
Leonce Kouakanou
Laboratory of Biology and Molecular Typing in Microbiology, Faculty of Sciences and Techniques, University of Abomey-Calavi, Benin
LiveDNA: 229.23499
Constance Agbemelo-Tsomafo
Noguchi Memorial Institute for Medical Research, Department of Animal Experimentation, University of Ghana, Ghana
Jesca Nakayima
National Livestock Resources Research Institute (NaLIRRI), Tororo, Uganda
LiveDNA: 256.23862
Taiwo Crossby Omotoriogun
Biotechnology Unit, Department of Biological Sciences, Elizade University, P.M.B. 002, Ilara-Mokin, Ondo State, Nigeria
Dieter Kabelitz
Institute of Immunology, University of Kiel, Kiel, Germany
Lamine Baba-Moussa
Laboratory of Biology and Molecular Typing in Microbiology, Faculty of Sciences and Techniques, University of Abomey-Calavi, Benin
Current Research in Poultry Science
Year: 2018 | Volume: 8 | Issue: 1 | Page No.: 1-8







ABSTRACT
Background and Aim: The use of microscopy-based and polymerase chain reaction (PCR)-based methods have offered considerable insights in detecting avian blood malaria infection in domestic and wild birds. The infection has a significant impact on the immunity of birds. However, some observations concerning the role of immune system in controlling the infection continue to question the extent of immune factors involved. To address this, the current study hypothesized that avian malaria infection may influence the humoral response of domestic birds. Materials and Methods: The prevalence of avian malaria parasites (both Plasmodium and Haemoproteus spp.) in free-ranging domestic birds from Uganda was evaluated using PCR and the level of Immunoglobulin Y (IgY) antibody in malaria-infected and uninfected birds was determined using enzyme-linked immunosorbent assay (ELISA). Result: The results showed that 10 (15.15%) of 66 individually tested birds were infected with avian malaria parasites. Interestingly, an increase in the level of IgY associated with the infection was found. Of note, the uninfected birds exhibited a consistent level of IgY, however, less than in malaria-positive birds. It is likely that avian malaria-independent factors may have been involved in this induction in uninfected birds. Conclusion: The data obtained in this study suggest that avian malaria infections influence the production of IgY in domestic birds, therefore indicating the potential of IgY as an immune biomarker for screening avian malaria infection in domestic birds. However, these observations are subject to further investigation with larger sample size.
PDF Abstract XML References Citation
How to cite this article
Leonce Kouakanou, Constance Agbemelo-Tsomafo, Jesca Nakayima, Taiwo Crossby Omotoriogun, Dieter Kabelitz and Lamine Baba-Moussa, 2018. Total Immunoglobulin Y Detection in Avian Malaria-infected Domestic Birds from Uganda. Current Research in Poultry Science, 8: 1-8.
URL: https://scialert.net/abstract/?doi=crpsaj.2018.1.8
URL: https://scialert.net/abstract/?doi=crpsaj.2018.1.8
INTRODUCTION
Despite recent advances in the strategies to control avian malaria, the disease is still at high risk in poultry production systems1,2. Avian malaria is caused by protozoan parasites including haemosporidians (Plasmodium spp. and Haemoproteus spp., both often referred to as "avian malaria parasites")3. Birds are present in a wide range of ecosystem distribution globally but African ecosystem harbor one of the highest continental bird diversities. As a consequence, the malaria parasites of these birds appear to be as diverse as their avian hosts4,5. Several studies have highlighted the diversity of haemosporidians in birds using microscopy, however, studies from Africa6,7 identified haemosporidians parasites in 27 avian families with the highest prevalence from Uganda.
Although the pathogenicity of many haemosporidians species in wild birds is not completely understood, many cases of mortality have been reported in domestic birds8,9. In poultry industry, avian malaria is a common cause of low quality meat, poor egg production and high mortality9. Clinical signs associated with avian malaria include a pale crest, green stools, anorexia, elevated body temperature and anemia. These symptoms are not necessarily due to malaria infection and clinical signs may vary from no clinical signs to a severe phase and cause death with mortality rate higher than 80%8,10. Native chickens are more resistant to infection than commercial chickens11. Nevertheless, some infected adult chickens may develop asymptomatic infections12.
