cylindrica (L) Raeuschel (3.5%), Priva curtisiae Kobuski (3.3%), Calamintha nepeta (L) Savi (3.2%), Ipomea hidelbrandtii Vatke (2.7%), Panicum maximum Jacq. (2.7%), Solanum incanum L. (2.6%) and Rhus natalensis Kraus (2.2%) (Table 1). The distribution of Tithonia diversifolia, Priva curtisiae and Calamintha nepeta and some other highly invasive species (Table 1) decreased from the road.
| Table 2: | Species composition per plant family |
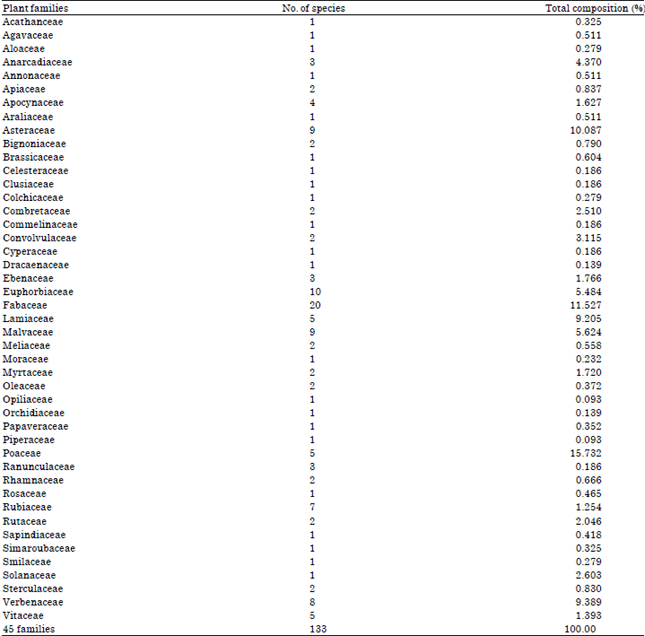 | |
DISCUSSION
Most plant species dominating the roadside were found to be invasive and there was a high likelihood that their distribution could have resulted from road construction. This is evidenced by the fact that away from the road, the number of species declined. Such a finding has been reported in some parts of the world where it was revealed that most species found along roads were invasive and mostly distributed seeds, runners and rhizomes (Forman and Alexander, 1998) transported by road construction machines (Forman et al., 2002; Pickering and Mount, 2010; Pauchard and Alaback, 2004). Tithonia diversifolia, Priva curtisiae and Calamintha nepeta (Table 1) were the most dominant species along Kisumu-Busia road with relatively higher percentage composition. These plant species were persistent on the road reserve for quite a long time and attempts to maintain the road failed to eliminate them completely.
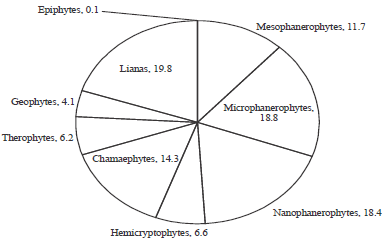 | |
| Fig. 2: | Life form spectrum analysis of plant species (in percentage) found in a road reserve within Kisumu country |
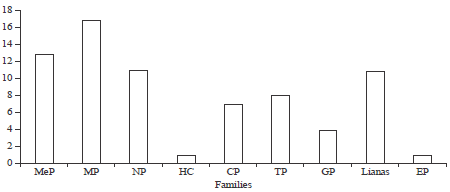 | |
| Fig. 3: | Familial composition of plant life forms in a road reserve within in Kisumu country, MeP: Mesophanerophytes, MP: Microphanerophytes, NP: Nanophanerophytes, HC: Hemicryptophytes, Cp: Chamaephytes, TP: Therophytes, GP: Geophytes, Ep: Epiphytes |
Thus, the main reason they flourish in spite of continuous clipping by construction machines during maintenance. Similar findings have also been reported in Tanzania (Mollel et al., 2012) and other parts of the world where the above plants among other invasive species have dominated roadsides (Forman et al., 2002; Holway et al., 2002; Moktan and Das, 2013). Besides the road construction machines, these plants have also been reported to have a facilitated means of spreading their propagules such as seeds with a lot of ease (Pauchard and Alaback, 2004). The therophytes (19.8%), microphanerophytes (18.8%), nanophanerophytes (18.4%) and chamaephytes (14.3%) were the most dominant life forms recorded from the study area. However, in terms of species composition, the microphanerophytes (17 species), were the most dominant followed by mesophanerophytes (13 species), nanophanerophytes (11 species) and therophytes (8 species) among others (Fig. 3). The therophytes mostly constituted the annual herbaceous species that bore seeds and were very prolific in their reproduction. The microphanerophytes and nanophanerophytes delimited the herbs and shrubs based on their heights ranging from 30-8 m. The chamaephytes included the lowly herbaceous species with perrenating buds located closer to the ground surface (less than 30 cm). Several studies on biodiversity have reported different findings on the most dominant life forms, based on the classification system used climatic conditions of the area and the nature of the ecosystem under study. For instance, most tropical ecosystems (Batalha and Martins, 2004; Koulibaly et al., 2006; Ouedraogo et al., 2011; Schmidt et al., 2005) have ascertained that the phanerophytes are the most dominant life form especially in the forested regions. In most savannas ecosystems however, the therophytes (Aghaei et al., 2013; Medvecka et al., 2013; Turki and El Shayeb, 2005), chamaephytes (Gimenez et al., 2004) and hemicryptophytes (Amjad, 2012; Dimopoulos and Georgiadis, 1992; Klimes, 2003) have been reported to dominate different regions depending on how well they interact with their environment (Zarezade et al., 2007). From our study, phanerophytes grouped together constituted 48.9%, which implied that they were the most dominant, however, there was need to delimit the groups (Govaerts et al., 2000) for ease of understanding, given the floral diversity of the study area.
