
Qinghua Liu
College of Animal Science, Fujian Agriculture and Forestry University, Fuzhou 350002, China
Yan Wang
Key Laboratory of Biotic Environment and Ecological Safety, College of Life Sciences, Anhui Normal University, Wuhu 241000, Anhui Province, China
Yafei Cai
Key Laboratory of Biotic Environment and Ecological Safety, College of Life Sciences, Anhui Normal University, Wuhu 241000, Anhui Province, China
Huiling Liu
Key Laboratory of Biotic Environment and Ecological Safety, College of Life Sciences, Anhui Normal University, Wuhu 241000, Anhui Province, China
Jie Chen
Key Laboratory of Biotic Environment and Ecological Safety, College of Life Sciences, Anhui Normal University, Wuhu 241000, Anhui Province, China
Ping Li
Key Laboratory of Biotic Environment and Ecological Safety, College of Life Sciences, Anhui Normal University, Wuhu 241000, Anhui Province, China
Jienan Cheng
Key Laboratory of Biotic Environment and Ecological Safety, College of Life Sciences, Anhui Normal University, Wuhu 241000, Anhui Province, China
Honglin Liu
College of Animal Science and Technology, Nanjing Agricultural University, Nanjing 210095, China
Current Research in Poultry Science
Year: 2011 | Volume: 1 | Issue: 2 | Page No.: 37-53







ABSTRACT
The process of T Lymphocyte involvement in the development of avian bursa of fabricius remains unclear. In order to clarify the feature of this process, the fetal goose bursas of fabricius from E17 (17-day-embryo) to 18 d (18-day after hatching) were adopted to study the morphological indices, 4 and CD8 co-localization in bursas of fabricius were determined by fluorescence microscopy. Cell apoptosis rate were analyzed the apoptosis with software Image Pro Plus 6.0. Like an appendix, the goose bursa of fabricius was located on the back of cloaca. The lymphatic follicle increased during the prenatal stage and became degenerated after hatching. For the first time, CD4 and CD8 co-expression (double positive) was found in the connective tissue and cortex and co-expression level increased from E17 to E26, then decreased from E28 to 7d. It was significantly different (p<0.01) among E17, E24, E26, E28, 18 and 3d. The apoptosis rate decreased from E17 to 3d, then dramatically increased after hatching and was extremely different (p<0.01) among E17, E24, E26, E28, 18 and 3d. The coordination between co-expression level and apoptosis rate in bursa of fabricius is significant. It is deduced that T lymphocyte selection in bursa of fabricius sharing the same feature as the T lymphocyte selection in thymus is indispensible for the development of goose bursa of fabricius.
PDF Abstract XML References Citation
Received: March 23, 2011;
Accepted: June 25, 2011;
Published: August 17, 2011
How to cite this article
Qinghua Liu, Yan Wang, Yafei Cai, Huiling Liu, Jie Chen, Ping Li, Jienan Cheng and Honglin Liu, 2011. T Lymphocyte Selection is Indispensible for the Development of Goose Bursa of Fabricius. Current Research in Poultry Science, 1: 37-53.
URL: https://scialert.net/abstract/?doi=crpsaj.2011.37.53
URL: https://scialert.net/abstract/?doi=crpsaj.2011.37.53
INTRODUCTION
Bursa of fabricius is one of central lymphoid organs in birds (Rodriguez-Mendez et al., 2010). In bursal development, it undergoes striking changes including the rapid development from the late embryogenesis to post-hatching and degeneration after sexual maturity (Luna et al., 2008). It contains many lymphatic follicles and is known as the primary lymphoid organ which dominates proliferation and diversification of B cell (Ackerman and Knouff, 1959; Cooper et al., 1969; Olah and Glick, 1978; Reynaud et al., 1991). It plays a central role in avian B-cell development and participates in the B-cell proliferation and immunoglobulin V gene diversification (Brown et al., 2004).
Previous studies revealed that there was a Diffusely Infiltrated Area (DIA) of lymphoid cells just dorsal to the chicken bursal duct opening (Odend’hal and Breazile, 1980; Dolfi et al., 1988) and DIA was previously described as a T-dependent bursal area (Cortes et al., 1995). DIA cells are almost T lymphocytes including CD4+ cells, CD8+ cells and TCR lymphocytes (Cortes et al., 1995) can activate B-cell (Larosa and Orange, 2008).
Avian T-cell populations can be divided into different subsets based on their different cell-surficial antigens such as CD4, CD8 and TCR (Erf et al., 1998). The CD4 and CD8 receptors are made of single polypeptides. Most CD4+ cells are helper T-cells responding to exogenous antigen in association with major histocompatibility complex Class II (MHC II) molecules; while CD8+ cells serve as cytotoxic T-cells responding to endogenous antigen in association with MHC I molecules. T lymphocyte subpopulations plays an important role in the innate immune response against intracellular pathogens (Liu et al., 2011).