For the rapid detection of the infection in birds and to control the spread of the disease, assays combining microscopy and PCR have been established and widely used. Moreover, molecular studies using gene sequence analysis have offered considerable support for the classification of the majority of genera and subgenera of avian haemosporidian parasites and have provided additional information on the phylogenetic relationships between characterized genera13,14. Although PCR assays and sequencing have provided a reliable diagnostic tool for molecular epidemiological studies of avian malaria infections in domestic and wild birds, little is still known about why some birds are more susceptible to infection, while others are resistant and/or tend to be more "asymptomatically" parasitized.
The immune system has over decades attracted great deal of interest in functional ecology15,16. Measurements of immune response against infection are becoming routine in many ecological studies17, requiring the use of simple standardized methods (e.g., ELISA) for quantifying levels of different components (e.g., antibodies) of the immune system. Antibodies (also termed immunoglobulins (Igs)) are a class of proteins that are associated with the adaptive immune response and are only expressed in jawed vertebrates (including birds)18. Numerous studies have demonstrated that birds express IgD (except in chickens, ducks, penguin and zebra finch), IgM, IgA and IgY isotypes subclasses19-21. Of particular relevance here, IgY antibody has been associated with evidences of protection towards different infections as described in several studies, suggesting the use of IgY antibody as immunodiagnostic and immunotherapeutic tool22-24. With regard to malaria infection, pioneering studies by Graczyk et al.25,26 described immunological tools to investigated anti-Plasmodium spp. humoral response of birds. Using ELISA, the authors reported a correlation between antibody titer to Plasmodium spp. and the resistance of birds to plasmodial parasites. Moreover, another study addressing the extent of Plasmodium exposure in Galapagos penguins, showed an association between higher seroprevalence of malarial antibodies and low Plasmodium-induced mortality27.
Seroprevalence detection using ELISA-based method has been developed for diagnosing malarial infections in a variety of bird species. However, estimating the total concentration of IgY antibody in avian malaria infection remains to be investigated. Addressing this, the present study examined an increase of total IgY antibody concentration in malaria-infected domestic birds. However, even though less than in malaria-infected birds, there was also a consistent production of IgY in malaria-uninfected birds. Together, these data suggested that malaria influences humoral response of domestic birds but malaria-independent factors seemingly may underpin the production of IgY in uninfected birds.
MATERIALS AND METHODS
Collection of blood samples: The blood samples used in the present study were collected from North-western Uganda at four districts Adjumani (Latitude: N 3.37, Longitude: E 31.78), Moyo (Latitude: N 3.65, Longitude: E 31.72), Yumbe (Latitude: N 3.46, Longitude: E 31.24) and Koboko (Latitude: N 3.41, Longitude: E 30.95) in October 2015. Blood samples were taken from free-ranging domestic birds i.e., chickens, duck, turkey and guinea fowl. All the birds were bled on the brachial vein (venipuncture), from which 5-50 μL of blood was spotted onto sheets of Whatman n°1 (Whatman, Maidstone, UK) filter paper. Spots were allowed to dry at ambient temperature and stored in dark self-sealing plastic bags, the innermost of which contained silica desiccant (Sigma). Filter papers were stored at -20°C, where the level of antibodies may remain stable for many years28,29.
DNA extraction: Total DNA was extracted using the chelex-100 resin-based DNA extraction method as previously described12 with some modifications. In brief, a small disc of dried blood spots was cut and transferred into 1.5 mL collection tube (Eppendorf) containing 1 mL of 10% Saponin in phosphate buffered saline (PBS). After an overnight incubation at 4°C, the suspension was centrifuged and the PBS/Saponin was poured and then replaced with 200 μL of 20% of chelex-100 resin in deionized water, heated at 100°C for 8 min and vortexed every 2 min. The suspension was centrifuged for 1 min to pellet the chelex-100 resin and cell debris. The supernatant containing the DNA was carefully collected and the concentration of DNA as well as the ratio A260/280 for each sample was measured using a NanodropTM spectrophotometer (Thermo Scientific, USA). DNA samples were stored at -20°C until molecular testing.
