
Kandasamy Kathiresan
Centre of Advanced Study in Marine Biology, Annamalai University, Parangipettai: 608502, Tamil Nadu, India
LiveDNA: 91.8103
Nabeel M. Alikunhi
Centre of Advanced Study in Marine Biology, Annamalai University, Parangipettai: 608502, Tamil Nadu, India







ABSTRACT
Mangroves, seagrasses and coral reefs are the major tropical coastal ecosystems of the world. The physico-chemical and biological interactions between the coastal ecosystems have been perceived, but are rarely explored for better understanding of their connectivity. Mangroves forests and seagrass meadows are the coastal ecosystems of intense nutrient cycling, whereas the coral reef areas are poor in nutrients and Hence, the nutrient-rich coastal systems supply nutrients to adjacent coral reefs. The mangroves and seagrass provide nursery grounds for many faunal species, while the coral reefs provide habitat for them. The mangroves protect seagrass beds and coral reefs from landward discharges and sedimentations, while the coral reefs also protect the other systems by buffering ocean currents and dissipating wave action. The successful connectivity between the three ecosystems is vital for fishery resources, nutrient balance and mitigation of climate change. Hence, the interactions between the ecosystems are not only the state of the biodiversity enrichment but also the flow nutrient and energy. The deleterious effect of any one ecosystem will directly or indirectly affect the other ecosystems. It is a matter of necessity to conserve coastal ecosystems simultaneously in order to ensure a better goods and ecosystem services. Propagation and enrichment of mangrove ecosystem is easier than that of seagrasses and coral reefs and hence, development and diversification of mangrove system is recommended for multiple ecosystem management.
PDF Abstract XML References Citation
Received: May 03, 2010;
Accepted: May 18, 2010;
Published: April 29, 2011
How to cite this article
Kandasamy Kathiresan and Nabeel M. Alikunhi, 2011. Tropical Coastal Ecosystems: Rarely Explored for their Interaction!. Ecologia, 1: 1-22.
DOI: 10.3923/ecologia.2011.1.22
URL: https://scialert.net/abstract/?doi=ecologia.2011.1.22
DOI: 10.3923/ecologia.2011.1.22
URL: https://scialert.net/abstract/?doi=ecologia.2011.1.22
INTRODUCTION
Coastal ecosystems are biologically diverse, ecologically productive and economically valuable. The tropical coastal ecosystems mainly encompass mangroves, seagrasses and coral reefs. These coastal systems often occur in close proximity to one another and interact with each other. The physico-chemical and biological interactions between the three major coastal ecosystems have been perceived, but are rarely explored. They often have a synergistic relationship, which leads to coastal environmental stability. Mangroves and seagrasses stabilize the sediments, slow down the water movements and trap the heavy metals and nutrient rich runoff, thus improving the water quality for corals and fish communities. They also enhance the biomass of coral reef fishes by providing nursery habitats. Coral reefs, in turn, stabilize the seascape by buffering ocean currents and dissipating wave action and favour the colonization of mangroves and seagrasses.
Mangrove ecosystem-the epicenter of tropical coastal productivity: Mangrove wetlands are prominent coastal features in tropical environment (Kathiresan and Bingham, 2001; Kathiresan and Qasim, 2005). Numerous studies have shown that mangroves are among the most productive wetland communities in the world (Lugo and Snedaker, 1974; Day et al., 1987; Twilley, 1988; Kathiresan and Bingham, 2001; Kathiresan and Qasim, 2005). Many goods and services have been attributed to mangroves that: (1) provide food and shelter to consumer fishes, (2) improve the water quality of adjacent estuaries, (3) regulate the processes of estuarine chemical cycles, (4) contribute to shoreline protection and (5) produce a variety of forest products (Kathiresan and Bingham, 2001; Kathiresan and Qasim, 2005). Mangrove ecosystem is mainly detritus-based system. It produces large amounts of litter in the form of falling leaves, branches and other debris. Litter produced from the canopy of mangrove forests represents a major source of organic matter and nutrients for outwelling to adjacent coastal waters (Odum and Heald, 1972; Twilley, 1988). Monetary value of the mangroves is estimated at US$ 9990/ha/year which is greater than that of coral reefs, continental shelves and/or the open sea (Costanza et al., l998). The economic value of small scale fisheries alone is US$ 37,500 per hectare of mangrove fringe in the Gulf of California (Oropeza et al., 2008).
Seagrasses beds-the fuel for coastal foodweb: Seagrasses are the only submerged flowering plants that grow in extensive beds in many coastal and estuarine areas of the world (Fourqurean and Robbins, 1999). They often dominate temperate shallow environments, forming productive habitats of complex structure that are rich in biodiversity (Clough et al., 1998). They serve in important ecological functions by providing habitat for a wide variety of plant and animal species and by physically stabilizing dynamic coastline, through the reduction of coastal erosion (Orth, 1977). They are also one of the important contributors to the organic matter in coastal environments (Canuel et al., 1997). In addition, seagrasses support subsistence and commercial fisheries worth as much as $3500/ha/year (Waycott et al., 2009). The productive assemblage of epiphytic algae on seagrasses sustain invertebrate grazers (Moncreiff and Sullivan, 2001), while seagrass detritus support consumer fish production through bacterial and fungal intermediates or remain unutilised in a trophic dead end as particulate organic carbon in marine sediments (Jones et al., 2003). The net organic matter production may accumulate in local sediments or can be transported to adjacent coastal waters through the export of the detrital material depending on the hydrodynamic conditions (Madsen et al., 2001).
Coral reefs-the biological paradise: Coral reefs are among the most biologically diverse and productive ecosystems in the world. They occur in tropical areas, where oceanic waters are generally very low in nutrients, yet these complex ecosystems have some of the highest biomasses and productivity (Lewis, 1977). It is generally believed that the main evolutionary adaptation to low nutrient conditions in reef environments has been the evolution of inter-relationships that lead to efficient recycling of nutrients. Due to the three-dimensional complexity of the coral reefs, niche diversification is well pronounced and it confirms the principle of diversity begetting diversity. This is particularly evident in the abundance and importance of symbiotic interactions; i.e., compared to temperate environments, the seasonally less variable nature of the tropics reflect a high degree of specialization and abundance of often complex and specific symbioses in order to maintain productivity (Paulay, 1997). Because of this fact, the coral reefs support large fishery resources (Lewis, 1977; Nagelkerken et al., 2000a, b, 2001). The reef fishes exhibit diel migration patterns, whereby they graze on the back reef and in the lagoons by day, but shelter in the deeper reef by night. Materials that the fishes transport in their guts at dusk are defecated over the fore reef or in their nocturnal shelters (Nagelkerken et al., 2000a, b). This form of transport may be an important means of upstream nutrient recycling on coral reefs.
Connectivity between coastal ecosystems: During the last three decades, ecology has evolved toward a vision of the ecosystem as a system of interactions where the center of interest is no longer the state of the biomasses of the different groups of organisms, but the interactions between them, quantified by flows of matter or energy (Fasham, 1984). The ecosystems of mangroves, seagrasses and coral reefs are functionally linked, but it is not properly understood for better management practices. The macro-level linkages between coastal ecosystems are at least known but there may be other linkages that are not known yet, especially at microlevel. The connectivity between coastal ecosystems is physico-chemical and biological linkages among habitats and ecosystems. The interaction between coastal ecosystems can influence both nutrient accumulation and loss (Ogden and Zieman, 1977) which makes the system more balanced. Nutrient conservation mechanisms are a function of the living system Hence, any disturbance can result in rapid loss of nutrients and the system must recover through successional processes (Ogden and Zieman, 1977; Atkinson et al., 1981).
In addition to physical and chemical connectivity, the biological connectivity which is mainly through the dispersal of organisms by active migration and ocean currents also exist. Mangroves and seagrasses also enhance the biomass of coral reef fish species by providing nursery habitats (Mumby et al., 2004; Unsworth et al., 2008). The strength of connectivity depends on the abundance and fecundity of diverse living organisms, the longevity and pre-competency periods of their larvae and the spawning sites and movement patterns of adults. The food resources provided through these diverse living organisms in the coastal marine ecosystems are essential for maintaining human adaptive capacity to waning resource availability. The biodiversity, in turn speaks about the healthiness of the ecosystem (Kay and Schneider, 1994). Hence, connectivity is imperative for the biodiversity replenishment in coastal habitats damaged by natural or human-related agents, including climate change. It is imperative to secure pathways of connectivity between mangroves, seagrass beds and coral reefs to enhance resilience (Mumby et al., 2004). However, the connectivity between the coastal ecosystems is not clearly studied in many part of the world. More studies are required for functional understanding of the coastal systems. The macro-level linkages between coastal ecosystems are at least known but there may be other linkages that are not known yet. This article reviews about the different kinds of the interrelation and connectivity between coastal marine ecosystem and various tools for studying it.
