
Neda Fazeli
Department of Biology, Khorramshahr University of Marine Science and Technology, Khorramshahr, Iran
Rasool Zare
Department of Biology, Khorramshahr University of Marine Science and Technology, Khorramshahr, Iran







ABSTRACT
The distribution and abundance of female and male of Paracalanidae presented within the near shore and off shore waters of Chabahar Bay. Paracalanidae conducted from zooplankton sampling during four oceanography cruises August 2007 (SW-monsoon), November 2007 (post-monsoon), February 2008 (NE-monsoon) and May 2008 (pre-monsoon). Five stations were investigated throughout the Bay. Paracalanidae represented by Acrocalanus (A. longicornis, A. gracilis, A. gibber, A. monachus and Acrocalanus sp.) and Paracalanus (P. aculeatus, P. denudatus, P. parvus, P. crassirostris, P. elegans and P. indicus). Two-way ANOVA showed that difference in Paracalanidae abundance amongst periods and locations was significant (p<0.05). Our result showed that environmental parameters (temperature, salinity, chlorophyll-a, pH, DO and depth) influence on the abundance and distribution of both sexes of Paracalanidae species. Paracalanidae female always outnumbered than males. The reason could be that copepod adult males live for shorter periods than females. Also sex change could be in some Paracalanidae species from copepodite stage 5 male to female.
PDF Abstract XML References Citation
Received: November 05, 2011;
Accepted: June 15, 2012;
Published: December 19, 2012
How to cite this article
Neda Fazeli and Rasool Zare, 2012. The Seasonal Change of Female and Male Paracalanidae (Calanoid Copepod) in Chabahar Bay: The Gulf of Oman. Ecologia, 2: 93-103.
DOI: 10.3923/ecologia.2012.93.103
URL: https://scialert.net/abstract/?doi=ecologia.2012.93.103
DOI: 10.3923/ecologia.2012.93.103
URL: https://scialert.net/abstract/?doi=ecologia.2012.93.103
INTRODUCTION
In copepod, males and females vary in the stage duration and males live for shorter periods than females (Uye et al., 1983). Seasonal variation in environment effects on adult sex ratio skew and also change abundance of sexes of juvenile stages (Hirst et al., 2010). It was stated males tend to mature faster than females and a population in growth phase will have more males arriving at the adult stage which will push the sex’s abundance toward male dominance (Hirst et al., 2010). Once recruitment has s topped the adult abundance may then swing to females if these have a greater longevity, if they arrive later given their slower development rates, or both (Kiorboe, 2006). Environmental factors like nutrition, temperature, pressure, population density and parasitism as possible inducers effect on copepod female and male abundance (Takeda, 1950; Katona, 1970) the environment can alters the male and female abundance (Irigoien et al., 2000). Studies on copepod development and ultimately, their productivity are amongst some of the best ways to quantify and understand these fluxes and therefore have become more common in recent marine zooplankton research (Runge and Roff, 2000).
Importance of major parameters on female and male copepod abundance is known well but there is not published information on effect of environmental parameters on abundance of zooplankton sexes in Chabahar Bay. However, there are many studies have described effect of various factors on zooplankton (Javed, 1999; Leghari et al., 2001; Jahangir et al., 2001; Tarkan et al., 2005; Ahmed et al., 2007; El-Sherbiny et al., 2007; Oueda et al., 2007; Rahimibashar et al., 2009; Kumar and Perumal, 2011) and abundance of female and male (Takeda, 1950; Katona, 1970).
To gain a better understanding on the role and importance of copepods in structuring pelagic food webs, their biology and life history traits are required to compare between season and stations. The present study is based on the examination of female and male Praracalanidae species abundance and effect of environmental parameters on it.