The therophytes, microphanerophytes and nanophanerophytes that constituted over 50% of the total composition, were mostly herbaceous species and were sampled closer (within 3 m from the edge of the road) to the road as compared to other life forms such as mesophanerophytes. Such kind of life form distribution could be attributed to the routine road construction that takes about two years, a period that cannot allow the trees and higher shrubs to be fully established. This has also been reported in other studies where the shrubs (mostly nanophanerophytes and microphanerophytes) have been the most dominant plant form in terms of species diversity (Gautam et al., 2014; Schnitzer and Bongers, 2002; Schnitzer and Carson, 2000) especially in disturbed areas. The frequent disturbance on the roadside resulting from machines and pedestrians could have limited the distribution of most trees and shrubs to the elevated land. Elsewhere (Trombulak and Frissell, 2000; Gelbard and Belnap, 2003), it has been reported that clearing vegetation during road construction, addition of road fill and grading of unpaved roads have created areas of bare and deeper soils that allow exotic seeds of grasses, herbs and shrubs to be established (Cadenasso and Pickett, 2001; Gelbard and Belnap, 2003). The ease in dispersal of the propagules was attributed to the short life cycle of the plant forms and their invasive nature resulting from their adaptability to various soil types and production of high seed capacity (Williamson and Fitter, 1996; Zavaleta et al., 2001).
The 10 most dominant plant species colonizing the area (Table 1) were drawn from all life forms except the lianas, epiphytes and mesophanerophytes and the interesting feature is that they were mostly invasive. Rhus natalensis, even though dominant in the study area, is mostly distributed far distances from the edge of the road and this implies that the dispersal of its propagules could result from natural agents rather than those acting in the area and not necessarily road construction. Tithonia diversifolia, was the most dominant plant species in the study area (Table 1) and this could be attributed to its adaptive features that enables it to propagate both through seeds and also by vegetative means, hence the movement of its roots and other plant parts could lead to further regeneration of a whole plant (Oludare and Muoghalu, 2014; Wang et al., 2003). As a result of this adaptation, the plant was mostly found adjacent to the road as reported elsewhere (Chukwuka et al., 2007; Mollel et al., 2012; Wang et al., 2003) and not remotely within the road reserve. Fabaceae was the most dominant family with 20 plant species most of which were the therophytes and mesophanerophytes (Table 1 and 2), yet in terms of percentage composition, Poaceae was the most dominant (15.73%), in spite of its lower species abundance (5 plant species) (Table 1 and 2). The same trend was also observed in other plant families such as Euphorbiaceae (10 species), Asteraceae (9 species), Rubiaceae (7 species) and Verbenaceae (8 species) which were all more diverse (Table 2) compared to Poaceae family. The grasses constituted the highest ground cover (Table 1) and were distributed evenly which implied that they had well developed reproductive mechanisms that enabled them to propagate with a lot of ease within the ecosystem. These findings are in conformity with some studies conducted in Kenya and East African savannas and ranches on species diversity that revealed that grasses constitute the greatest percentage cover of floral diversity (Vanacker et al., 2005) due to their sensitivity to rainfall. Besides, some findings have revealed that Imperata cylindrica and Themeda triandra have higher percentage cover along highways and power lines (Goosen and Turton, 2006), what was attributed to the viability of their seeds and ease to disperse through wind (Cheplick, 2009). Parallel studies conducted elsewhere in various parts of Africa and globally (Mbayngone et al., 2008; Ouedraogo et al., 2011; Schmidt et al., 2007) have also shown that the family