THE FOREMOST ENDEAVOR!
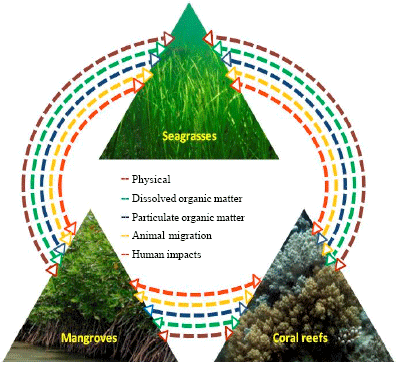
Realizing the importance of the interactions among coastal ecosystems, UNESCO was the first to conduct a workshop in 1982, on Coral reefs, seagrass beds and mangroves: Their interaction in the coastal zones of the Caribbean in US Virgin Islands, West Indies (UNESCO, 1983). The health and productivity of individual habitats existing in coastal zone is dependent on a number of complex interactions, which are classified into five major types: namely (1) physical, (2) dissolved organic matter, (3) particulate organic matter, (4) animal migration and (5) human impacts (Fig. 1; UNESCO, l983).
Physical interaction: Reduction of water energy, flow regulation and sediment relationships are important physical interactions among the ecosystems. Coral reefs act as the hydrodynamic barriers, which dissipate wave energy and favour a relatively calm environment for colonization of mangrove and seagrasses communities.
 | |
| Fig. 1: | Interactions between coastal ecosystems (Modified from UNESCO, 1983) |
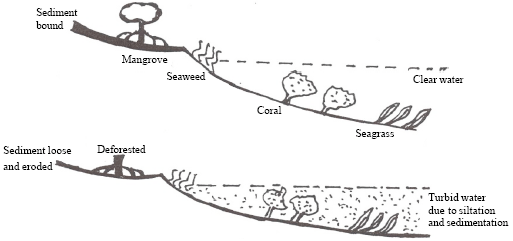 | |
| Fig. 2: | Trapping of sediments by mangroves to better the water quality for colonization of seaweeds, seagrasses and corals (Adopted from Kathiresan and Bingham, 2001) |
Due to the wave action and water currents, the sediments in the reef lagoons form shoals and islands which in due course may be colonized by seagrasses and mangroves (Atkinson et al., 1981). On the other hand, mangroves and seagrass act as biofilter and facilitate deposition of sediments from river run-off and make the water clear for better photosynthesis and growth of zooxanthellae associated with coral reefs (Fig. 2; (Kathiresan and Bingham, 2001; Kathiresan and Qasim, 2005). Mangroves also regulate fresh water flow into the coastal areas (Kathiresan and Bingham, 2001). The excess fresh water which flows into this ecosystem and also dilutes the excess salts accumulated in the mangrove environment during drought and thus probably buffering the salinity changes.
Dissolved organic matter: Dissolved Organic Matter (DOM) is ubiquitous and abundant form of nutrient. Several studies consistently indicate that a significant fraction of primary production of mangroves can be exported through tidal currents as dissolved organic matter (Twilley, 1985; Boto and Wellington, 1988; Ayukai et al., 1998; Dittmar et al., 2001; Dittmar and Lara, 2001a; Nabeel, 2010). The mangroves contribute considerable quantities of organic matter, primarily as leaves, to adjacent waters and sediments as well coral reef environment (Heald, 1971; Lugo and Snedaker, 1974). In addition to mangroves, seagrasses also provide DOM to the oligotrophic coral reefs (Robertson and Blabber, 1992). Tidally mediated flux of sediment pore water to the ocean seems to be the principal pathway of DOM outwelling (Dittmar and Lara, 2001b). Surprisingly, despite enormous ecological differences among the mangroves in various parts of the world, the area-normalized DOM export from mangroves appears to be independent of location (Twilley, 1985; Boto and Wellington, 1988; Dittmar et al., 2001).
Particulate organic matter: Particulate Organic Matter (POM) is a vital source of organic matter to the intertidal zone (Hemminga et al., 1994; Bouillon et al., 2003, 2007). It plays an important role in biogeochemical processes and cycles of carbon and other bio-reactive elements in open oceans and marginal seas (Twilley, 1985; Boto and Wellington, 1988; Walsh, 1991). The mangrove forests and seagrass beds contribute to detritus and POM in water column and sediments of coastal biotope (Kathiresan and Bingham, 2001; Hemminga et al., 1994; Lee, 1995; Bouillon et al., 2004). Mangrove-derived material is exported as POM to the aquatic environment for sustaining the pelagic and benthic food webs (Robertson and Blabber, 1992; Clough et al., 1998; Kathiresan and Bingham, 2001). Likewise, input of POM from seagrass detritus to the open water has been proved (Maie et al., 2008). Hence, as far as POM fluxes are concerned, the mangrove forest and adjacent seagrass meadows are tightly coupled, but the nearby coral reefs may exists in relative isolation (Hemminga et al., 1994).
Animal migration: Animal migration provides an important link among coral reefs, seagrass beds and mangroves. Merriam (1984) was the first to stress the importance of habitat connectivity and movements of biota among the habitats. Many species are dependent on the maintenance of connectivity between habitats to complete their life cycles (Zeller, 1998). Many aquatic organisms undertake ontogenetic migrations from one habitat to another during their lifetime. Inter-habitat migrations do occur with invertebrate (Hiddink, 2003) and vertebrate fauna (Law and Dickman, 1998; Nagelkerken et al., 2000a, b). Mangroves and seagrass beds are vital nursery areas for many important coral reef fishes (Mumby et al., 2004; Unsworth et al., 2008). The nursery functions of the mangroves and seagrasses are mainly due to the availability of shelter for juvenile organisms and abundant supply of food in the form of organic detritus (Robertson, 1986; Robertson and Daniel, 1989; Camilleri, 1992; Kathiresan and Bingham, 2001; Rajendran and Kathiresan, 2004, 2007).
Human impacts: The coastal ecosystems such as mangroves, seagrasses and coral reefs are critical habitats which are biologically productive, but ecologically sensitive to human impacts. The human population has both positive and negative impacts on the connectivity of coastal ecosystems. The anthropogenic nutrient inputs into shallow systems may yield different results with regard to the productivity. Increased nutrient levels in the water column of oligotrophic waters stimulate the growth of phytoplankton, macroalgae (including seaweeds) and epiphytic algae, as well as the seagrasses (Rabalais and Nixon, 2002; Smith, 2003). This may enhance the productivity of the oligotrophic coral reef environment. However, the eutrophication can often lead to seagrasses being outcompeted by a heavy overgrowth of macroalgae (Short et al., 1995). In Western Australia, heavy epiphyte fouling is due to nutrient-rich effluents, leading to extensive seagrass losses (Cambridge et al., 1986). The artificial dredging facilitates the free runoff of water from mangroves to coral reefs through seagrass beds. However, the runoff of water along with sediments can affect water quality of the coastal ecosystems leading to light reductions from increased turbidity which cause decline in seagrass beds (Hall et al., 1999).
CARBON OUTWELLING FROM COASTAL ECOSYSTEMS
A central paradigm of coastal ecology over the past four decades describes the outwelling of organic matter from coastal wetlands, as a major source of energy that supports much of the secondary production of estuaries and nearshore waters. Odum (1968) proposed this break-through hypothesis in coastal ecology describing primary ecological function of mangroves in the coastal zone and described it as the outwelling hypothesis. The mangrove leaf litter provides trophic subsidy in adjacent coastal waters, through a detritus-based food web whereby mangrove litter is converted into more palatable microbial biomass, which in turn, acts as the dominant food source for higher trophic levels (Odum and Heald, 1975). Although, there is little direct evidence that wetland detritus supports the nutritional requirements of higher trophic assemblages in adjacent coastal environments, the concept is generally acceptable by the scientific community (Nixon, 1980). The outwelling idea was first questioned by Haines (1975) who reported that Georgia and South Carolina shelf ecosystems received minor inputs from outwelling and that the most important process in maintaining high rates of secondary productivity was in situ primary production. Followed by this, several studies were conducted on outwelling hypothesis. However, the role of mangroves as carbon source in adjacent ecosystems is still under debate.