MATERIALS AND METHODS
Chabahar Bay is a small semi-enclosed and sub-tropical Bay on the southeastern coasts of Iran (from 25°17' 45"N-60°37' 45" E). The Bay surface area is 290 km2 with 14 km wide located between of Chabahar and Konarak. The year is divided into periods of northeast (NE) monsoon and southwest (SW) monsoon and its following inter-monsoon periods (post-monsoon) and (pre-monsoon). This Bay is one of the five major ports in the Arabian Sea and is located in naturally suitable fish sites (Wilson, 2000). The samples conducted from zooplankton during four oceanography cruises August 2007 (SW-monsoon), November 2007 (post-monsoon), February 2008 (NE-monsoon) and May 2008 (pre-monsoon). Five stations were investigated throughout the Bay. Two stations (St 1 and 2) were located far from shore waters with 22 m depth, another two stations were the near shore with 6 m depth (St. 3 and 5) and the final station (St. 4) was located in the middle of the Bay with 12 m depth (Fig. 1). Samples were typically collected by vertical hauls (with 100 μm mesh nets) from bottom to the sea surface. Plankton samples fixed immediately in 4-5% formalin, buffered to a pH of 8 with sodium tetraborate (borax). Adult Paracalanidae counts were made under a MBS-9 stereomicroscope in Bogorov counting chamber. Sex and species of samples were identified using the taxonomic keys of Davis (1955), Krishnapillai (1986) and Conway et al. (2003).
 | |
| Fig. 1: | Map of study area, St: Sampling stations 1-5 |
Statistical analysis: We used SPPS (version 12.0) to analyze the data. Two-way ANOVA was used to assess the significant of difference in abundance amongst periods and locations. The Pearson correlation and multivariate regression analyses were performed to determine the significance between environmental parameters and Paracalanidae abundance.
RESULTS AND DISCUSSION
Environmental variables: The annual water temperature variation ranged from an average (av.) of 21.33±0.13°C in NE-monsoon to an av. of 29.92±0.45°C in SW-monsoon; whereas salinity ranged from an av. of 36.66±0.03 during the NE-monsoon to an av. of 36.91±0.14 in pre-monsoon (Fig. 2). The average chlorophyll-a concentrations ranged from 0.77±0.08 mg m-3 in SW-monsoon to 1.94±0.92 mg m-3 in NE-monsoon. pH values showed a small variation during four surveys ranging from 8.29±0.04 in NE-monsoon to 8.21 in SW-monsoon. Minimum and maximum values of Dissolved Oxygen (DO) water were from 5.66±0.05 ml L-1 in SW-monsoon to 8.80±0.61 ml L-1 in NE-monsoon.
Temporal and spatial variation of female and male Acrocalanus abundance: Seasonally average abundance of female and male Acrocalanus varied considerably. A significant higher female and male abundance of A. longicornis (209 and 20 ind m-3), A. gibber (167 and 20 ind m-3), A. gracilis (177 and 20 ind m-3) and A. monachus (63 and 91 ind m-3) was found during SW-monsoon (p<0.05; Fig. 3). Male of A. longicornis, A. gibber and A. gracilis disappeared during post, NE and pre-monsoon.