Fabaceae, Poaceae, Asteraceae, Cyperaceae and Eurphobiaceae are some of the most dominant (Schmidt et al. 2007), though depending on the climatic conditions, either Poaceae (Kabelo and Mafokate, 2004; Klaasen and Craven, 2003; Singh and Singh, 2014) or Fabaceae (Mbayngone et al., 2008; Ouedraogo et al., 2011; Schmidt et al., 2005; Schmidt et al., 2010) could be the most diverse. The diversity of the former, though accompanied with dominance of percentage cover is usually dependent on edaphic factors such as soil fertility and topography (Dukes, 2001). Research conducted on roadside plant species diversity in Arusha, Tanzania, revealed that family Asteraceae dominated the roadside vegetation followed by Fabaceae, Poaceae and Euphorbiaceae (Mollel et al., 2012).
Some studies conducted in Africa and other parts of the world have also confirmed that plant families such as Rubiaceae, Malvaceae and Lamiaceae also constitute a larger percentage of species in most ecosystem (Gaston, 2000; Mutke and Barthlott, 2005; Onyango et al., 2004; Schmidt et al., 2010). Other studies (Fridley, 2001; Johnson et al., 2008; Schwartz et al., 2000) have revealed that species diversity and percentage cover can differ depending on the ability of individual plant species to disperse and adapt to their surrounding environment as a result of varying edaphic factors. Further revelations have indicated that species composition and that of relatively few species is likely the best predictor of resource partitioning among plant species and not the number of local species (Johnson et al., 2008; Schwartz et al., 2000; Tilman et al., 1997). From the findings of this study, it was quite clear that the vegetation by the roadside did not vary much from that of the adjacent areas (mostly native) except for some invasive shrubs and herbs such as Ageratum conyzoides, Mystroxylon aethiopicum and Ipomoea kituiensis among others. The propagules of these species were highly likely to have been moved from other areas by road construction machines, thus limiting their distribution by the road side (Forman et al., 2002; Pickering and Mount, 2010). The slight difference in species composition could also be attributed to the variation in environmental condition and factors such as soil texture and compaction (Gelbard and Belnap, 2003) which might have hindered their spread remotely. This pattern in distribution of species by the roadside has been reported by similar studies conducted in other parts of the globe (Ferguson et al., 2002; Hansen and Clevenger, 2005). This trend if maintained through further protection, will greatly aid in the maintenance of taxa of conservation concern, thus increasing floral diversity within the country at large. This is possible since, the roads provide adequate landscape linkages that can be used in conservation networks.
CONCLUSION
Road reserves being corridors for conservation of species diversity should be given special consideration. This comes at a time when biodiversity loss has drawn the attention of very many conservationists especially in the areas perceived to be protected such as game reserves and parks whose accessibility are usually restricted. Plant species a long road reserves should be included in the National Biodiversity Conservation Strategies as a way of maintaining floral diversity within Kenya and Kisumu County. This way they will contribute valuable data that can supplement the currently available information on species diversity and also help to conserve some plant species that are considered threatened in the region. Besides, such measures will be essential in both current and future floral conservation.
REFERENCES
- Aghaei, R., S. Alvaninejad, R. Zolfaghari and M.R.M. Gharehlar, 2013. Flora, life form and geographical distribution of plants in West-South forests of Iran (Case study: Vezg, Yasouj). Int. J. Farming Allied Sci., 2: 1046-1053.
Direct Link - Amjad, M.S., 2012. Life form and leaf size spectra of vegetation in Kotli Hills, Azad Jammu and Kashmir (Pakistan). Greener J. Agric. Sci., 2: 345-350.
Direct Link - Batalha, M.A. and F.R. Martins, 2004. Floristic, frequency and vegetation Life-form spectra of a cerrado site. Braz. J. Biol., 64: 203-209.