PCR-based detection of avian malaria parasites gene fragments: The extracted DNA was used in nested Polymerase Chain Reaction (nPCR) to amplify DNA fragments according to method described by Waldenstrom et al.30. The presence of extracted DNA was first assessed by gel electrophoresis using 2 μL of the extract on a 1.5% agarose gel, post stained with Diamond nucleic acid dye and visualized under ultraviolet light (UV). Samples were then screened for the presence of Plasmodium and Haemoproteus using the nPCR method, amplifying a 478 bp fragment of the mitochondrial cytochrome b gene. The PCR reactions were performed, in two separate rounds. First-round primers were HaemNF (5-CATATATTAAGAGAATTATGGAG-3) and HaemNR2 (5-AGAGGTGTAGCATATCTATCTAC-3). The first reaction contained 0.5 μL of 50-90ng genomic DNA, 0.4 μM of each primer, 7.5 μL of Go taq® G2 Hot Start Green Master Mix, 2x (Promega corporation, USA) and 5.8 μL of nuclease free water in a total volume of 15 μL reaction mixture. The cycling condition consisted of a 3 min 94°C initial denaturation, followed by 35 cycles of 94°C for 30 sec, 55°C for 30 sec and 72°C for 45 sec, ending with an elongation step of 72°C for 10 min. In the second PCR round, primers HaemF (5-ATGGTGCTTTCGATATATGCATG-3) and HaemR2 were used (5-GCATTATCTGGATGTGATAATGGT-3). The reaction was in a total volume of 30 μL containing 1 μL of the PCR product from the first round as template, 0.4 μM of each primer, 15 μL of Go taq® G2 Hot Start Green Master Mix, 2x (Promega corporation, USA) and 11.6 μL of nuclease free water. The PCR cycling condition was the same as used in the first round. The PCR products (5 μL) from the second round were run on 1.5% agarose gel and post stained with Diamond nucleic acid dye solution for 20-30 min and visualized under UV light.
Elution of serum from dried blood spots: Plastic bags containing blood spots were allowed to return to ambient temperature before opening. Individual discs of dried blood spots were punched manually into 1.5 mL collection tube (Eppendorf). The blood was eluted out in 200 μL phosphate buffered saline (PBS) containing 0.4% Tween20 overnight at 4°C. The tubes were centrifuged and the supernatants containing the eluted serum proteins form the "master" from which dilutions were made for subsequent ELISA testing.
Antibody IgY determination by sandwich ELISA: Total IgY ELISAs were performed using Chicken IgY ELISA kit (Immunology Consultants Laboratory, Inc, Portland, USA) according to the manufacturer’s recommendations. All the reagents used in this test are included in the ELISA kit. Briefly, the eluted serum was diluted 1:6250, 1:12500, 1:25000 and 1:50000 in diluent buffer and 100 μL of the diluted serum and standards were added in duplicate to a 96-well microtiter plate pre-coated with anti-IgY antibody. The plate was incubated in dark at room temperature (RT) for 30 min. After the incubation, unbound proteins were removed by washing the plate four times and 100 μL of anti-IgY-HRP antibody, diluted 1:100 with diluent, was added per well, incubated for 30 min RT. Plate was again washed and 100 μL of tetramethylbenzidine (TMB)-peroxidase substrate solution was added into each well and the plate was incubated in the dark at RT for precisely 10 min. the reaction was stopped with 100 μL of 0.3 M Sulfuric acid. The plate was immediately read at 450 nm using Infinite MT200 microplate reader (TECAN, Switzerland) calibrated according to the manufacturer’s specifications.
Statistical analysis: Data were entered in Excel sheet 2007 (Microsoft) and were analyzed (where indicated) using Prism 6.01 (GraphPad, La Jolla, USA). Man-Whitney U-test was used to evaluate difference in IgY concentration values between malaria-positive and negative samples. The p<0.05 was considered statistically significant.
