
I.J. Reddy
Division of Animal Physiology, National Institute of Animal Nutrition and Physiology, Adugodi, Hosur Road, 560 030 Bangalore, India
Ashish Mishra
Division of Animal Physiology, National Institute of Animal Nutrition and Physiology, Adugodi, Hosur Road, 560 030 Bangalore, India
S. Mondal
Division of Animal Physiology, National Institute of Animal Nutrition and Physiology, Adugodi, Hosur Road, 560 030 Bangalore, India
LiveDNA: 91.1478
Vaibhav Awachat
Division of Animal Physiology, National Institute of Animal Nutrition and Physiology, Adugodi, Hosur Road, 560 030 Bangalore, India
G. Ravi Kiran
Division of Animal Physiology, National Institute of Animal Nutrition and Physiology, Adugodi, Hosur Road, 560 030 Bangalore, India
Current Research in Poultry Science
Year: 2016 | Volume: 6 | Issue: 1 | Page No.: 13-17







ABSTRACT
Objective: The aim of this study was to investigate the effects of different wavelengths of light on egg production and associated hormonal changes during the later stages of productive period in white leghorn hens. Methodology: The effect of light colour on egg laying and endocrine profiles during 82-92 weeks of egg production was examined. Hens were exposed to normal (450 nm, n = 12, control) and far red (675 nm, n = 12, treated) sources of light. Results: Light intensity was equalized at photon output. Total egg production per hen was significantly influenced by the colour of light with the most eggs procured in red light and the fewest in normal light. Hens were more sensitive to light colour with elevated levels of estradiol (E2β), progesterone (P4), GnRH mRNA, Luteinizing hormone (LH) and egg production. Hens in normal light produced significantly fewer eggs than those under red spectrum light. The GnRH mRNA level was higher (p<0.01) in treated birds with high concentrations of LH, E2β and P4 in plasma. More egg lay, less pause days in treated group may be the result of high GnRH associated with high concentrations of LH, E2β and P4 concentration required for completion of egg formation and egg lay. Conclusion: It can be concluded that the red spectrum of light at 675 nm of wavelength of light may influence GnRH, gonadotrophic, gonadal hormones and increases egg production in ageing hens.
PDF Abstract XML References Citation
Received: May 26, 2016;
Accepted: June 05, 2016;
Published: June 15, 2016
How to cite this article
I.J. Reddy, Ashish Mishra, S. Mondal, Vaibhav Awachat and G. Ravi Kiran, 2016. Endocrine Changes in Hens (Gallus gallus domesticus) Exposed to Red Spectrum of Light. Current Research in Poultry Science, 6: 13-17.
URL: https://scialert.net/abstract/?doi=crpsaj.2016.13.17
URL: https://scialert.net/abstract/?doi=crpsaj.2016.13.17
INTRODUCTION
The photo sexual responses and subsequent alterations in reproductive activities in birds have been shown to be mediated by retinal and extra-retinal (brain) photoreceptors (EL-Halawani, 2007). Birds subjected to a gonad stimulating photoperiods, long wave radiation (630-750 nm) penetrates the tissue and directly acts on hypothalamic extra-retinal photoreceptors to stimulate reproductive function (Benoit, 1964; Benoit and Assenmacher, 1966; Menaker and Underwood, 1976; EL-Halawani, 2007). In contrast to the stimulatory effect of long wave radiation on reproductive activity, activation of retinal photoreceptors by visible radiation appears to be less effective to reproduction (Homma et al., 1972; Siopes and Wilson, 1980; EL-Halawani, 2007). The response to visible radiation is surmised to be mediated by the green-yellow bands of the light spectrum (545-575 nm), where the avian retina is maximally sensitive (Prescott and Wathes, 1999; Lewis and Morris, 2000; EL-Halawani, 2007). Taken together, the aforementioned findings lead to the following hypothesis: Photostimulation with radiation in the red band of the spectrum (630-750 nm) under minimum light intensity (brightness; minimum radiation in the green-yellow bands of the spectrum) will provide the greatest photostimulation effect and enhances egg production (EL-Halawani, 2007). This study aims at optimizing existing procedures and developing new tools for reproductive technologies of the future. To optimize existing management procedures, the effects of different light wavelength and photoperiod on egg production using ageing chickens was evaluated.