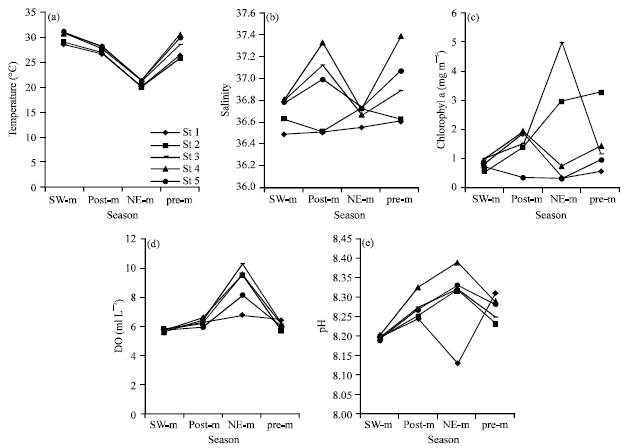 | |
| Fig. 2(a-e): | Distribution of major physicochemical variables during monsoonal seasons, SW.m: SW-monsoon, post-m: Post-monsoon, NE-m: NE-monsoon and pre-m: Pre-monsoon |
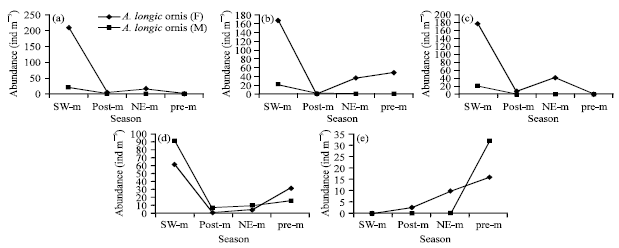 | |
| Fig. 3(a-e): | Abundance of female (F) and male (M) Acrocalanus (ind m-3) in monsoonal seasons, SW.m: SW-monsoon, post-m: Post-monsoon, NE-m: NE-monsoon and pre-m: Pre-monsoon |
Male and female abundance of A. sp. observed rarely showing highest of female and male in pre-monsoon (16 and 30 ind m-3). Female disappeared in SW-monsoon and male disappeared in SW, post and NE-monsoon. Some studies in India showed large numbers of adult males and females of A. gibber in the months of December-January and A. gracilis and A. monachus in January and April (Ummerkutty, 1965).
Female of A. longicornis was absent in station 3 while male was absent in stations 1, 3 and 4. A. gracilis female was not in station 5 and male disappeared in stations 1, 3 and 4. A. gibber female showed highest (average (av.) 73 ind m-3) in station 5 and lowest in station 2 (av. 32 ind m-3). Male disappeared in stations 4 and 5. Male of A. monachus showed significant increase in station 1 (av. 82 ind m-3) and was absent at station 5 during four seasons. Female was highest in station 5 (av.46 ind m-3).
Male of A. sp. observed only at station 1 (160 ind m-3) during pre-monsoon (Fig. 4) (McKinnon, 1996; McKinnon and Ayukai, 1996; Araujo, 2006) stated Acrocalanus are often the most common copepods in tropical and subtropical coastal waters.
Temporal and spatial variation of female and male Paracalanus abundance: Paracalanus represented by P. aculeatus, P. denudatus, P. parvus, P. crassirostris, P. elegans and P. indicus in Chabahar Bay. A significantly higher female and male abundance of P. aculeatus (132 and 112 ind m-3) observed in SW-monsoon (p<0.05). P. aculeatus female showed minimum abundance in NE-monsoon (18 ind m-3) and male was entirely absent in NE-monsoon and pre-monsoon (Fig. 5). This species was found at its maximum during January-March in Gulf of Mannar, India (Ummerkutty, 1965). Female and male abundance of P. crassirostris were higher significantly in post-monsoon (163 and 44 ind m-3). Male was absent in NE-monsoon and pre-monsoon. Araujo (2006) observed low abundance of P. crassirostris in the whole year in Brazil, however highest abundance was observed in dry seasons. The maximum of abundance P. elegans female was in post monsoon (308 ind m-3), whereas male observed highest in SW-monsoon (96 ind m-3). P. denudatus showed low abundances during four periods. Female showed highest abundance in SW-monsoon (48 ind m-3) and completely was absent in pre monsoon. Male showed highest abundance in post monsoon (8 ind m-3).