RESULTS
Prevalence of avian malaria parasites: Domestic birds samples examined in this study were composed of chickens, duck, turkey and guinea fowl from four different districts located in North-Western Uganda.
 | |
| Fig. 1: | nPCR-based detection of avian malaria parasites DNA. DNA was extracted from filter paper dried blood sample by the chelex-100 resin method. The DNA samples were used as templates for nPCR reaction with primers specific for parasite gene fragments, with amplicons being analyzed by standard agarose gel electrophoresis (Mat and Meth section) |
Lanes 1-18: Malaria parasites-negative samples, Lanes 19-20: Malaria parasites-positive samples, Lane L: Ladder | |
| Table 1: | Occurrence of avian malaria parasites in the domestic birds |
 | |
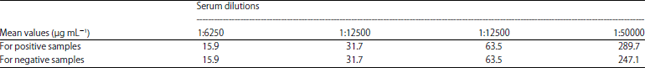
| Table 2: | Optimization of serum dilution |
 | |
The prevalence of avian malaria parasites was determined using nPCR (Fig. 1). As the primers used in this nested PCR amplify both Plasmodium and Haemoproteus spp., the term avian malaria parasites considered in this report includes both parasites. As shown in Table 1, of 66 individuals tested, 10 (15.15%) were positive to avian malaria parasites.
Optimization of serum dilution for ELISA: As this study aimed to detect the level of IgY antibody toward malaria parasites infection in domestic birds, ten malaria-positive samples (M+) and nine malaria-negative samples (M-) were tested to determine their serum level of IgY antibody. First used different serum dilutions (titrated from 1:6250 to 1:50000) in a standardized ELISA (Table 2). The standard curve was fitted to the absorbance observed for the standards value by a second order polynomial curve using Excel (Microsoft). Test sample values were interpolated from the standard curve and were subsequently corrected for sera dilution factor to finally obtain the IgY concentration in the original samples.
The standardization of ELISA for IgY detection in birds showed optimum result at 1:50000 serum dilution. In contrast, no changes in IgY between positive and negative samples were observed using 1:6250, 1:12500 and 1:25000 dilutions. Thus, in the experimental settings, the dilution of serum at 1:50000 was suitable for the IgY quantification and was considered for further analysis.
Impact of malaria infection on the IgY production in birds: To investigate the impact of malaria infection on the humoral response of birds, the level of IgY in malaria-infected birds in comparison to the uninfected birds was determined.
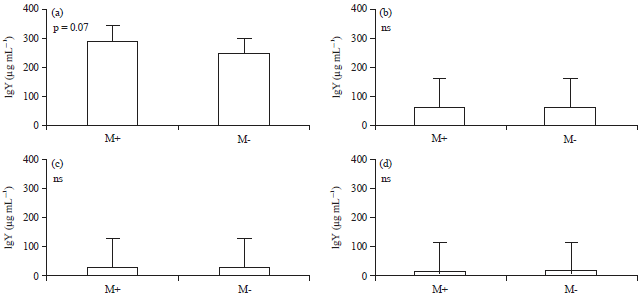 | |
| Fig. 2(a-d): | Avian malaria infection modifies IgY production of domestic birds. Serum samples from malaria positive (M+) birds (n = 10) and malaria-negative (M-) birds (n = 9) were diluted at (a) 1:50000, (b) 1:25000, (c) 1:12500 and (d) 1:6250. Thereafter, diluted sera were analyzed for the detection of total IgY antibody using sandwich ELISA |
Bars indicate the antibody level as Mean±SD in each group. Statistical comparison between M+ and M- was based on Mann-Whitney U-test. ns: Not significant | |
| Table 3: | Range concentration of IgY detection in malaria-positive vs negative birds |
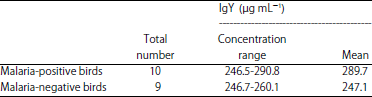 | |
Results showed that the range in IgY concentration (μg mL–1) for the positive sample at 1:50000 dilution was 246.5-290.8 with a Mean±SD of 289.7±20.9, while IgY concentration for the group of negative samples ranged from 246.7-260.1 with a mean value±SD of 247.1±16.2 (Table 3). Interestingly, although statistically not significant (as determined by Mann-Whitney Unpaired-test), serum of malaria positive samples displayed a slight increase in IgY production when compared to the uninfected birds group (Fig. 2). These data suggested that avian malaria infection induces an elevated production of IgY antibody in domestic birds.