Mangroves as carbon source in coastal ecosystems: Among the primary producers mangrove play an important role in the carbon balance of tropical coastal ecosystems (Kathiresan and Bingham, 2001; Kathiresan and Qasim, 2005; Kathiresan, 2008). However, the role of mangrove litter in sustaining nearshore or adjacent aquatic secondary production continues to be an issue of debate. About 50% of net primary production in mangroves is exported as organic matter to the ocean (Robertson and Blabber, 1992; Dittmar and Lara, 2001a, b; Jennerjahn and Ittekkot, 2002) which is almost 2 orders of magnitude higher than the global average for terrestrial ecosystems (Spitzy and Leenheer, 1991). The mangroves contribute around 15% to the organic carbon globally accumulating in marine sediments (Twilley et al., 1992; Jennerjahn and Ittekkot, 2002). Similarly, Dittmar et al. (2006) estimated that mangroves contribute 10% of the terrestrial Dissolved Organic Carbon (DOC) exported to the ocean globally, despite their small area relative to other habitats. Mangroves are efficient habitats for carbon sequestration: about 2.4-fold as high as saltmarshes, 5.2-fold as high as seagrasses and as much as 50 times the amount of carbon in their sediment per hectare of tropical forest (Kathiresan, 2008). This potentially large role of mangrove carbon in the coastal zone highlights the importance of understanding mangrove ecosystem in carbon dynamics to better construct the global oceanic carbon budgets.
Fueling of coastal food web by mangrove carbon: Mangroves provide an important nutrient base for food webs leading to enhancement of fish resources in coastal areas (Turner, 1977; Fell and Master, 1981; Rajendran and Kathiresan, 1998, 1999a, b, 2000, 2004, 2007; Kathiresan and Bingham, 2001). Considerable quantities of organic matter, primarily leaves derived through primary production, are exported to adjacent waters and the large flux of mangrove detritus to the coastal ocean can have recognizable effects on aquatic food webs (Odum and Heald, 1975; Alongi et al., 1989; Alongi et al., 2000; Kathiresan and Bingham, 2001; Kathiresan and Qasim, 2005; Jennerjahn and Ittekkot, 2002; Mumby et al., 2004; Moriniere et al., 2004).
The connectivity that exists between mangroves and fishes is in the form of trophic (flow of material from mangroves to adjoining habitats where different stages of life cycle), or physical contacts between the habitats (migration pathways for completion of different life history stages) is still debated (Lee, 2004). A positive correlation between nearshore catch of fishery organisms (shrimp or fish) and mangrove area has been reported for many tropical regions (Macnae, 1974; Turner, 1977; Jothy, 1984; Sasekumar et al., 1992; Primavera, 1996; Kathiresan and Rajendran, 2002; Oropeza et al., 2008). Correlations do not mean causal relationships; however, they show that where extensive mangrove habitats are present, productive fisheries exist. Turner (1989) predicted a reduction in natural shrimp production by 15x106 kg year-1 after the removal of 250 km2 of mangrove forest in Ecuador. It was reported from the Gulf of California that fisheries landings are positively related to local abundance of mangroves and mangrove-related fish and crab species account for 32% of the small scale fisheries landings in the region (Oropeza et al., 2008).
The role of mangroves as the primary food source of prawns has been explained with the help of stable isotopes of C and N, in the estuaries of the Matang mangrove swamps (Chong et al., 2001). However, contribution of mangroves to food web of fishes is often under debate throughout the world. For example, Stoner and Zimmerman (1988) while studying the mangrove fringed lagoon in Puerto Rico, observed that penaeid prawns and their food items derived their organic carbon mostly from benthic algae rather than from mangrove detritus. The penaeid prawns in a mangrove-fringed river in the Philippines are more dependent on phytoplankton and epiphytic algae than on mangrove leaves, as suggested by Primavera (1996). While studying stable C and N isotopes in the tissues of prawn species in relation to several sources of primary producers in tropical Northeastern Australia, Loneragan et al. (1997) emphasized that the seagrass beds and their epiphytes are a major source of energy that supports the food web of prawns in estuary. A study conducted in mangrove plantation of the south India revealed that microbes associated with decomposing mangrove leaves contribute more carbon and nitrogen to the food web inside the mangrove forest, but this role is transferred to phytoplankton in off-shore region (Nabeel et al., 2010a). Thus, mangroves act as an indirect source through microbes in coastal food web. However, many tropical coastal food webs lack a clear signal of mangrove detritus in the tissue of estuarine-dependent organisms. The majority of studies have found that mangrove organic matter is exported and incorporated into coastal food webs only to a very limited extent (Lee, 1995).
Recently, a study was conducted by Nabeel (2010) in the Islands of Gulf of Mannar, south east India, where the mangroves, seagrass and coral reefs are distributed in close proximity. The results of various analysis using stable isotopes, fatty acid and other biochemical markers revealed the mangroves produce litter facilitating microbial decomposition. The nutrients released in this process enhance the microbial and planktonic populations. The organic matter associated with microbes is transferred to adjacent coral reef ecosystem through seagrass beds (Fig. 3). The mangroves contribute significantly to foodweb through the indirect pathway and thus the coastal ecosystems are interconnected (Nabeel, 2010).
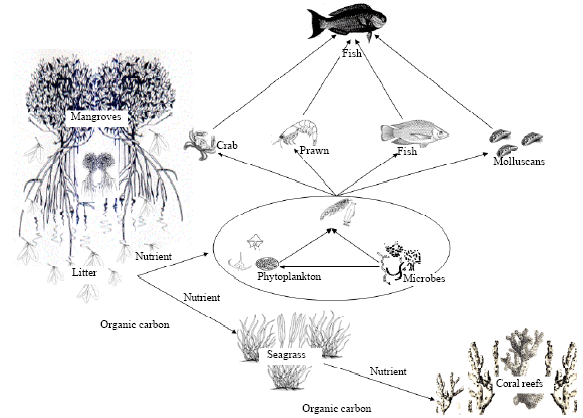 | |
| Fig. 3: | Diagrammatic representation of the trophic and functional linkage among coastal ecosystems (Adopted from Nabeel, 2010) |
METHODS OF STUDYING THE INTERACTIONS BETWEEN COASTAL ECOSYSTEMS
Stable isotopes as a tracer: Stable isotope measurements have been used extensively since l970s as indicators of organic matter sources to investigate the trophic ecology of aquatic ecosystems. The isotopic approaches have improved our knowledge of organic matter flow, food web structures, trophic transfers, resource partitioning, ontogenetic shifts in consumer diets, habitat usage and species migration in various parts of the world (Kneib and Stiven, 1980; Zieman et al., 1984; Muscatine et al., 1989; Michener and Schell, 1994; Newell et al., 1995; Kwak and Zedler, 1997; Deegan and Garritt, 1997; Moncreiff and Sullivan, 2001 Kharlamenko et al., 2001; Ramos et al., 2003; Lepoint et al., 2004; Persic et al., 2004; Abed-Navandi and Dworschak, 2005; Nyunja et al., 2009; Nabeel et al., 2010a). Recently, multi sources mixing models based on stable isotopes have been proposed to derive the relative contributions of each source when more than one primary source is considered (Phillips and Gregg, 2003). The multiple-isotope approaches are used to identify the contribution of different sources of organic matter (Peterson et al., 1985). However, naturally occurring stable isotopes of elements such as carbon (δ13C) and nitrogen (δ15N) are more effectively used in coastal ecosystems (Michener and Schell, 1994). The δ13C and δ15N data of aquatic organisms can, therefore, provide useful information about food sources and trophic level. The nitrogen isotope can be used to identify the relative trophic position of various organisms within the food web, while carbon isotope provides information regarding the source of primary production in the ecosystem and the flow of carbon from primary producers to consumers (Gu et al., 1996).
Fatty acids as biomarker: Fatty acids are carbon-rich compounds that are ubiquitous in all organisms and that are relatively easy to metabolize when consumed as part of the animal’s diet. The fatty acids are biologically important because they may contribute energy resources, essential nutrients for survival and growth and may be integral components of cell membrane structures and function of an organism (Hazel et al., 1991; Parrish, 1998; Parrish et al., 2000). The concept of Fatty Acids (FA) being transferred conservatively through aquatic food webs was first suggested in 1935 (Lovern, 1935). This seminal work found that Calanus finmarchicus could be distinguished from three freshwater copepod species based on the concentrations of various FA. Almost 30 years later, Kayama et al. (1963) performed one of the first experiments demonstrating the transfer of FA through a linear, experimental food web consisting of Chaetoceros simplex (diatom)-Artemia salina (branchiopod)-Lebistes reticulatus (freshwater guppy). This trophic biomarker approach makes use of the fact that specific lipid components, particularly fatty acids, are characteristic of specific groups of organisms. These marker fatty acids are incorporated largely unaltered into the consumers, which thus retain a signature of their dietary origin (Sargent and Whittle, 1981).