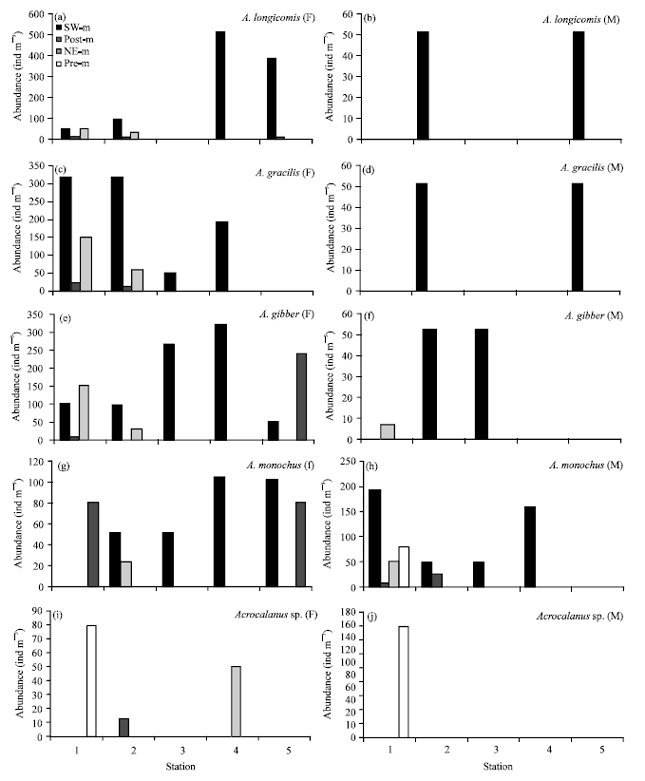 | |
| Fig. 4(a-j): | Abundance of female (F) and male (M) Acrocalanus in Chabahar Bay stations, SW.m: SW-monsoon, post-m: Post-monsoon, NE-m: NE-monsoon and pre-m: Pre-monsoon |
Female and male of P. parvus showed low abundance during four periods. Female showed highest abundance in NE-monsoon (21 ind m-3). Male was almost entirely absent or present only in small numbers. P. parvus is known to be widely distributed in most of the world's oceans (Peterson, 1998). This species showed maximum abundance in February (Tseng et al., 2008) and in spring (Hsieh et al., 2005) in Taiwan Strait.
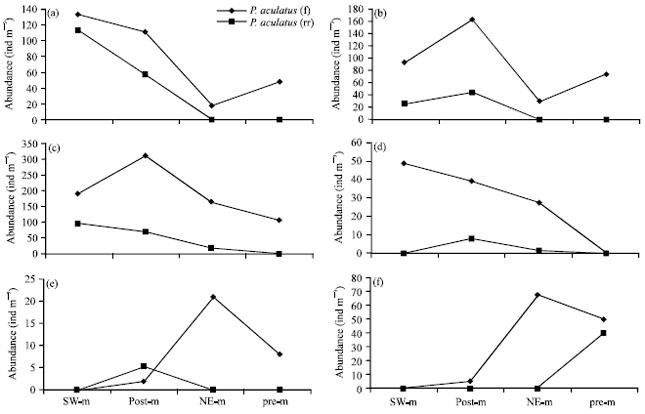 | |
| Fig. 5(a-f): | Abundance of female (F) and male (M) Paracalanus in seasons, SW.m: SW-monsoon, post-m: Post-monsoon, NE-m: NE-monsoon and pre-m: Pre-monsoon |
This species is common in waters of southern China during winter (Hwang and Wong, 2005). Male and female of P. indicus was observed in low number. This species showed highest abundance of female in NE-monsoon (68 ind m-3). Male was observed just in pre-monsoon (40 ind m-3). P. indicus was more abundant in the rainy months in Brazil (Araujo, 2006).
Spatially, female and male of P. aculeatus showed higher abundance at station 1 (av.133 and av.113 ind m-3) (Fig. 6). This species showed maximum abundance in coastal waters off China (Chen and Zhang, 1974). Female of P. crassirostris showed highest abundance in station 1 (av.203 ind m-3) whereas male showed highest in station 2 (av.46 ind m-3). Female of P. elegans observed highest at station 2 (av.330 ind m-3) whereas male was highest in station 1 (av.127 ind m-3). Abundance of female P. denudatus showed highest abundance in station 2 (av.75 ind m-3) whereas male just observed at station 2 (40 ind m-3) in post-monsoon. Male of P. parvus observed only in station 2 in post monsoon (27 ind m-3), where female showed highest at station 4. Lopes et al. (1999) stated Paracalanus is one of the most important genus in the neurotic region in the Brazilian waters.