DISCUSSION
This study aimed to measure the production of total IgY antibody in response to avian malaria infection in free-ranging domestic birds from Uganda. The most interesting finding is that malaria-infected birds exhibit a slight higher level of IgY antibody when compared to the uninfected birds. This result was obtained with an especially high dilution factor (1:50000) of the serum, possibly due to the binding specificity property of the microplate. Seroprevalence of malaria in different bird species have been reported by various studies27,31-33. For example, a study on Galapagos penguins Spheniscus mendiculus showed elevated seroprevalence of anti-malaria antibodies, despite low detection of Plasmodium spp. DNA using PCR. This suggests high exposure to the parasite and low Plasmodium-induced mortality27 and thus, a role of host immunity in controlling the infection. Similarly, Jarvi et al.33 reported that serological methods for detecting chronic malarial infections were significantly more sensitive than PCR-based methods and microscopy. These studies also emphasize the necessity of combining different strategies to measure the real extent of Plasmodium exposure and infection.
Domestic birds samples examined in the present study showed few avian malaria parasite infections. However, the birds were trapped in several habitats recognized as abundant midges for mosquitoes as described6. According to Pattaradilokrat et al.12, the chelex-100 resin extraction method, in comparison to the phenol/chloroform method, was found to be a reliable method for the extraction of DNA of avian malaria parasite P. gallinaceum from dried blood spot. Moreover, the nPCR assay presented in this study has been shown to be highly specific and reliable method for assessing prevalence of the haemosporidian blood parasites Haemoproteus spp. and Plasmodium spp. from avian blood30. Hence, the low prevalence of avian malaria infections observed here may not be parasites detection method-dependent.
As one of the possible explanation for this unexpected result is that PCR amplifies haemosporidian parasite DNA in the circulating blood, regardless of the parasite’s life cycle and therefore may not be detecting gametocytes34. Moreover, a previous study in Uganda reported an unexpected low prevalence of Plasmodium and Haemoproteus spp. in birds but a predominance of Leucocytozoon spp., suggesting that Leucocytozoon may be the most common hematozoan in chickens from Uganda35. This study showed a consistent production of IgY in uninfected birds (Table 1, Fig. 2), however, less than in malaria-positive birds. This could suggest that the production of IgY detected in this study is not completely exclusive to malaria infection but could also be the result of other infections which might occurred in domestic birds. However, more investigations are needed to identify the malaria-independent factor (s) responsible for the production of IgY in uninfected birds.
Antibodies play an important role in the initial recognition of foreign particles and they support subsequent defense by the complement cascade and the acquired humoral response36,37. Therefore, organisms may benefit by maintaining a minimum level of antibodies as these molecules likely play a crucial role in terms of earlier detection of parasites38. With regard to the protective effect of IgY, several studies have reported the effective protection against pathogens such as Salmonella enteritidis, Salmonella e. typhimurium, Campylobacter jejuni and Enterotoxigenic Escherichia coli (ETEC)22-24. In addition, a previous study found that, in in vitro experimental system, chicken IgY-specific antibodies directed against the E. coli enteropathogenic BfpA virulence have also been shown to inhibit, in a dose-dependent manner, the virulence factor-induced apoptosis of Vera cells39. However, it has not been straightforward to demonstrate an association between antibody production and protection of domestic birds from malaria infection. Birds infected with malaria show clinical signs such as pale crest, green stools, ruffled feathers, anorexia, cerebral capillary blockade9,40. Although, these symptoms are not exclusively specific to malaria, the disease may run a very acute phase and cause death within the second or third week8,10. Nevertheless, some infected adult chickens may develop asymptomatic infection, resulting in misdiagnosis12 or the result of chronic infection. In this study, domestic birds infected with malaria (as determined by nPCR) seem in "good health" and do not show any sign of the aforementioned malaria symptoms (data not shown) but interestingly, they display an increase in IgY antibody production. Taken together, this finding suggests the increase in IgY production as an indicator of survival in infected birds. However, this observation remains to be further verified with larger sample size.