CrossRefDirect Link - Brothers, T.S. and A. Spingarn, 1992. Forest fragmentation and alien plant invasion of central Indiana Old-growth forests. Conserv. Biol., 6: 91-100.
CrossRefDirect Link - Cadenasso, M.L. and S.T.A. Pickett, 2001. Effect of edge structure on the flux of species into forest interiors. Conserv. Biol., 15: 91-97.
CrossRefDirect Link - Christen, D.C. and G.R. Matlack, 2009. The habitat and conduit functions of roads in the spread of three invasive plant species. Biol. Invasions, 11: 453-465.
CrossRefDirect Link - Chukwuka, K.S., S. Ogunyemi and I. Fawole, 2007. Ecological distribution of Tithonia diversifolia (Hemsl). A. Gray-a new exotic weed in Nigeria. J. Biol. Sci., 7: 709-719.
CrossRefDirect Link - Dimopoulos, P. and T. Georgiadis, 1992. Floristic and phytogeographical analysis of mount killini (NE Peloponnisos, Greece). Phyton, 32: 283-305.
Direct Link - Dukes, J.S., 2001. Productivity and complementarity in grassland microcosms of varying diversity. Oikos, 94: 468-480.
Direct Link - Flory, S.L. and K. Clay, 2006. Invasive shrub distribution varies with distance to roads and stand age in eastern deciduous forests in Indiana, USA. Plant Ecol., 184: 131-141.
CrossRefDirect Link - Gaston, K.J., 2000. Biodiversity: Higher taxon richness. Progress. Phys. Geogr., 24: 117-128.
CrossRefDirect Link - Gautam, M.K., R.K. Manhas and A.K. Tripathi, 2014. Plant species diversity in unmanaged deciduous forests in Northern India. Curr. Sci., 106: 277-287.
Direct Link - Gelbard, J.L. and J. Belnap, 2003. Roads as conduits for exotic plant invasions in a semiarid landscape. Conserv. Biol., 17: 420-432.
CrossRefDirect Link - Gimenez, E., M. Melendo, F. Valle, F. Gomez-Mercado and E. Cano, 2004. Endemic flora biodiversity in the south of the Iberian Peninsula: Altitudinal distribution, life forms and dispersal modes. Biodiver. Conserv., 13: 2641-2660.
CrossRefDirect Link - Hansen, M.J. and A.P. Clevenger, 2005. The influence of disturbance and habitat on the presence of non-native plant species along transport corridors. Biol. Conserv., 125: 249-259.
CrossRefDirect Link - Fridley, J.D., 2001. The influence of species diversity on ecosystem productivity: How, where and why? Oikos, 93: 514-526.
CrossRefDirect Link - Johnson, D., G.K. Phoenix and J.P. Grime, 2008. Plant community composition, not diversity, regulates soil respiration in grasslands. Biol. Lett., 4: 345-348.
CrossRefDirect Link - Klimes, L., 2003. Life-forms and clonality of vascular plants along an altitudinal gradient in E Ladakh (NW Himalayas). Basic Applied Ecol., 4: 317-328.
CrossRefDirect Link - Koulibaly, A., D. Goetze, D. Traore and S. Porembski, 2006. Protected versus exploited savannas: Characteristics of the sudanian vegetation in ivory coast. Candollea, 61: 425-452.
Direct Link - Singh, D.K. and R. Singh, 2014. Study of angiospermic wall floristic composition of city Buxar (Bihar) India. J. Pharmacogn. Phytochem., 2: 52-54.
Direct Link - Mbayngone, E., M. Schmidt, K. Hahn-Hadjali, A. Thiombiano and S. Guinko, 2008. Magnoliophyta of the partial faunal reserve of Pama, Burkina Faso. Check List, 4: 251-266.
Direct Link - Moktan, S. and A.P. Das, 2013. Diversity and distribution of invasive alien plants along the altitudinal gradient in Darjiling Himalaya, India. Pleione, 7: 305-313.
Direct Link - Mutke, J. and W. Barthlott, 2005. Patterns of vascular plant diversity at continental to global scales. Biol. Skr., 55: 521-537.
Direct Link - Ouedraogo, O., M. Schmidt, A. Thiombiano, K. Hahn, S. Guinko and G. Zizka, 2011. Magnoliophyta, Arly National Park, Tapoa, Burkina Faso. Check List, 7: 85-100.