Several studies have successfully used fatty acids to trace the transfer of organic matter in coastal and estuarine food webs (Kharlamenko et al., 1995; Napolitano et al., 1997; Meziane and Tsuchiya, 2000; Bachock et al., 2003; Hall et al., 2006; Nabeel et al., 2010b). Due to their biological specificity and the fact that they are transferred from primary producers to higher trophic levels without change, make fatty acids suitable for use as biomarkers (Parrish et al., 2000). For example, fatty acids with greater than 24 carbon atoms are synthesized mostly by vascular plants (Volkman et al., 1980) and are considered indicative of higher plant markers in sediments (Scribe et al., 1991). The odd carbon-numbered and branched-chain (ISO-and ANTEISO-) fatty acids and the monounsaturated fatty acids (MUFA) like 18:1u7 are generally considered to be synthesized by bacterial communities (Jeffries, 1972; Volkman et al., 1980) and are therefore used as biomarkers of bacteria (Parkes, 1987). Polyunsaturated Fatty Acids (PUFAs) such as 20:5u3 and 16:4u1 are used as diatom markers in marine habitats (Meziane and Tsuchiya, 2000).
The advancement of various chemical tracer techniques, such as stable isotopes and fatty acid, has significantly contributed to the analysis of organic matter flow (Canuel et al., 1995; Lee, 1995; Meziane and Tsuchiya, 2000). While both of these approaches have limitations, the combination of these techniques has proved to be a powerful tool to determine trophic interactions within complex food webs (Kharlamenko et al., 2001; Ramos et al., 2003). For example, some fatty acid markers may be metabolized and transformed once consumed by the animal and only relative rather than absolute amounts may be measured. Thus, biomarkers should be used with caution or in conjunction with other quantitative techniques, like stable isotopes.
n-alkenes as biomarker: Long-chain n-alkanes with the number of carbon ranging from 25 to 35 are characteristic components of epicuticular waxes of mangrove leaf surfaces and Hence, they can be used as biomarker of the higher plant remains (Dodd et al., 1995, 1998; Rafii et al., 1996; Versteegh et al., 2004; Tanner et al., 2010). The distribution pattern of long-chain n-alkanes (C21-C35) in marine sediments can be used as a source indicator to differentiate between terrestrial organic input and marine-derived organic matter (Prahl et al., 1994; Pearson and Eglinton, 2000). Terrestrial plants have a greater abundance of long-chain n-alkanes (usually C25-C35) with predominance of a strong odd-to-even carbon number (Kolattukudy et al., 1976; Eglinton and Hamilton, 1967; Cranwell, 1973). In contrast, marine organisms usually produce abundant short-chain n-alkanes (C14-C22) while long-chain n-alkanes are minor components (Sakata et al., 1997). Short-chain n-alkanes, particularly the C15 and C17 are presumed to originate from plankton (Cranwell, 1982). The concentration ratio of (C27 + C29 + C31)/(C15 + C17 + C19) n-alkanes is used to assess terrestrial/marine derived organic matter in sediments (Fernandes and Sicre, 2000; Silliman and Schelske, 2003). This approach has successfully been applied elsewhere (Dittmar et al., 2001; Zimmerman and Canuel, 2001; Jaffe et al., 2001).
The most abundant lipid at the Rhizophora leaf surface is C29 n-alkane, which accounts for 0.22% of the dry leaf material (Versteegh et al., 2004; Mead et al., 2005). It is suggested that when the n-alkanes derived from two different sources, i.e., when an algal signal is super-imposed on a higher plant distribution, microbial oxidation of the latter prior to incorporation in the sediment could generate the formation of type of n-alkane-2-one (Volkman et al., 1980). However, the n-alkane-2-one has been later reported to be present in biomass of higher plants and phytoplankton (Rieley et al., 1991) suggesting direct biological inputs to sediments. The distribution of n-alkan-2-one in sediments does not frequently resemble that of n-alkanes, suggesting that n-alkan-2-one may have alternative sources (Jaffe et al., 1993). Cranwell et al., (1987) reported the occurrence of n-alkan-2-one in the dominant plants, including freshwater wetland vegetation, seagrasses and mangroves and suggested these as direct sources of n-alkan-2-ones to sediments. Ficken et al. (2000) proposed an n-alkane proxy (Paq) for discriminating between submerged/floating and emergent macrophytes. Paq is defined as the concentration ratio of (C23 + C25) over (C23+ C25 + C29 + C31) n-alkanes (Ficken et al., 2000; Mead et al., 2005). It is used to differentiate seagrass from mangrove organic matter (Mead et al., 2005). This proxy takes advantage of the fact that submerged/floating species have abundant mid chain n-alkanes relative to emergent macrophytes and terrestrial plant vegetation, which are enriched in the longer chain length n-alkanes. This method is particularly useful when dealing with organic matter produced by biomass utilizing different carbon fixing pathways (Jaffe et al., 2001; Keeley and Sandquist, 1992).
Triterpenoids and sterol as biomarkers: Pentacyclic triterpenoids and sterols are common chemical constituents of marine and terrigenous plants (Kulshreshtha et al., 1972) and have frequently been used to characterize sources of sedimentary organic matter (e.g., Volkman, 1986; Yunker et al., 1995; Mudge and Norris, 1997; Munoz et al., 1997). Dodd et al. (1998) reported 11 triterpenoids in epicuticular waxes which account for up to 3.5% of Rhizophora mangle leaves. Higher plant-derived triterpenoids, pentacyclic 3-oxytriterpenoid compounds with the carbon skeleton of taraxerol, amyrin and lupeol are reportedly derived from higher plants (Ten Haven et al., 1992; Killops and Frewin, 1994), but are rarely observed in other organisms.
The sterols are commonly used as biomarkers of various organisms. The C28 sterol (24-norcholesta-5,22E-dien-3b-ol, 24-methylcholesta-5,22E-dien-3b-ol and 24-methylcholesta-5, 24 (28)-dien-3b-ol) are commonly used as biomarkers of diatoms (Volkman, 1986). The C29 sterols are traditionally used as biomarkers of terrestrial Organic Matter (OM) inputs (Huang and Meinschein, 1978, 1979). Three sterols normally associated with vascular plants: 24-ethylcholesta-5,22E-dien-3b-ol (stigmasterol), 24-methylcholest-5-en-3b-ol (campesterol) and 24-ethylcholest-5-en-3b-ol, are used as tracers for terrigenous plant organic matter (Volkman, 1986). However, assignment of these sterols exclusively to terrigenous vascular plant material is compromised by their presence in some, particularly freshwater phytoplankton (Volkman, 1986, Volkman et al., 1988). Thus, the sum of stigmasterol, campesterol and 24-ethylcholest-5-en-3bol is attributed to a combined vascular plant/riverine algal source, which although not strictly terrigenous, is nonetheless unique to riverine organic matter. The sterol composition of crustaceans in general and zooplankton in particular, is usually dominated by cholesterol (cholest-5-en-3b-ol) (Volkman, 1986; Killops and Killops, 1993). However, cholesterol can be present in low levels in microalgae as well as aquatic and terrigenous plants (Volkman, 1986; Canuel and Martens, 1993).
Biomass analysis show that mangrove leaves contain exceptionally high abundance of taraxerol to 1.4 mg g-1 (Versteegh et al., 2004), while seagrasses do not biosynthesize this compound. Hence, taraxerol is chosen as a specific biomarker for mangroves. Taraxerol is present mostly in leaves, whereas β-amyrin (olean-12-en-3β-ol) is dominant in epicuticular waxes. Leaves of Rhizophora have high amounts of germanicol (olean-18-en-3β-ol) and lupeol (lup-20(29)-en-3β-ol). Avicennia germinans contains betulin (lup-20 (29)-en-3β, 28β-diol), lupeol and β-sitosterol (24-ethylcholest-5-en-3β-ol). Oku et al. (2003) suggested that triterpenoids may have a special function in the adaptation of mangrove to salt stress. While betulin tracer of Avicennia can be efficiently degraded in the sediments, taraxerol seems to be unreactive with respect to microbial attack (Killops and Frewin, 1994; Hernes et al., 2001; Versteegh et al., 2004). Consequently, taraxerol can be a useful biomarker for paleoenvironmental reconstructions.