CONCLUSION
In conclusion, the present study confirmed that avian malaria infection occurred in domestic birds from different free-ranging poultry in Uganda. Interestingly, an increase in IgY production of birds infected with avian malaria parasites was detected. These findings highlight the importance of using IgY antibodies either for immunodiagnostic or for passive immunization against avian malaria infections in domestic birds.
SIGNIFICANCE STATEMENT
Avian malaria infection has a significant impact on the immunity of birds. Data obtained in this study indicate that avian malaria-infected domestic birds display an increase in IgY level and do not show any sign of avian malaria symptoms. These findings suggest that avian malaria infections influence the production of IgY in domestic birds, therefore indicating the potential of IgY as an immune biomarker for screening avian malaria infection in domestic birds.
ACKNOWLEDGMENTS
This study was supported by a Collaborative Research Grant from International Foundation for Science (IFS) to LK (J/5722-1), CA-T (J/5721-1), JN (J/5723-1) and TCO (J/5724-1). We thank the Uganda-based team for sample collection. We also thank Dr. Ulrich F. Prodjinotho (Institut für Medizinische Mikrobiologie, Immunologie und Hygiene, Technische Universität München) for critical reading of the manuscript.
REFERENCES
- Ruff, M.D., 1999. Important parasites in poultry production systems. Vet. Parasitol., 84: 337-347.
CrossRefDirect Link - Tasai, S., T. Saiwichai, M. Kaewthamasorn, S. Tiawsirisup, P. Buddhirakkul, S. Chaichalotornkul and S. Pattaradilokrat, 2017. Artesunate-tafenoquine combination therapy promotes clearance and abrogates transmission of the avian malaria parasite Plasmodium gallinaceum. Vet. Parasitol., 233: 97-106.
CrossRefDirect Link - Clark, N.J., S.M. Clegg and M.R. Lima, 2014. A review of global diversity in avian haemosporidians (Plasmodium and Haemoproteus: Haemosporida): New insights from molecular data. Int. J. Parasitol., 44: 329-338.
CrossRefDirect Link - Lauron, E.J., C. Loiseau, R.C.K. Bowie, G.S. Spicer, T.B. Smith, M. Melo and R.N.M. Sehgal, 2015. Coevolutionary patterns and diversification of avian malaria parasites in African sunbirds (Family Nectariniidae). Parasitology, 142: 635-647.
CrossRefDirect Link - Lutz, H.L., W.M. Hochachka, J.I. Engel, J.A. Bell and V.V. Tkach et al., 2015. Parasite prevalence corresponds to host life history in a diverse assemblage of afrotropical birds and haemosporidian parasites. PloS One, Vol. 10, No. 4.
CrossRefDirect Link - Valkiunas, G., R.N.M. Sehgal, T.A. Iezhova and T.B. Smith, 2005. Further observations on the blood parasites of birds in Uganda. J. Wildlife Dis., 41: 580-587.
CrossRefDirect Link - Sehgal, R.N.M., H.I. Jones and T.B. Smith, 2005. Blood parasites of some West African rainforest birds. J. Vet. Med. Sci., 67: 295-301.
PubMedDirect Link - Permin, A. and J. Juhl, 2002. The development of Plasmodium gallinaceum infections in chickens following single infections with three different dose levels. Vet. Parasitol., 105: 1-10.
CrossRefDirect Link - Williams, R.B., 2005. Avian malaria: Clinical and chemical pathology of Plasmodium gallinaceum in the domesticated fowl Gallus gallus. Avian Pathol., 34: 29-47.