Female and male paracalanidae abundance and environmental parameters: According to Table 1 some environmental parameters influence on Paracalanidae sex, similar to some copepod species (Svensen and Tande, 1999). Female A. longicornis, A. gibber, A. gracilis, A. monachus showed negative correlation with pH and positive with temperature. This result suggests female of these species prefer lower pH and higher temperature.
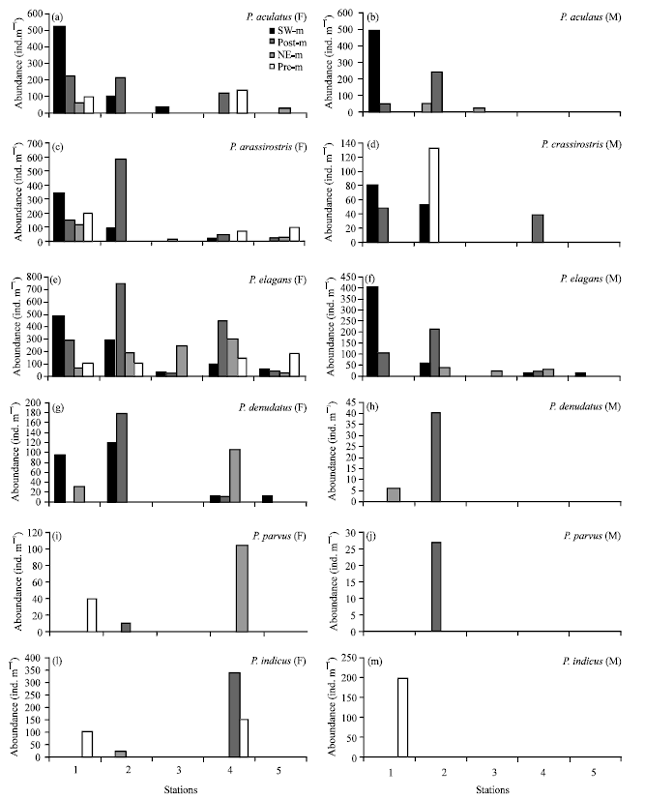 | |
| Fig. 6(a-m): | Abundance of female (F) and male (M) Paracalanus in Chabahar Bay in stations, SW.m: SW-monsoon, post-m: Post-monsoon, NE-m: NE-monsoon and pre-m: Pre-monsoon |
Similar cases have been reported in some copepod species in inland Sea of Japan by Uye and Sano (1995) and in Arabian Sea by Smith (1995) who found positive relationship between female abundance and temperature. Salinity, chlorophyll-a and DO appears to play a minor role in influencing sex of Acrocalanus.
| Table 1: | Pearson correlation of major environmental parameters and female and male paracalanidae abundance |
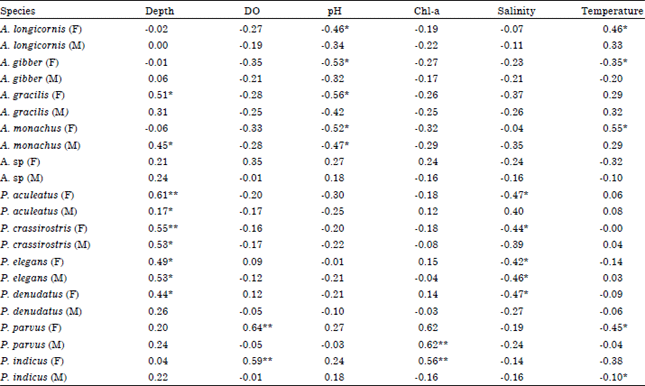 | |
| *Significant at 0.05 level, **Significant at 0.01 level, F: Female, M: Male, DO: Dissolved oxygen | |
No significant relationship was found between those parameters and female and male Acrocalanus abundance. A negative correlation was found between male A. monachus and pH also a positive correlation between depth and male A. monachus and female A. gracilis (Chen, 1986) observed a relationship between vertical range of males and females of copepoda abundance and depth in the eastern tropical Pacific.