MATERIALS AND METHODS
At 82 weeks of age, 24 white leg horn birds were housed in individual cages (one bird per cage) under two-tier battery system. Birds were divided into two groups consisting of 12 birds in each. All birds were fed on the same layer ration (16% CP and 11.72 MJ ME kg–1) as per the standard recommendations (Ranjhan, 1993) and were provided 16 h light and 8 h dark. Clean water was made available round the clock throughout the experimental period. From 82-92 weeks of age the birds in treatment group were exposed to 650 nm of wavelengths of light (red spectrum of light-treatment group). Controls were exposed to normal spectrum of light with white bulbs at 450 nm of wavelength of light and treated group were exposed to red spectrum of light at 650 nm of wavelengths of light with red bulbs. Bulbs were fixed in the experimental unit so as to provide uniform intensity of light to all the birds within the group without any variation as for as source of light with an intensity of 0.1 W m–2 at bird-head level. Watts per square meter units were used because extra retinal photoreceptors located in bird’s head detect energy level penetrating through the skull. The control group was (housed in separate partisan) provided with normal light bulbs 0.1 W m–2 at bird-head level. The lighting schedule was 16 h light and 8 h darkness. Daily egg production was recorded for each hen at the same time for 77 days. Mean weekly egg production was recorded from both the groups. Total number of pause days during 11 weeks period (77 days) was recorded. Ovipositions were recorded between 9 am to 12 noon in both the groups. Ovipositions in the control group were irregular with more days between the sequences of egg lay compared to treated group. Blood samples were collected from each bird by superficial venipuncture of the brachial vein starting from 82 weeks of age onwards at weekly intervals and continued until the end of experimental period at 92 weeks of age. Plasma was separated and stored at -20°C for hormone assay. Oviposition times were recorded, while collecting blood samples in both the groups. To determine the effect of red and normal spectrum of light on the distribution of Yellow Yolky Follicles (YYFs) (>8 mm) in the ovary, birds in both the groups were decapitated at the end of 92 weeks of age. Hypothalamic were dissected to include GnRH-I (Lewis and Morris, 2000) cell bodies. Hypothalamic and pituitary glands were weighed and snap frozen in liquid nitrogen and stored at -80°C until RNA extraction. The number of yellow yolky follicles >8 mm, follicles between 5 and 8 mm, residual ovary and oviduct weights were recorded. Hypothalamic cell bodies containing GnRH-I were cultured separately from both the groups in DMEM with phenol red, supplemented with 3.75% fetal calf serum and 65% horse serum, antibiotics: 100 μg mL–1 streptomycin and 100 μg mL–1 penicillin. The culture plates were placed in air tight containers and after equilibration with 95% O2 and 5% CO2 transferred to an incubator maintained at 37°C. Media changed for every 2 days and maintained for 6 days. Superficial fluid is removed and the cells which were attached are subjected to RNA extraction with microliter trizol (Invitrogen). The tissues were disrupted using homogenizer. The yield of RNA was quantified by measuring the optical density of a sample diluted to 1:50 at 260 and 280 nm and its quality was confirmed by running a sample out on a formaldehyde gel.
Reverse transcription of total RNA: A sample of (4 μL) of total RNA was reverse transcribed using a first strand synthesis kit as per the manufacturer’s instructions. Reverse transcribed samples were diluted to 40 μL in dH2O. The PCR primers in QC RT-PCR assays for GnRH mRNA were made from GenBank Accession No: X69491. The GnRH-I forward primer as TGGGTTTGTTGATGGTGTTGT and reverse primer as ATTTTCCAGCGGGAAGAGTTG. Chicken GnRH-I in both control and treated group were measured by quantitative (QC) RT-PCR assays. Oligonucleotide primers for the amplification of neuroendocrine gene GnRH were designed using the "primer" software package version 0.5 and published cDNA sequences. The PCR amplification of was carried out in a thermo-fast low profile 96 well plate on a programmable heating block. The PCR conditions were 30 cycles (94°C, 20 sec; 62°C, 20 sec; 72°C, 20 sec) for GnRH-I.