CrossRefDirect Link - De Matos Macchi, B., J.A.S. Quaresma, A.M. Herculano, M.E. Crespo-Lopez, R.A. DaMatta and J.L.M. do Nascimento, 2010. Pathogenic action of Plasmodium gallinaceum in chickens: Brain histology and nitric oxide production by blood monocyte-derived macrophages. Vet. Parasitol., 172: 16-22.
CrossRefDirect Link - Sohsuebngarm, D., J. Sasipreeyajan, S. Nithiuthai and N. Chansiripornchai, 2014. The efficacy of artesunate, chloroquine, doxycycline, primaquine and a combination of artesunate and primaquine against avian malaria in broilers. J. Vet. Med. Sci., 76: 813-817.
CrossRefDirect Link - Pattaradilokrat, S., W. Tiyamanee, P. Simpalipan, M. Kaewthamasorn, T. Saiwichai, J. Li and P. Harnyuttanakorn, 2015. Molecular detection of the avian malaria parasite Plasmodium gallinaceum in Thailand. Vet. Parasitol., 210: 1-9.
CrossRefDirect Link - Valkiunas, G., T.A. Iezhova, C. Loiseau, T.B. Smith and R.N. Sehgal, 2009. New malaria parasites of the subgenus Novyella in African rainforest birds, with remarks on their high prevalence, classification and diagnostics. Parasitol. Res., 104: 1061-1077.
CrossRefDirect Link - Martinsen, E.S., I. Paperna and J.J. Schall, 2006. Morphological versus molecular identification of avian Haemosporidia: An exploration of three species concepts. Parasitology, 133: 279-288.
CrossRefDirect Link - Sheldon, B.C. and S. Verhulst, 1996. Ecological immunology: Costly parasite defences and trade-offs in evolutionary ecology. Trends Ecol. Evol., 11: 317-321.
CrossRefDirect Link - Zuk, M., 1996. Disease, endocrine‐immune interactions and sexual selection. Ecology, 77: 1037-1042.
CrossRefDirect Link - Norris, K. and M.R. Evans, 2000. Ecological immunology: Life history trade-offs and immune defense in birds. Behav. Ecol., 11: 19-26.
CrossRefDirect Link - Flajnik, M.F., 2002. Comparative analyses of immunoglobulin genes: Surprises and portents. Nat. Rev. Immunol., 2: 688-698.
CrossRefDirect Link - Han, B., H. Yuan, T. Wang, B. Li and L. Ma et al., 2016. Multiple IgH isotypes including IgD, subclasses of IgM and IgY are expressed in the common ancestors of modern birds. J. Immunol., 196: 5138-5147.
CrossRefDirect Link - Han, B., Y. Li, H. Han, Y. Zhao, Q. Pan and L. Ren, 2017. Three IgH isotypes, IgM, IgA and IgY are expressed in Gentoo penguin and zebra finch. PloS One, Vol. 12, No. 4.
CrossRefDirect Link - Lundqvist, M.L., D.L. Middleton, S. Hazard and G.W. Warr, 2001. The immunoglobulin heavy chain locus of the duck: Genomic organization and expression of D, J and C region genes. J. Biol. Chem., 276: 46729-46736.
CrossRefPubMedDirect Link - Chalghoumi, R., Y. Beckers, D. Portetelle and A. Thewis, 2009. Hen egg yolk antibodies (IgY), production and use for passive immunization against bacterial enteric infections in chicken: A reviewe. Biotechnol. Agron. Soc. Environ., 13: 295-308.
Direct Link - Rahimi, S., Z.M. Shiraz, T.Z. Salehi, M.A.K. Torshizi and J.L. Grimes, 2007. Prevention of Salmonella infection in poultry by specific egg-derived antibody. Int. J. Poult. Sci., 6: 230-235.
CrossRefDirect Link - Gurtler, M., U. Methner, H. Kobilke and K. Fehlhaber, 2004. Effect of orally administered egg yolk antibodies on Salmonella enteritidis contamination of hen's eggs. J. Vet. Med. B Infect. Dis. Vet. Public Health, 51: 129-134.