Depth and salinity play key role in female and male abundance of P. aculeatus, P. crassirostris, P. denudatus and P. elegans in Chabahar Bay. A positive correlation was found between abundance of male and female P. aculeatus and depth. Also a negative relationship was between salinity and female. The similar trend was observed in P. crassirostris. P. crassirostris has been showed highest abundance in the dry season in Brazil (Araujo, 2006).
Our results showed chlorophyll-a is not key factor controlling abundance of these species while P. aculeatus has been reported that increase in high food concentration (Paffenhofer, 1998).
There was a positive correlation between female and male abundance of P. elegans and depth. A negative correlation was found between abundance of male P. elegans and salinity. Female of P. denudatus showed positive relationship with depth and negative with salinity. This result showed in these species both sex prefer off-shore stations. (Chen, 1986) observed a relationship between vertical range of males and females of copepoda abundance and depth in the eastern tropical Pacific. Overall, female of P. aculeatus, P. crassirostris, P. denudatus and male of P. elegans prefer low saline water. They did not show significant correlation with chlorophyll-a. (Suzuki et al., 1999) stated adult P. indicus feed actively on microprotozoans, especially heterotrophic dinoflagellates and ciliates like Acrocalanus adults and late copepodites that may also feed on microzooplankton (McKinnon, 1996). Therefore, our result could be explained by change in the selectivity of food particles with age in these species.
Depth, salinity and pH appear to play a minor role in influencing sex abundance of P. parvus and P. indicus. No significant relationship was found between those parameters and female and male P. parvus and P. indicus abundance. Female abundance of P. parvus showed positive correlation with chlorophyll-a and DO and negative with temperature. This result showed female abundance of P. parvus increase in high chlorophyll-a concentration. Also, it prefers lower water temperature and higher DO. P. indicus showed positive relationship with chlorophyll-a concentration and DO. In those species male did not show any significant correlation with environmental parameters.
Some studies in P. parvus showed a negative correlation between abundance and depth. They observed this species prefer coastal waters in China (Chen and Zhang, 1974; Peterson et al., 2002; Hwang and Wong, 2005) and colder temperature in Taiwan Strait (Peterson et al., 2002).
Conclusions drawn both from field and experimental studies indicate that the environment may influence abundance of sexes in copepods (Takeda, 1950; Katona, 1970). These researchers have suggested factors like nutrition, temperature, pressure, population density and parasitism as possible inducers. For crustacea in general, it is also supposed that factors like day length (Penelope, 1994), pH, carbon dioxide, UV light, metabolic products and exposure to the opposite sex (Korpelainen, 1990; Kawasaki, 1995) can influence the sex abundance.
In our result, female Paracalanidae always outnumbered than males. Our study suggested 2 hypotheses for that: According to some experiment in copepod, males lived for shorter periods as adults than females even in the presence of excess food (Rhodes, 2004). Also sex change can be found in some Paracalanidae from copepodite stage 5 male to female (Gusmao and McKinnon, 2009).
The present result was similar to other researches in tropical water (Ummerkutty, 1965; Chen, 1986; Uye and Sano, 1995). The sexes abundance in field populations is strongly skewed, with adult females outnumbering males (Hirst and Kiorboe, 2002; Kiorboe, 2006) with males often having shorter durations, is well documented (Uye et al., 1983).