Direct Link - Oludare, A. and J.I. Muoghalu, 2014. Impact of Tithonia diversifolia (Hemsly) A. Gray on the soil, species diversity and composition of vegetation in Ile-Ife (Southwestern Nigeria), Nigeria. Int. J. Biodivers. Conserv., 6: 555-562.
Direct Link - Onyango, J.C., R.A.O. Nyanja and R.W. Bussmann, 2004. Conservation of biodiversity in the East African tropical forest. Lyonia J. Ecol. Applied, 7: 151-157.
Direct Link - Pauchard, A. and P.B. Alaback, 2004. Influence of elevation, land use and landscape context on patterns of alien plant invasions along roadsides in protected areas of South-Central Chile. Conserv. Biol., 18: 238-248.
CrossRefDirect Link - Pickering, C. and A. Mount, 2010. Do tourists disperse weed seed? A global review of unintentional Human-mediated terrestrial seed dispersal on clothing, vehicles and horses. J. Sustainable Tourism, 18: 239-256.
CrossRefDirect Link - Forman, R.T.T. and L.E. Alexander, 1998. Roads and their major ecological effects. Ann. Rev. Ecol. Syst., 29: 207-231.
CrossRefDirect Link - Schmidt, M., H. Kreft, A. Thiombiano and G. Zizka, 2005. Herbarium collections and field data-based plant diversity maps for Burkina Faso. Divers. Distrib., 11: 509-516.
CrossRefDirect Link - Schmidt, M., H. Kreft, A. Thiombiano and G. Zizka, 2007. Indicator value of different vascular plant families for overall plant diversity in the savannah belt of Western Africa (Abridged title: Grasses as indicators of plant iversity). Ecotropica, 13: 135-142.
Direct Link - Schnitzer, S.A. and F. Bongers, 2002. The ecology of lianas and their role in forests. Trends Ecol. Evol., 17: 223-230.
CrossRefDirect Link - Schnitzer, S.A. and W.P. Carson, 2000. Have we forgotten the forest because of the trees? Trends Ecol. Evol., 15: 375-376.
CrossRefPubMedDirect Link - Schwartz, M.W., C.A. Brigham, J.D. Hoeksema, K.G. Lyons, M.H. Mills and P.J. van Mantgem, 2000. Linking biodiversity to ecosystem function: Implications for conservation ecology. Oecogia, 122: 297-305.
Direct Link - Spooner, P.G., 2005. Response of Acacia species to disturbance by roadworks in roadside environments in southern New South Wales, Australia. Biol. Conserv., 122: 231-242.
CrossRefDirect Link - Tilman, D., J. Knops, D. Wedin, P. Reich, M. Ritchie and E. Siemann, 1997. The influence of functional diversity and composition on ecosystem processes. Science, 277: 1300-1302.
CrossRefDirect Link - Trombulak, S.C. and C.A. Frissell, 2000. Review of ecological effects of roads on terrestrial and aquatic communities. Conserv. Biol., 14: 18-30.
CrossRefDirect Link - Turki, Z.A. and F.M. El Shayeb, 2005. Floristic and ecological studies of the western Mediterranean Coastal land in Egypt (Wadi um Rakham). Flora Mediterraea, 15: 331-341.
Direct Link - Vanacker, V., M. Linderman, F. Lupo, S. Flasse and E. Lambin, 2005. Impact of short-term rainfall fluctuation on interannual land cover change in Sub-Saharan Africa. Global Ecol. Biogeogr., 14: 123-135.
CrossRefDirect Link - Wang, S., W. Sun and X. Cheng, 2003. Attributes of plant proliferation, geographic spread and the natural communities invaded by the naturalized alien plant species Tithonia diversifolia in Yunnan, China. Acta Ecologica Sinica, 24: 444-449.
PubMedDirect Link - Williamson, M.H. and A. Fitter, 1996. The characters of successful invaders. Biol. Conserv., 78: 163-170.
CrossRefDirect Link - Zavaleta, E.S., R.J. Hobbs and H.A. Mooney, 2001. Viewing invasive species removal in a whole-ecosystem context. Trends Ecol. Evolut., 16: 454-459.
CrossRefDirect Link - Holway, D.A., L. Lach, A.V. Suarez, N.D. Tsutsui and T.J. Case, 2002. The causes and consequences of ant invasions. Annu. Rev. Ecol. Syst., 33: 181-233.
CrossRefDirect Link