CrossRefDirect Link - Graczyk, T.K., M.R. Cranfield, M.L. Shaw and L.E. Craig, 1994. Maternal antibodies against Plasmodium spp. in African black-footed penguin (Spheniscus demersus) chicks. J. Wildl. Dis., 30: 365-371.
CrossRefDirect Link - Graczyk, T.K., M.R. Cranfield and C.J. Shiff, 1993. ELISA method for detecting anti-Plasmodium relictum and anti-Plasmodium elongatum antibody in infected duckling sera using Plasmodium falciparum antigens. J. Parasitol., 79: 879-885.
CrossRefDirect Link - Palmer, J.L., T.F. McCutchan, F.H., Vargas, S.L. Deem, M. Cruz, D.A. Hartman and P.G. Parker, 2013. Seroprevalence of malarial antibodies in Galapagos penguins (Spheniscus mendiculus). J. Parasitol., 99: 770-776.
CrossRefDirect Link - Van den Akker, R., H. Kooy, H.P. van der Meyden and B.H. Lumey, 1990. Sir: Recovery of HIV antibodies in eluates from plasma and erythrocytes dried on filter paper and stored under various conditions. Aids, 4: 90-91.
PubMedDirect Link - Arya, S.C., 1993. Stability of human immunodeficiency virus type 1 antibodies in whole blood-impregnated filter papers under various tropical conditions. J. Clin. Microbiol., 31: 765-766.
PubMedDirect Link - Waldenstrom, J., S. Bensch, D. Hasselquist and O. Ostman, 2004. A new nested polymerase chain reaction method very efficient in detecting Plasmodium and Haemoproteus infections from avian blood. J. Parasitol., 90: 191-194.
CrossRefPubMedDirect Link - Atkinson, C.T., R.J. Dusek and J.K. Lease, 2001. Serological responses and immunity to superinfection with avian malaria in experimentally-infected Hawaii Amakihi. J. Wildl. Dis., 37: 20-27.
CrossRefDirect Link - Ha, H.J., M. Banda, M.R. Alley, L. Howe and B.D. Gartrell, 2012. The seroprevalence of avipoxvirus and its association with avian malaria (Plasmodium spp.) infection in introduced passerine birds in the Southern regions of the North Island of New Zealand. Avian Dis., 57: 109-115.
CrossRefDirect Link - Jarvi, S.I., J.J. Schultz and C.T. Atkinson, 2002. PCR diagnostics underestimate the prevalence of avian malaria (Plasmodium relictum) in experimentally-infected passerines. J. Parasitol., 88: 153-158.
CrossRefDirect Link - Valkiunas, G., 2011. Haemosporidian vector research: Marriage of molecular and microscopical approaches is essential. Mol. Ecol., 20: 3084-3086.
CrossRefDirect Link - Sehgal, R.N.M., G. Valkiunas, T.A. Iezhova and T.B. Smith, 2006. Blood parasites of chickens in Uganda and Cameroon with molecular descriptions of Leucocytozoon schoutedeni and Trypanosoma gallinarum. J. Parasitol., 92: 1336-1343.
CrossRefDirect Link - Ochsenbein, A.F. and R.M. Zinkernagel, 2000. Natural antibodies and complement link innate and acquired immunity. Immunol. Today, 21: 624-630.
CrossRefDirect Link - Mendes, L., T. Piersma, D. Hasselquist, K.D. Matson and R.E. Ricklefs, 2006. Variation in the innate and acquired arms of the immune system among five shorebird species. J. Exp. Biol., 209: 284-291.
CrossRefDirect Link - Melo, A.R., E.B. Lasunskaia, C.M.C. De Almeida, A. Schriefer, T.L. Kipnis and W.D. da Silva, 2005. Expression of the virulence factor, BfpA, by enteropathogenic Escherichia coli is essential for apoptosis signalling but not for NF‐κB activation in host cells. Scand. J. Immunol., 61: 511-519.
CrossRefDirect Link