ACKNOWLEDGMENTS
Authors would like to express their sincere gratitude to Dr. Irina Prusova for her useful comments and Saeed Sanjani for assistance in sampling.
REFERENCES
- Ahmed, M.S., M. Raknuzzamman, H. Akther and S. Ahmed, 2007. The role of cyanobacteria blooms in cholera epidemic in Bangladesh. J. Applied Sci., 7: 1785-1789.
CrossRefDirect Link - Araujo, H.M.P., 2006. Distribution of Paracalanidae species (Copepoda, Crustacea) in the continental shelf off sergipe and Alagoas States, Northeast Brazil. Brz. J. Oceanogr., 54: 173-181.
Direct Link - Chen, Y.Q., 1986. The vertical distribution of some pelagic copepods in the eastern tropical pacific. Calcofi Rep., 27: 205-227.
Direct Link - El-Sherbiny, M.M., M.H. Hanafy and M.A. Aamer, 2007. Monthly variations in abundance and species composition of the epipelagic zooplankton off Sharm El-Sheikh, Northern Red Sea. Res. J. Environ. Sci., 1: 200-210.
CrossRefDirect Link - Hirst, A.G. and T. Kiorboe, 2002. Mortality of marine planktonic copepods: Global rates and patterns. Mar. Ecol. Prog. Ser., 230: 195-209.
Direct Link - Hirst, A.G., D. Bonnet, D.V.P. Conway and T. Kiorboe, 2010. Does predation control adult sex ratios and longevities in marine pelagic copepods?. J. Limnol. Oceanogr., 55: 2193-2206.
CrossRefDirect Link - Hsieh, C., C.S. Chen and T.S. Chiu, 2005. Composition and abundance of copepods and ichthyoplankton in Taiwan Strait (Western North Pacific) are influenced by seasonal monsoons. J. Mar. Freshwater Res., 56: 153-161.
CrossRefDirect Link - Hwang, J.S. and C.K. Wong, 2005. The China Coastal Current as a driving force for transporting Calanus sinicus (Copepoda: Calanoida) from its population centers to waters off Taiwan and Hong Kong during the winter northeast monsoon period. J. Plankton Res., 27: 205-210.
CrossRefDirect Link - Irigoien, X., B. Obermuller, R.N. Head, R.P. Harris and C. Rey et al.,., 2000. The effect of food on the determination of sex ratio in Calanus spp.: Evidence from experimental studies and field data. ICES J. Mar. Sci., 57: 1752-1763.
CrossRefDirect Link - Jahangir, T.M., M.Y. Khuhawar, S.M. Leghari and A. Laghari, 2001. Pysico-chemical and biological study of mangho pir euthermal springs Karachi, Sindh Pakistan. J. Biol. Sci., 1: 636-639.
CrossRefDirect Link - Javed, M., 1999. Studies on metal eco-toxicity of river Ravi stretch from shahdera to head baloki. Pak. J. Biol. Sci., 2: 1062-1068.
CrossRefDirect Link - Kiorboe, T., 2006. Sex, sex-ratios and the dynamics of pelagic copepod populations. Oecologia, 148: 40-50.
CrossRefDirect Link - Korpelainen, H., 1990. Sex ratios and conditions required for environmental sex determination in animals. Biol. Rev., 65: 147-184.
CrossRefDirect Link - Kumar, C.S. and P. Perumal, 2011. Hydrobiological investigations in ayyampattinam coast (Southeast coast of India) with special reference to zooplankton. Asian J. Biol. Sci., 4: 25-34.
CrossRefDirect Link - Leghari, S.M., T.M. Jahangir, M.Y. Khuhawar and A. Leghari, 2001. Physico-chemical and biological studies of euthermal sulphur and chliarothermal springs Lakki Shah Saddar (District Dadu) Sindh, Pakistan. J. Biological Sci., 1: 929-934.