Lignin and tannin as biomarkers: Lignin is a nitrogen-free co-polymer of various phenyl propenyl alcohols that is present in vascular plants. It is formed as a product of enzymatic reactions between cellulose, tyrosine and phenylalanine in the process of plant tissue lignification (Donaldson, 2001; Micic et al., 2002). Lignin is a major biomass component of trees, grasses, mosses and ferns (Hatcher et al., 1995) and it constitutes about 25% of the tree tissue. As such, it is the second most abundant biopolymer in the environment. The lignin derived from different plant species or even plants of the same species may differ due to variation in the proportion of monomers from which they are formed (Li et al., 2002). It has been frequently used to characterize the contribution of land-derived organic matter to marine (Hedges and Mann, 1979; Hedges et al., 1988), coastal (Hedges and van Geen, 1982; Staniszewski et al., 2001), estuarine (Readman et al., 1986), river (Farella et al., 2001; Pepe et al., 1995) and lacustrine sediments (Hu et al., 1999; Hedges et al., 1982; Ishiwatari and Uzaki, 1987) and to quantify major plant classes contributing to the organic matter. Both the presence and proportions of the oxidation products are commonly utilized as indicators of sedimentary organic matter quality (Farella et al., 2001; Fisher et al., 2003; Hedges et al., 1982; Hu et al., 1999; Miltner and Emeis, 2001; Pempkowiak and Pocklington, 1983; Staniszewski et al., 2001). Lignin is generally more refractory than other molecular compounds and is resistant to microbial attack (Marchand et al., 2005). Hence, both the composition and concentration of lignin are used as biomarkers of organic matter (Pempkowiak et al., 2006).
Tannin is the fourth most abundant component of vascular plant. In addition to quantitative importance, its unique structure and reactivity may lead to new understanding of carbon digenesis (Hernes and Hedges, 2004). The tannin in vascular plants occurs as two types-condensed tannin and hydrolysable tannin-which are more abundant in plant leaves than in woody tissues and contributing to the color and astringency of the bulk organic matter. The tannin content of mangrove leaves is higher and more polymerized than that found for 40 other dicotyledonous plant species (Hernes et al., 2001). Condensed tannin consists of 80% procyanidin and 20% prodelphinidin. The latter, with its higher degree of hydroxylation, appears to be more labile than procyanidin tannin. Leaching induces an increase in polymerization of condensed tannin, which is an important mechanism for tannin removal from leaves. Hence, the study of the tannin in natural samples shows great promise for addressing various geochemical questions.
Nucleic acid as biomarker: Analysis of nucleic acids through a variety of molecular techniques provide insights into the specific origin of DNA encountered in the ecosystem (Danovaro et al., 2005). Among the biochemical classes of organic compounds, DNA ranks fourth after carbohydrates, proteins and lipids (Dell’anno et al., 1998). The presence of DNA as a constituent of the dissolved organic matter pool in aquatic systems has been known since the early 1970s (Pillai and Ganguly, 1972). The organic carbon inventory is largely dominated by non-living materials (i.e., detrital carbon) present in both the dissolved and particulate states, whose cycling is primarily mediated by heterotrophic prokaryotes (DeFlaun et al., 1986). Hence, studies on microbial ecology are very essential for better understanding of biogeochemical cycles (Danovaro et al., 2005). The molecular techniques using nucleic acids will help in identification of microorganisms without using culture-dependent methods as majority (>99%) of microbes are unculturable (Amann et al., 1995). In addition to this, DNA microsatellites through individual assignment-based methods and genetic tagging are used in genetic studies of fisheries connectivity (Carmen and Ablan, 2006). Hence, the DNA is useful complements which provide substantial evidences for all the other biomarkers in inter-relation studies.
CONCLUSION
The functional linkage between critical habitats such as mangroves, seagrass and corals are rarely investigated but vital aspect of marine research. The knowledge of the ecological interactions between ecosystems is rudimentary and far from clear. Coastal ecosystems have significant multiple benefits that are global (long term carbon sequestration) to local (community fisheries). Any reduction of connectivity between coastal ecosystems by habitat fragmentation will be detrimental to coastal ecology and fishery economy. Conservation of coastal ecosystems is urgently required in order to evolve protocol for synergic improvement of coastal habitats simultaneously and to evolve management-cum-monitoring protocol for ensuring better inter-ecosystem production. Because of the interdependence of these habitats, the deleterious effects on one particular system are not necessarily confined to that habitat. Hence, the conservation efforts should protect connected corridors of mangroves, seagrass beds and coral reefs and it is very essential to secure pathways of connectivity among them (Mumby et al., 2004). Interactions between interconnected coastal ecosystems need to be studied for better understanding with the help of advanced tools such as stable isotopes, fatty acids and other molecular biomarkers such as DNA, n-alkenes, terpenoids, lignins and tannins.
Restoration and conservation of mangroves are easier than those of seagrass and coral reefs. However, experimental studies are warranted on the impact mangrove restoration on other coastal ecosystems in terms of connectivity, biodiversity, productivity and other ecosystems services and the impact has to be monitored at a long term basis. This would provide more information for manipulation of coastal ecosystems in management practices for augmenting the marine resources better and long term carbon sequestration.
ACKNOWLEDGMENTS
The authors are thankful to Prof. T. Balasubramanian, Director of this Centre and authorities of Annamalai University for providing all facilities.
REFERENCES
- Moncreiff, C.A. and M.J. Sullivan, 2001. Trophic importance of epiphytic algae in subtropical seagrass beds: evidence from multiple stable isotope analyses. Marine Ecol. Prog. Ser., 215: 93-106.
Direct Link - Moriniere, E.C., I. Nagelkerken, H. van der Mejj and G. van der Velde, 2004. What attracts juvenile coral reef fish to mangroves, habitat complexity or shade. Marine Biol., 144: 139-145.
Direct Link - Mudge, S.M and C.E. Norris, 1997. Lipid biomarkers in the Conway Estuary (North Wales, UK.): A comparison between fatty alcohols and sterols. Marine Chem., 57: 61-84.
CrossRef - Mumby, P., A. Edwads, E.A. Gonzalez, K. Lindeman and P. Blacwell et al., 2004. Mangroves enhance the biomass of coral reef fish communities in the Caribbean. Nature, 427: 533-536.
Direct Link - Munoz, D., M. Guiliano, P. Doumenq, F. Jacquot, P. Scherrer and G. Mille, 1997. Long term evolution of petroleum biomarkers in mangrove soil (Guadeloupe). Mar. Pollut. Bull., 34: 868-874.
CrossRefDirect Link - Alikunhi, N.M., R. Narayanasamy and K. Kandasamy, 2010. Fatty acids in an estuarine mangrove ecosystem. Revista de Biologia Tropical, 58:: 577-587.
Direct Link - Nagelkerken, I., M. Dorenbosch, W.C.E.P. Verberk, E.C. de la Moriniere and G. van der Velde, 2000. Importance of shallow-water biotopes of a Caribbean bay for juvenile coral reef fishes: Patterns in biotope association, community structure and spatial distribution. Mar. Ecol. Prog. Ser., 202: 175-192.
CrossRefDirect Link - Nagelkerken, I., G. Van Der Velde, M.W. Gorissen, G.J. Meijer, T. van't Hof and C. Den Hartog, 2000. Importance of mangroves, seagrass beds and the shallow coral reef as a nursery for important coral reef fishes, using a visual census technique. Estuarine Coastal Shelf Sci., 51: 31-44.
CrossRef - Nagelkerken, I., S. Kleijnen, T. Klop, R.A.C.J. van den Brand, E.C. de la Moriniere and G. van der Velde, 2001. Dependence of Caribbean reef fishes on mangroves and seagrass beds as nursery habitats: A comparison of fish faunas between bays with and without mangroves/seagrass beds. Mar. Ecol. Progr. Ser., 214: 225-235.
CrossRefDirect Link - Napolitano, G.E., R.J. Pollero, A.M. Gayoso, B.A. Macdonald and R.J. Thompson, 1997. Fatty acids as trophic markers of phytoplankton blooms in the Bahia Blanca estuary (Buenos Aires, Argentina) and in Trinity Bay (Newfoundland, Canada). Biochem. Syst. Ecol., 25: 739-755.
CrossRefDirect Link - Newell, R.I.E., N. Marshall, A. Sasekumar and V.C. Chong, 1995. Relative importance of benthic microalgae, phytoplankton, and mangroves as sources of nutrition for penaeid prawns and other coastal invertebrates from Malaysia. Marine Biol., 123: 595-606.
Direct Link - Nyunja, J., M. Ntiba, J. Onyari, K. Mavuti, K. Soetaert and S. Bouillon, 2009. Carbon sources supporting a diverse fish community in a tropical coastal ecosystem (Gazi Bay, Kenya). Estuarine Coastal Shelf Sci., 83: 333-341.