Analysis of hormones: Chicken cLH antisera and pure hormone were obtained from John. A. Proudman, USDA as a gift from USA. The intra and inter coefficient variation for cLH was 4.45 and 8.12%, respectively with sensitivity of the hormone 0.01 ng mL–1 per tube as per the method described previously (Sharp et al., 1987). The E2β and P4 samples were analysed with RIA kits obtained from ImmunoTech, France.
Statistical analysis: Measurements were given as Mean±SE. The significance of differences between means was analysed by F-test. The data on egg production, LH, E2β and P4 were subjected to correlation coefficient analysis to study the influence of the hormones on egg production. Differences were considered significant at a value of p<0.01. The statistical analysis were carried out following the standard method (Snedecor and Cochran, 1994). Log transformed data was performed for GnRH-I mRNA data by one way ANOVA analysis.
RESULTS
The plasma E2β level in birds of control group varied between 241.11±1.02 to 51.12±1.01 pg mL–1 during 82-92 weeks of age (Fig. 1). In treatment group plasma E2β increased from 236.14±1.33 to 269.19±1.20 pg mL–1 during 82-92 weeks of age. The P4 secretion in the two groups also followed a similar pattern as estradiol and is presented in Fig. 2. However, intermittent hormonal fluctuations were observed in both control and treated groups. Egg production in birds exposed to red spectrum of light was positively correlated with E2β (r = 071), P4 (r = 0.65) and LH (r = 0.54), whereas GnRH-I mRNA level was positively correlated with E2β (r = 0.63), P4 (r = 0.71) and LH (r = -0.59) and negatively correlated with pause days (r = -0.18). Plasma LH concentration in control group varied between 2.32±0.09 to 3.5±0.4 ng mL–1 during 82-92 weeks of age (Fig. 3). In treatment group, plasma LH increased from 2.15±0.11 to 5.01±0.02 ng mL–1 during 82-92 weeks of age.
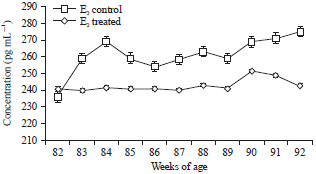 | |
| Fig. 1: | Plsma estradiol (pg mL–1) between control and treated birds |
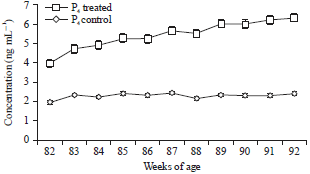 | |
| Fig. 2: | Plasma progesterone (pg mL–1) between control and treated birds |
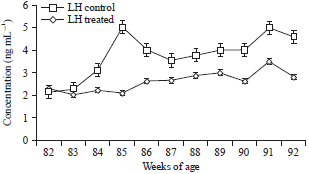 | |
| Fig. 3: | Mean LH concentration between control birds and treated birds |
The LH levels fluctuated between the two groups and treated birds showed significantly (p<0.01) higher levels over the controls. In treatment group, the increase in LH level was of greater magnitude due to the exposure of red spectrum of light (treatment) for eleven weeks (Fig. 3).
GnRH-I mRNA: There were differences in hypothalamic GnRH-I mRNAs expression between treated and control hens. The GnRH-I mRNA levels were significantly higher in birds exposed to red spectrum of light (16.44) over the control birds (8.31). The GnRH concentration was expressed in 10–14moles per hypothalamus (Fig. 4).