CrossRefDirect Link - Gusmao, L.F.M. and A.D. McKinnon, 2009. Acrocalanus gracilis (Copepoda: Calanoida) development and production in the Timor Sea. J. Plankton Res., 31: 1089-1100.
CrossRefDirect Link - McKinnon, A.D. and T. Ayukai, 1996. Copepod egg production and food resources in Exmouth Gulf, Western Australia. J. Mar. Freshwater Res., 47: 595-603.
CrossRefDirect Link - McKinnon, A.D., 1996. Growth and development in the subtropical copepod Acrocalanus gibber. Limnol.Oceanogr., 41: 1438-1447.
Direct Link - Oueda, A., W. Guenda, A.T. Kabre, F. Zongo and G.B. Kabre, 2007. Diversity, abundance and seasonal dynamic of zooplankton community in a South-Saharan reservoir (Burkina Faso). J. Boil. Sci., 7: 1-9.
CrossRefDirect Link - Paffenhofer, G.A., 1998. On the relation of structure, perception and activity in marine planktonic copepods. J. Mar. Syst., 15: 457-473.
CrossRefDirect Link - Peterson, W., 1998. Life cycle strategies of copepods in coastal upwelling zones. J. Mar. Syst., 15: 313-326.
CrossRefDirect Link - Peterson, W.T., J.E. Keister and L. Feinberg, 2002. The effects of the 1997-99 El Nino/La Nina events on hydrography and zooplankton off the central Oregon coast. Prog. Oceanogr., 54: 381-398.
CrossRefDirect Link - Rahimibashar, M.R., A. Esmaeili-Sary, S.A. Nezami, A. Javanshir, S.M.R. Fatemi and S. Jamili, 2009. The planktonic community structure and fluxes nutrients in the sefid-rood river estuary (South Caspian Sea). Res. J. Environ. Sci., 3: 149-162.
CrossRefDirect Link - Smith, S.L., 1995. The Arabian Sea: Mesozooplankton response to seasonal climate in a tropical Ocean. ICES J. Mar. Sci., 52: 427-438.
CrossRefDirect Link - Suzuki, K., Y. Nakamura and J. Hiromi, 1999. Feeding by the small calanoid copepod Paracalanus sp. on heterotrophic dinoflagellates and ciliates. J. Aqua. Microb. Ecol., 17: 99-103.
CrossRefDirect Link - Svensen, C. and K. Tande, 1999. Sex change and female dimorphism in Calanus finmarchicus. Mar. Ecol. Prog. Ser., 176: 93-102.
CrossRefDirect Link - Takeda, N., 1950. Experimental studie so n the effect of external agencies on the sexuality of a marine copepod. Physiol. Zool., 23: 288-301.
Direct Link - Tarkan, A.N., M.I. Inibilir and A.S. Tarkan, 2005. Seasonal variations of the zooplankton composition and abundance in the istanbul strait. Pak. J. Biol. Sci., 8: 1327-1336.
CrossRefDirect Link - Tseng, L.C., R. Kumar, H.U. Dahms, Q.C. Chen and J.S. Hwang, 2008. Monsoon-driven succession of copepod assemblages in coastal waters of the Northeastern Taiwan strait. Zool. Stud., 47: 46-60.
Direct Link - Uye, S.I. and K. Sano, 1995. Seasonal reproductive biology of the small cyclopoid copepod Oithona davisae in a temperate eutrophic inlet. Mar. Ecol. Prog. Ser., 118: 121-128.
Direct Link - Uye, S., Y. Iwai and S. Kasahara, 1983. Growth and production of the inshore marine copepod Pseudodiaptomus marinus in the central part of the Inland Sea of Japan. Mar. Biol., 73: 91-98.
CrossRefDirect Link - Penelope, J.W., 1994. Parental control of sex ratio in Gammarus duebeni, an organism with environmental sex determination. J. Evol. Biol., 7: 177-187.
CrossRefDirect Link