Direct Link - Oku, H., S. Baba, H. Koga, K. Takara and H. Iwasaki, 2003. Lipid composition of mangrove and its relevance to salt tolerance. J. Plant Res., 116: 37-45.
Direct Link - Oropeza, O.A., E. Exequiel, G. Danemann, V. Valdez, J. Murray and E. Sala, 2008. Mangroves in the Gulf of California increase fishery yields. Environ. Sci. PNAS, 105: 10456-10459.
Direct Link - Orth, R.J., 1977. Effect of nutrient enrichment on growth of the seagrass Zostera marina in the Chesapeake Bay, Virginia, USA. Marine Biol., 44: 187-194.
CrossRef - Parrish, C.C., T.A. Abrajano, S.M. Budge, R.J. Helleur, E.D. Hudson, K. Pulchan and C. Ramos, 2000. Lipid and Phenolic Biomarkers in Marine Ecosystems: Analysis and Applications. In: The Handbook of Environmental Chemistry, Part D, Marine Chemistry, Wangersky, P. (Ed.). Springer, Berlin, Heidelberg, pp:193-233.
- Parrish, C.C., 1998. Lipid biogeochemistry of plankton, settling matter and sediments in Trinity Bay, Newfoundland. I. Lipid classes. Organic Geochem., 29: 1531-1545.
CrossRef - Pearson, A. and T.I. Eglinton, 2000. The origin of n-alkanes in Santa Monica Basin surface sediment: A model based on compound-specific 14 C and 13 C data. Organic Geochem., 31: 1103-1116.
CrossRef - Pempkowiak, J., W. Tylmann, A. Staniszewski and R. Gołebiewski, 2006. Lignin depolymerization products as biomarkers of the organic matter sedimentary record in 210 Pb-137 Cs-dated lake sediments. Organic Geochem., 37: 1452-1464.
CrossRef - Pepe, S., P. Scribe, J. Dagaut, M. Nallard, B. Pollet and C. Lapierre, 1995. Lignin signature in riverine suspension by capillary chromatography-mass spectrometry of thioacidolysis products. Analysis, 23: 114-116.
Direct Link - Persic, A., H. Roche and F. Ramade, 2004. Stable carbon and nitrogen isotope quantitative structural assessment of dominant species from the Vaccares Lagoon trophic web (Camargue Biosphere Reserve, France). Estuarine Coastal Shelf Sci., 60: 261-272.
Direct Link - Peterson, B.J., R.W. Howarth and R.H. Garritt, 1985. Multiple stable isotopes used to trace the flow of organic matter in estuarine food webs. Science, 227: 1361-1363.
CrossRef - Phillips, D.L. and J.W. Gregg, 2003. Source partitioning using stable isotopes: Coping with too many sources. Oecologia, 136: 261-269.
CrossRef - Prahl, F.G., J.R. Ertel, M.A. Goni, M.A. Sparrow and B. Eversmeyer, 1994. Terrestrial organic matter contributions to sediments of the Washington margin. Geochim. Cosmochim. Acta, 58: 3035-3048.
CrossRef - Primavera, J.H., 1996. Stable carbon and nitrogen isotope ratios of penaeid juveniles and primary producers in a riverine mangrove in Guimaras, Philippines. Bull. Marine Sci., 58: 675-683.
Direct Link - Rafii, Z.A., R.S. Dodd and F. Fromard, 1996. Biogeographic variation in foliar waxes of mangrove species. Biochem. Syst. Ecol., 24: 341-345.
CrossRef - Rajendran, N. and K. Kathiresan, 1998. Mangrove vegetation trap for improving fishery resources in coastal waters. Cur. Sci., 75: 429-429.
Direct Link - Rajendran, N. and K. Kathiresan, 1999. Seasonal occurrence of juvenile prawn and environmental factors in a Rhizophora mangal, southeast coast of India. Hydrobiologia, 394: 193-200.
CrossRef - Rajendran, N. and K. Kathiresan, 2004. How to increase juvenile shrimps in mangrove waters?. Weterland Ecol. Manag., 12: 179-188.
CrossRef - Rajendran, N. and K. Kathiresan, 2000. Biochemial changes in decomposing leaves of mangrove. Chem. Ecol., 17: 91-102.
CrossRef - Rajendran, N. and K. Kathiresan, 2007. Microbial flora associated with submerged mangrove leaf litter in India. Rev. Biol. Trop., 55: 393-400.
Direct Link - Ramos, C.S., C.C. Parrish, T.A.O. Quibuyen and T.A. Abrajano, 2003. Molecular and carbon isotopic variations in lipids in rapidly settling particles during a spring phytoplankton bloom. Org. Geochem., 34: 195-207.
CrossRefDirect Link - Readman, J.W., R.F.C. Mantoura, C.A. Llewellyn, M.R. Preston and A.D. Reeves, 1986. The use of pollutant and biogenic markers as source discriminants of organic inputs to estuarine sediments. Int. J Environ. Anal. Chem., 27: 29-54.
CrossRef - Rieley, G., R.J. Collier, D.M. Jones and G. Eglinton, 1991. The biogeochemistry of Ellesmere Lake, U.K-I: source correlation of leaf wax inputs to the sedimentary lipid record. Organic Geochem., 17: 901-912.
CrossRef - Robertson, A.I., 1986. Leaf-burying crabs: their influence on energy flow and export from mixed mangrove forests (Rhizophora spp.) in northeastern Australia. J. Exp. Marine Biol. Ecol., 102: 237-248.
CrossRef - Sasekumar, A., V.C. Chong, M.U. Leh and R. D'Cruz, 1992. Mangroves as a habitat for fish and prawns. Hydrobiologia, 247: 195-207.
CrossRef - Short, F.T., D.M. Burdick and J.E. Kaldy, 1995. Mesocosm experiments quantify the effects of eutrophication on eelgrass, Zostera marina. Limnol. Oceanogr., 40: 740-749.
Direct Link - Silliman, J.E. and C.L. Schelske, 2003. Saturated hydrocarbons in the sediments of Lake Apopka, Florida. Organic Geochem., 34: 253-260.
CrossRef - Smith, V.H., 2003. Eutrophication of freshwater and coastal marine ecosystems a global problem. Environ. Sci. Pollut. Res., 10: 126-139.
CrossRefDirect Link - Tanner B.R., M.E. Uhle, C.I. Mora, J.T. Kelley, P.J. Schuneman, C.S. Lane and E.S. Allen, 2010. Comparison of bulk and compound-specific δ 13 C Analyses and determination of carbon sources to salt marsh sediments using n-alkane distributions. Estuarine Coastal Shelf Sci., 86: 283-291.
Direct Link - Turner, R.E., 1977. Intertidal vegetation and commercial yield of penaeid shrimp. Trans Am. Fish Soc., 5: 411-416.
Direct Link - Turner, P.J., 1989. Romann Coins from India. Institute of Archaeology, Left Coast Press Inc., London.
Direct Link - Twilley, R.R., 1985. The exchange of organic carbon in basin mangrove forests in a southwest Florida estuary. Estuarine Coastal Shelf Sci., 20: 543-557.
CrossRefDirect Link - Twilley, R.R., R.H. Chen and T. Hargis, 1992. Carbon sinks in mangroves and their implications to carbon budget of tropical coastal ecosystems. Water Air Soil Pollut., 64: 265-288.
CrossRefDirect Link - Versteegh, G.J.M., E. Schefub, L. Dupont, F. Marret, J.S.S. Damste and J.H.F. Jansen, 2004. Taraxerol and Rhizophora pollen as proxies for tracking past mangrove ecosystems. Geochim. Cosmochim. Acta, 68: 411-422.
CrossRef - Volkman, J.K., R.B. Johns, F.T. Gillan, G.J. Perry and H.J. Bavor Jr., 1980. Microbial lipids of an intertidal sediment-I. Fatty acids and hydrocarbons. Geochimica Cosmochimica Acta, 44: 1133-1143.
CrossRefDirect Link - Volkman, J.K., S.M. Barrett, S.I. Blackburn, M.P. Mansour, E.L. Sikes and F. Gelin, 1988. Microalgal biomarkers: A review of recent research developments. Organic Geochem., 29: 1163-1179.
CrossRef - Walsh, J.J., 1991. Importance of continental margins in the marine biogeochemical cycling of carbon and nitrogen. Nature, 350: 53-55.
CrossRefDirect Link - Waycott, M., C.M. Duarte, T.J.B. Carruthers, R.J. Orth and W.C. Dennison et al., 2009. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. USA., 106: 12377-12381.