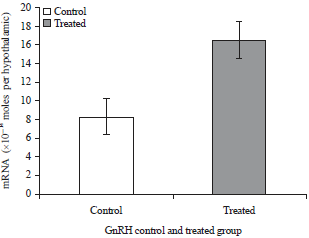 | |
| Fig. 4: | Hypothalamic GnRH-I mRNA in control birds and treated birds |
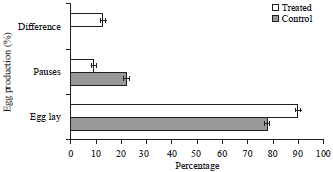 | |
| Fig. 5: | Percentage of egg production, percentage of pause days, difference in percentage of egg production between control and treated birds |
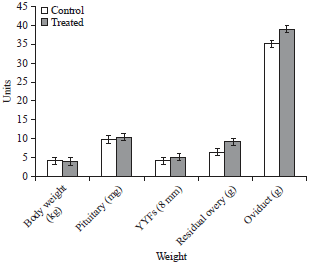 | |
| Fig. 6: | Effect of wavelengh of light on body weight, pituitary weight, residual ovary weight and oviduct weight and numbers of yellow yolk follicles in control and treated birds |
Influence of different spectrum of light on egg production parameters: The average egg production (bird per week) exposed to red spectrum of light (treatment group) from 82-92 weeks of age was significantly (p<0.01) higher than the controls (Fig. 5). It fluctuated between 89-90% and 75-77.92% in birds exposed to red spectrum of light and incandescent light, respectively. The difference in egg production per bird per week was significant between two groups from 82 weeks of age to 92 weeks of age. The percentages of pause days during the 11 weeks period (77 days) (Fig. 5) were 17.0±0.12 in the treatment group as against 7.00±0.41 in control group. Residual ovary is the one after removal of the hierarchal follicles F1-F5. It comprises of small yellow and white follicles but not stroma alone. Egg production in treatment group between 82 and 92 weeks of age, improved by 12.93% over control group (Fig. 6).
DISCUSSION
It is evident that hens photostimulated with light spectrum at the 675 nm wavelength showed more egg production as compared with those photostimulated with 450 nm wavelengths or full light spectrum from incandescent lamps. However, it is not known whether the amount of energy emitted by red light is exceeding or below the level that is ideal for optimum egg production. It is possible that excess energy penetrating the brain may have stimulatory effect on egg production and less energy may not be sufficient to maximize egg production. Accordingly, the effect of 675 and 450 nm of wavelength of light on egg production was tested. This may be explained by the fact that wavelengths of higher intensities possess higher power and are thus able to penetrate the skull, brain tissue more easily and stimulate receptors responsible for GnRH release in the hypothalamus and these receptors are suggested to be sensitive to light directly passing through the skull instead of perception of light by retina (Kuenzel, 1993; Bedecarrats et al., 2006, 2009; Lewis and Morris, 2000; Lewis et al., 2005). The GnRH intern stimulates gonadotrophic and gonadal hormones. It is well established that GnRH, LH, FSH and steroid hormones act simultaneously on follicular development, ovulation, egg formation and egg lay in hen. Thus the red spectrum of light in layer hen house changed the reproductive system of hen from low functional sate to a high functional and increased egg production by 12.93% in treated group from 82-92 weeks of age (Fig. 5). This may be attributed to high GnRH in the treated group relative to control group (Fig. 4). Further, normal wavelengths of light may not be able to penetrate the skull (Morgan et al., 1995; Bedecarrats et al., 2006, 2009) thus, light at this intensity may be unable or less likely to excite receptors to release GnRH. Decreased GnRH, LH, E2β and P4 levels in control birds under normal wavelengths of light also support this hypothesis. The decreased GnRH in the control group failed to increase the circulating levels of LH, E2β and P4, whereas red light was the most effective. Higher levels of estradiol during the initiation of egg laying have been correlated with the activity of small follicles (Bedecarrats et al., 2009). This effect of light wavelength on ovarian activity was independent of a fully functional retina of the eye and is thus probably mediated via extra-retinal photoreceptors, most likely hypothalamic. The effect of red spectrum of light on ovarian morphology was stronger than the normal light. The results shows that reduced egg production in birds exposed to normal spectrum of light is associated with a reduction in YYFs and reduced plasma LH (Fig. 6). The reduction in numbers of YYFs and plasma LH agrees with a previous study in broiler breeders (Sharp et al., 1990) of hens. It is therefore likely that reduced ovarian function in control hens is caused by a reduction in gonadotropin secretion (Sharp, 2005). Changes in pituitary glands may be attributed to longer wavelengths are transmitted through neural tissue more readily than shorter wavelengths (Lewis et al., 2005). The feed intake was recorded only during the 82-92 weeks period of the study and no difference was found in the intake between the groups within the short period of time. The body weights were taken only at slaughter and whatever difference observed may be due to the changes before the initiation of study prior to 92 weeks age of the birds. Egg production of control hens that were photostimulated with incandescent lamps peaked by the 5th week of photostimulation. The highest peak production (90.90%) was observed at the highest energy level 675 nm. However, the overall egg production during the 77 days period was lowest (77.92%) in hens photostimulated with incandescent lamps.