CrossRefDirect Link - Yunker, M.B., R.W. Macdonald, D.J. Veltkamp and W.J. Cretney, 1995. Terrestrial and marine biomarkers in a seasonally Ice-covered Arctic estuary-integration of multivariate and biomarker approaches. Marine Chem., 49: 1-50.
CrossRef - Zeller, D.C., 1998. Spawning aggregations: patterns of the coral trout Plectropomus leopardus (Serranidae) as determined by ultrasonic telemetry. Marine Ecol. Prog. Ser., 162: 253-263.
Direct Link - Zieman, J.C., S.A. Macko and A.L. Mills, 1984. Role of sea grasses and mangroves in estuarine food webs: temporal and spatial changes in stable isotope composition and aminoacid content during decomposition. Bull. Marine Sci., 35: 380-392.
Direct Link - Zimmerman, A.R. and E.A. Canuel, 2001. Bulk organic matter and lipid biomarker composition of the Chesapeake Bay surficial sediments as indicators of environmental processes. Estuarine Coastal Shelf Sci., 53: 319-341.
CrossRef - Abed-Navandi, D. and P.C. Dworschak, 2005. Food sources of tropical thalassinidean shrimps: A stable isotope study. Mar. Ecol. Prog. Ser., 291: 159-168.
Direct Link - Amann, R.I., W. Ludwig and K.H. Schleifer, 1995. Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol. Rev., 59: 143-169.
PubMedDirect Link - Ayukai, T., D. Miller, E. Wolanksi and S. Spagnol, 1998. Fluxes of nutrients and dissolved and particulate organic matter in two mangrove creeks in northeastern Australia. Mang. Salt Marshes., 2: 223-230.
CrossRef - Bachock, Z., P.L. Mfilinge and M. Tsuchiya, 2003. The diet of the mud clam Geloina coaxans (Mollusca, Bivalvia) as indicated by fatty acid markers in a subtropical mangrove forest of Okinawa, Japan. J. Exp. Mar. Biol. Ecol., 292: 187-197.
CrossRef - Bouillon, S., M. Frankignoulle, F. Dehairs, B. Velimirov and A. Eiler et al., 2003. Inorganic and organic carbon biogeochemistry in the Gautami Godavari estuary (Andhra Pradesh, India) during pre-monsoon: the local impact of extensive mangrove forests. Glob. Biogeochem. Cycles., 17: 1114-1127.
CrossRef - Bouillon, S., T. Moens, I. Overmeer, N. Koedam and F. Dehairs, 2004. Resource utilization patterns of epifauna from mangrove forests with contrasting inputs of local versus imported organic matter. Mar. Ecol. Pro. Ser., 278: 77-88.
Direct Link - Cambridge, M.L., A.W. Chiffings, C. Brittan, L. Moore and A.J. McComb, 1986. Loss of seagrass in Cockburn Sound, Western Australia: II. Possible causes of seagrass decline. Aquat. Bot., 24: 269-285.
CrossRef - Camilleri, J.C., 1992. Leaf-litter processing by invertebrates in a mangrove forest in Queensland. Marine Biol., 114: 139-145.
Direct Link - Canuel, E.A. and C.S. Martens, 1993. Seasonal variations in the sources and alteration of organic matter associated with recently-deposited sediments. Organic Geochem., 20: 563-577.
CrossRef - Canuel, E.A., J.E. Cloern, D.B. Ringelberg, J.B. Guckert and G.H. Rau, 1995. Molecular and isotopic tracers used to examine sources of organic matter and its incorporation into the food webs of San Francisco Bay. Limnol. Oceanography, 40: 67-81.
Direct Link - Canuel, E.A., K.H. Freeman and S.G. Wakeham, 1997. Isotopic compositions of lipid biomarker compounds in estuarine plants and surface sediments. Limnol. Oceanography, 42: 1570-1583.
Direct Link - Carmen, M.A. and A. Ablan, 2006. Genetics and the study of fisheries connectivity in Asian developing countries. Fish Res., 78: 158-168.
CrossRef - Chong, V.C., C.B. Low and T. Ichikawa, 2001. Contribution of mangrove detritus to juvenile prawn nutrition: a dual stable isotope study in a Malaysian mangrove forest. Marine Biol., 138: 77-86.
CrossRef - Costanza, R., R. D`Arge, R. de Groot, S. Farber and M. Grasso et al., 1998. The value of the world`s ecosystem services and natural capital. Nature, 387: 253-260.
Direct Link - Cranwell, P.A., G. Eglinton and N. Robinson, 1987. Lipids of aquatic organisms as potential contributors to lacustrine sediments-II. Org. Geochem., 11: 513-527.
CrossRefDirect Link - Cranwell, P.A., 1982. Lipids of aquatic sediments and sedimenting particulates. Prog. Lipid Res., 21: 271-308.
PubMedDirect Link - Danovaro, R., M. Armeni, G.M. Luna, C. Corinaldesi and A. Dell`Anno et al., 2005. Exo-enzymatic activities and dissolved organic pools in relation with mucilage development in the Northern Adriatic Sea. Sci. Total Environ., 353: 189-203.
Direct Link - Deegan, L.A. and R.H. Garritt, 1997. Evidence for spatial variability in estuarine food webs. Marine Ecol. Prog. Ser., 147: 31-47.
Direct Link - DeFlaun, M.F., J.H. Paul and D. Davis, 1986. Simplified method for dissolved DNA determination in aquatic environments. Applied Environ. Microbiol., 52: 654-659.
PubMedDirect Link - Dell`anno, A., M. Fabiano, G.C.A. Duineveld, A. Kok and R. Danovaro, 1998. Nucleic acid (DNA, RNA) quantification and RNA/DNA ratio determination in marine sediments: Comparison of spectrophotometric, fluorometric and high performance liquid chromatography methods and estimation of detrital DNA. Applied Environ. Microbiol., 64: 3238-3245.
Direct Link - Dittmar, T. and R.J. Lara, 2001. Do mangroves rather than rivers provide nutrients to coastal environments south of the Amazon River? Evidence from long-term flux measurements. Marine Ecol. Prog. Ser., 213: 67-77.
Direct Link - Dittmar, T. and R.J. Lara, 2001. Molecular evidence for lignin degradation in sulfate reducing mangrove sediments (Amazonia, Brazil). Geochim. Cosmochim Acta, 65: 1417-1428.
CrossRef - Dittmar, T., R.J. Lara and G. Kattner, 2001. River or mangrove? Tracing major organic matter sources in tropical Brazilian coastal waters. Marine Chem., 73: 253-271.
Direct Link - Dittmar, T., N. Hertkorn, G. Kattner and R.J. Lara, 2006. Mangroves, a major source of dissolved organic carbon to the oceans. Global Biogeochem. Cycles, 20: 1012-1026.
CrossRef - Dodd, R.S., F. Fromard, Z.A. Rafii and F. Blasco, 1995. Biodiversity among west African Rhizophora: foliar wax chemistry. Biochem. Syst. Ecol., 23: 859-868.
CrossRef - Dodd, R.S., Z.A. Rafii, F. Fromard and F. Blasco, 1998. Evolutionary diversity among Atlantic coast mangroves. Acta Oecol., 19: 323-330.
CrossRefDirect Link - Donaldson, A.L., 2001. Lignification and lignin topochemistry-ultrastructural view. Phytochemistry, 57: 859-873.
PubMedDirect Link - Farella, N., M. Lucotte, P. Louchouarn and P. Roulet, 2001. Deforestation modifying terrestrial organic transport in the Rio Tapajes, Brazilian Amazon. Org. Geochem., 32: 1443-1458.
CrossRefDirect Link - Fisher, E., F. Oldfield, R. Wake, J. Boyle, P. Appleby and G.A. Wolff, 2003. Molecular marker record of land use change. Organic Geochem., 34: 105-119.
CrossRef - Fernandes, M.B. and M.A. Sicre, 2000. The importance of terrestrial organic carbon inputs on Kara Sea shelves as revealed by n-alkanes, OC and ∂ 13 C values. Organic Geochem., 31: 363-374.
CrossRef - Ficken, K.J., B. Li, D.L. Swain and G. Eglinton, 2000. An n-alkane proxy for the sedimentary inputs of submergedfloating freshwater aquatic macrophytes. Organic Geochem., 31: 745-749.
CrossRef - Fourqurean, J.W. and M.B. Robbins, 1999. Florida Bay: A history of recent ecological changes. Estuaries, 22: 345-357.
CrossRef - Hall, D., S.Y. Lee and T. Meziane, 2006. Fatty acids as trophic tracers in an experimental estuarine food chain: Tracer transfer. J. Exp. Mar. Biol. Ecol., 336: 42-53.