CONCLUSION
Results of this study show that red light is necessary to adequately initiate the activation of the reproductive axis, through GnRH release to increase ovarian activity to increase total number of eggs in aged birds.
ACKNOWLEDGMENTS
The authors are thankful to Director, NIANP, Bangalore for providing necessary facilities to carryout the work. We are thankful to Dr. John. A. Proudman, USDA, USA for providing chicken LH hormone and antisera as a gift.
REFERENCES
- Bedecarrats, G.Y., H. McFarlane, S.R. Maddineni and R. Ramachandran, 2009. Gonadotropin-inhibitory hormone receptor signaling and its impact on reproduction in chickens. Gen. Comp. Endocrinol., 163: 7-11.
CrossRefDirect Link - Bedecarrats, G.Y., M. Shimizu and D. Guemene, 2006. Gonadotropin releasing hormones and their receptors in avian species. J. Poult. Sci., 43: 199-214.
CrossRefDirect Link - Benoit, J., 1964. The role of the eye and of the hypothalamus in the photostimulation of gonads in the duck. Ann. New York Acad. Sci., 117: 204-215.
CrossRefDirect Link - Benoit, J. and I. Assenmacher, 1966. [Research on the photosensitivity of superficial and deep neural receptors in gonad stimulation by visible radiation in the immature Pekin duck]. Comptes Rendus Hebdomadaires Seances L'Academie Sciences Serie D: Sciences Naturelles, 262: 2750-2752, (In French).
PubMedDirect Link - Homma, K., W.O. Wilson and T.D. Siopes, 1972. Eyes have a role in photoperiodic control of sexual activity of Coturnix. Science, 178: 421-423.
CrossRefDirect Link - Kuenzel, W.J., 1993. The search for deep encephalic photoreceptors within the avian brain, using gonadal development as a primary indicator. Poult. Sci., 72: 959-967.
CrossRefDirect Link - Lewis, P.D. and T.R. Morris, 2000. Poultry and coloured light. World's Poult. Sci. J., 56: 189-207.
CrossRefDirect Link - Lewis, P.D., M. Ciacciariello, N.A. Ciccone, P.J. Sharp and R.M. Gous, 2005. Lighting regimens and plasma LH and FSH in broiler breeders. Br. Poult. Sci., 46: 349-353.
CrossRefDirect Link - Menaker, M. and H. Underwood, 1976. Extraretinal photoreception in birds. Photochem. Photobiol., 23: 299-306.
CrossRefDirect Link - Prescott, N.B. and C.M. Wathes, 1999. Reflective properties of domestic fowl (Gallus gallus domesticus), the fabric of their housing and the characteristics of the light environment in environmentally controlled poultry houses. Br. Poult. Sci., 40: 185-193.
CrossRefDirect Link - Sharp, P.J., R.T. Talbot, G.M. Main, I.C. Dunn, H.M. Fraser and N.S. Huskisson, 1990. Physiological roles of chicken LHRH-I and -II in the control of gonadotrophin release in the domestic chicken. J. Endocrinol., 124: 291-299.
CrossRefDirect Link - Sharp, P.J., 2005. Photoperiodic regulation of seasonal breeding in birds. Ann. New York Acad. Sci., 1040: 189-199.
CrossRefDirect Link - Siopes, T.D. and W.O. Wilson, 1980. Participation of the eyes in the photostimulation of chickens. Poult. Sci., 59: 1122-1125.
CrossRefDirect Link - Sharp, P.J., I.C. Dunn and R.T. Talbot, 1987. Sex differences in the LH responses to chicken LHRH-I and -II in the domestic fowl. J. Endocrinol., 115: 323-331.
CrossRefDirect Link - Morgan, I.G., M.K. Boelen and P. Miethke, 1995. Parallel suppression of retinal and pineal melatonin synthesis by retinally mediated light. Neuroreport, 6: 1530-1532.
PubMed