CrossRef - Hatcher, P.G., M.A. Nanny, R.D. Minard, S.D. Dible and D.M. Carson, 1995. Comparison of two thermochemolytic methods for the analysis of lignin in decomposing gymnosperm wood: The CuO oxidation method and the method of thermochemolysis with tetramethylammonium hydroxide (TMAH). Organic Geochem., 23: 881-888.
CrossRef - Hazel, J.R., E.E. Williams, R. Livermore and N. Mozingo, 1991. Thermal adaptation in biological membranes: Functional significance of changes in phospholipids molecular species composition. Lipids, 26: 277-282.
CrossRef - Hedges, J.I., J.R. Ertel and E.B. Leopold, 1982. Lignin geochemistry of a late Quaternary sediment core from Lake Washington. Geochim. Cosmochim. Acta, 46: 1869-1877.
CrossRef - Hedges, J.I. and D.C. Mann, 1979. The characterization of plant tissues by their lignin oxidation products. Geochim. Cosmochim. Acta, 43: 1803-1807.
CrossRef - Hedges, J.I. and A. van Geen, 1982. A comparison of lignin and stable carbon isotope composition in quaternary sediments. Marine Chem., 11: 43-53.
CrossRef - Hemminga, M.A., F.J. Slim, J. Kazungu, G.M. Ganssen, J. Nieuwenhuize and N.M. Kruyt, 1994. Carbon outwelling from a mangrove forest with adjacent seagrass beds and coral reefs (Gazi Bay, Kenya). Marine Ecol. Prog. Ser., 106: 291-301.
Direct Link - Hernes P.J. and J.I. Hedges, 2004. Tannin signatures of barks, needles, leaves, cones and wood at the molecular level. Geochim. Cosmochim. Acta, 5: 1293-1307.
CrossRefDirect Link - Hernes, P.J., R. Benner, G.L. Cowie, M.A. Goni, B.A. Bergamaschi and J.I. Hedges, 2001. Tannin diagenesis in mangrove leaves from a tropical estuary: A novel molecular approach. Geochim. Cosmochim. Acta, 65: 3109-3122.
CrossRefDirect Link - Hiddink, J.G., 2003. Modelling the adaptive value of intertidal migration and nursery use in the bivalve Macoma balthica. Marine Ecol. Prog. Ser., 252: 173-185.
Direct Link - Hu, F.S., J.I. Hedges, E.S. Gordon and L.B. Brubaker, 1999. Lignin biomarkers and pollen in postglacial sediments of an Alaskan lake. Geochimica et Cosmochimica Acta, 63: 1421-1430.
CrossRef - Huang, W.Y. and W.G. Meinschein, 1978. Sterols in sediments from Baffin Bay, Texas. Geochim. Cosmochim. Acta, 42: 1391-1396.
CrossRef - Huang, W.Y. and W.G. Meinschein, 1979. Sterols as ecological indicators. Geochim. Cosmochim. Acta, 43: 739-745.
CrossRef - Ishiwatari, R. and M. Uzaki, 1987. Diagenetic changes of lignin compounds in a more than 0.6 million-year-old lacustrine sediments. Geochim. Cosmochim. Acta, 51: 321-328.
CrossRef - Jaffe, R., R. Mead, M.E. Hernandez, M.C. Perlaba and O.A. DiGuida, 2001. Origin and transport of sedimentary organic matter in two subtropical estuaries: A comparative, biomarker-based study. Organic Geochem., 32: 507-526.
CrossRef - Jennerjahn, T.C. and V. Ittekkot, 2002. Relevance of mangroves for the production and deposition of organic matter along tropical continental margins. Aturwissenschaften, 89: 23-30.
PubMedDirect Link - Kathiresan, K. and B.L. Bingham, 2001. Biology of mangroves and mangrove ecosystems. Adv. Mar. Biol., 40: 81-251.
CrossRefDirect Link - Kathiresan, K. and N. Rajendran, 2002. Fishery resources and economic gain in three mangroves areas on the southeast coast of India. Fish. Manage. Ecol., 49: 277-283.
Direct Link - Kay, J.J. and E. Schneider, 1994. Embracing complexity, the challenge of the ecosystem approach. Alternatives, 20: 32-38.
Direct Link - Keeley, J.E. and D.R. Sandquist, 1992. Carbon: Freshwater plants: Commissioned review. Plant Cell Environ., 15: 1021-1035.
Direct Link - Kharlamenko, V.I., N.V. Zhukova, S.V. Khotimchenko, V.I. Svetashev and G.M. Kamenev, 1995. Fatty acids as markers of food sources in a shallow-water hydrothermal ecosystem (Kraterna Bight, Yankich Island, Kurile Islands). Mar. Ecol. Prog. Ser., 120: 231-241.
Direct Link - Kharlamenko, V.I., S.I. Kiyashko, A.B. Imbs and D.I. Vyshkvartzev, 2001. Identification of food sources of invertebrates from the seagrass Zostera marina community using carbon and sulfur stable isotope ratio and fatty acid analyses. Mar. Ecol. Prog. Ser., 220: 103-117.
CrossRef - Killops, S.D. and N.L. Frewin, 1994. Triterpenoid diagenesis and cuticular preservation. Organic Geochem., 21: 1193-1209.
CrossRef - Kneib, R.T. and A.E. Stiven, 1980. Stable carbon isotope ratios in Fundulus heteroclitus (L.) muscle tissue and gut contents from a North Carolina Spartina marsh. J. Exp. Marine Biol. Ecol., 46: 89-98.
CrossRef - Kwak, T.J. and J.B. Zedler, 1997. Food web analysis of southern California coastal wetlands using multiple stable isotopes. Oecologia, 110: 262-277.
Direct Link - Law, B.S. and C.R. Dickman, 1998. The use of habitat mosaics by terrestrial vertebrate fauna: Implications for conservation and management. Biodiversity Conserv., 7: 323-333.
CrossRef - Lee, S.Y., 2004. Relationship between mangrove forests and prawn production: a re-evaluation. Marine Biol., 145: 943-949.
CrossRef - Lepoint, G., P. Dauby and S. Gobert, 2004. Applications of C and N stable isotopes to ecological and environmental studies in seagrass ecosystems. Marine Pollut. Bull., 49: 887-891.
CrossRef - Li, B., X. Zhang and R. Su, 2002. An investigation of thermal degradation and charring of larch lignin in the condensedphase: The effects of boric acid, guanyl urea phosphate, ammonium dihydrogen phosphate and ammonium polyphosphate. Polymer Degradation Stability, 75: 35-44.
CrossRef - Loneragan, N.R., S.E. Bunn and D.M. Kellaway, 1997. Are mangroves and seagrasses sources of organic carbon for penaeid prawns in a tropical Australian estuary? A multiple stable-isotope study. Marine Biol., 130: 289-300.
CrossRef - Lugo, A.E. and S.C. Snedaker, 1974. The ecology of mangroves. Ann. Rev. Ecol. Syst., 5: 39-64.
Direct Link - Maie, N., O. Pisani and R. Jaffe, 2008. Mangrove tannins in aquatic ecosystems: Their fate and possible influence on dissolved organic carbon and nitrogen cycling. Limnol. Oceanograpy, 53: 160-171.
Direct Link - Marchand, C., J.R. Disnar, E. Lallier-Verges and N. Lottier, 2005. Early diagenesis of carbohydrates and lignin in mangrove sediments subject to variable redox conditions (French Guiana). Geochim. Cosmochim. Acta, 69: 131-142.
CrossRefDirect Link - Mead, R., Y. Xu, J. Chong and R. Jaffe, 2005. Sediment and soil organic matter source assessment as revealed by the molecular distribution and carbon isotopic composition of n-alkanes. Organic Geochem., 36: 363-370.
CrossRef - Meziane, T. and M. Tsuchiya, 2000. Fatty acids as tracers of organic matter in the sediment and food web of a mangrove/intertidal flat ecosystem, Okinawa, Japan. Mar. Ecol. Progr. Ser., 200: 49-57.
Direct Link - Micic, M., J. Orbulescu, K. Radotic, M. Jeremic, G. Sui, Y. Zheng and R.M. Leblanc, 2002. ZLDHP lignin model compound at the air water interface. Biophys. Chem., 99: 55-62.
CrossRef - Miltner, A. and K. Emeis, 2001. Terrestrial organic matter in surface sediments of the Balic Sea, northwest Europe, as determined by CuO oxidation. Geochimica Cosmochimica Acta, 65: 1285-1299.
CrossRef