The CD4 and CD8 expressions can be detected in the thymus and spleen of 2-7 week-old broilers (Erf et al., 1998). At early embyo phase, the thymocytes express neither CD4 nor CD8. Then CD4 and CD8 were co-expressed in immature thymocyte at middle embyo phase. The double-positive cells react with self antigens and induce apoptosis in the thymus (negative selection), thus negative selection results in the occurance of CD4+CD8- or CD4-CD8+ T-cells (Oguma et al., 2009). CD4 and CD8 co-expression is considered as a marker of immature cell. CD4+ or CD8+ single-positive T-cells are mature cells which enter secondary immune organs and travel in the circulatory lymphatic systems (Erf et al., 1998).
All in all, whether T-dependent bursal area participates in B cell proliferation in bursa of fabricius remains unclear (Yamamoto et al., 1996), especially in goose. Up to now, little is known about T or B lymphocyte selection in lymphoid organs. Unveiling the role of T lymphocyte selection in bursa of fabricius will contribute to understand their roles in immune response in goose. Therefore, we marked T cells with CD4 and CD8 expression in bursa of fabricius during bursal development. The objective of study was demonstrated that whether T lymphocyte selection in bursa of fabricius is similar to T cell development in the thymus and is indispensible for bursal development or not.
MATERIALS AND METHODS
Materials and reagents: Goose embryos [17-day (E17), 24-day (E24), 26-day (E26) and 28-day (E28) in appendix embryos] and young geese [3-day (3d), 7-day (7d) and 18-day (18d)](supplement material Fig. 1A-D in appendix) were collected from Sanyuan Breeding Limited-liability Company (Wuwei, Anhui Province, China).
Polyclonal rabbit anti-rat/mouse CD4 antibody and goat anti-rabbit IgG-FITC were bought from Beijing Biosynthesis Biotechnology Co Ltd (Beijing, China). Monoclonal mouse anti-human CD8 antibody, goat anti-mouse IgG-TRITC (tetraethyl rhodamine isothiocyanate), APES (aminopropyl-triethoxysilane), BSA (bovine serum albumin) and PBS (phosphate buffered saline) were bought from Wuhan Boster Biology Technology Ltd. (Wuhan, China).
Histology development of goose bursa of fabricius: The bursa of fabricius collecting from E17 to 18d samples (n = 5 at each developmental stage) were fixed in paraformaldehyde for 24 h at room temperature, dehydrated in ethanol and embedded in paraffin wax. Tissue sections (5 μm thick) were cut with a microtome and mounted onto APES-coated glass slides. Serial sections were cleared in dimethylbenzene, rehydrated in a graded series of ethanol and then incubated for Hematoxylin-Eosin (HE) staining.
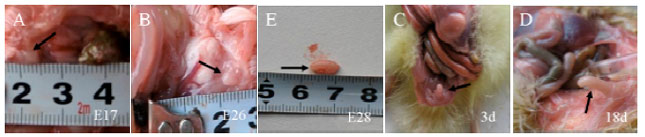 | |
| Fig. 1 (A-D): | The anatomical photos of goose bursa of fabricius at different stages |
Immunofluorescence analysis, CD4 CD8 expression and T lymphocyte localization: Tissue sections were incubated with polyclonal rabbit anti-rat/mouse CD4 antibody (at a dilution of 1:100) and mouse monoclonal anti-CD8 antibody (at a dilution of 1:100) in 1% PBS at 4°C overnight. After being washed (3x10 min) with PBS, sections were incubated at 37°C 1h with the secondary antibodies: Goat anti-rabbit IgG-FITC (1:32) and goat anti-mouse IgG-TRITC (1:32). After being rinsed (3x10 min) with PBS, sections were sealed with glycerin. CD4 and CD8 co-localization were observed via fluorescence microscopy with lasers at excitation wavelengths of 488 nm (FITC), 514 nm (TRITC), 350 nm (Hoeschst33342), respectively. OLYMPUS BX61 fluorescence microscopy observation analysis were performed to determine CD4 and CD8 co-localization in bursa of fabricius.
Apoptosis analysis of bursal development: Sections were also sealed with 1% BSA, then stained with 1% Hoeschst33342 in PBS at 37°C for 15 min, rinsed (3x10 min) with PBS and coated with glycerin, analyzed the apoptosis rate with software Image Pro Plus 6.0.
Statistical analysis: The data were expressed as Mean±SE and analyzed using one-way ANOVA. Data were transformed to ensure homogeneity of variance. LSD’s multiple comparisons were applied to identify differences with homogeneity of variance. Tamhane’s multiple comparisons were adopted to check non-homogeneity of variance. Probability values were considered to be significant at the 5% between groups. Statistical analysis was performed using SPSS13.0 package version by a two-tailed test.
RESULTS
Anatomical profile of goose bursa of fabricius at different stages: The goose bursa of fabricius was connecting with cloacal back wall via the bursal duct (black arrows, Fig. 1A-D). It increased in length and diameter from E17 (0.4 cm) to 18d (1.6 cm) and reached maximum at 18d (1.6 cm). The bursa of fabricius was like a sac, linking up with cloaca via ostioles. The bursal cystic wall was similar to digestive canal, consisted of mucous membrane, the submucosa and muscularis and outer membrane.
The bursal histologic changes at different stages: The morphology of goose embryonic bursa of fabricius:
E17:The mucosal epithelium of bursa of fabricius was composed by stratified epithelium; the submucosa was consisted of loose connective tissue. The outer membrane was the serosa which comprised mesothelium and connective tissue (Fig. 2A)
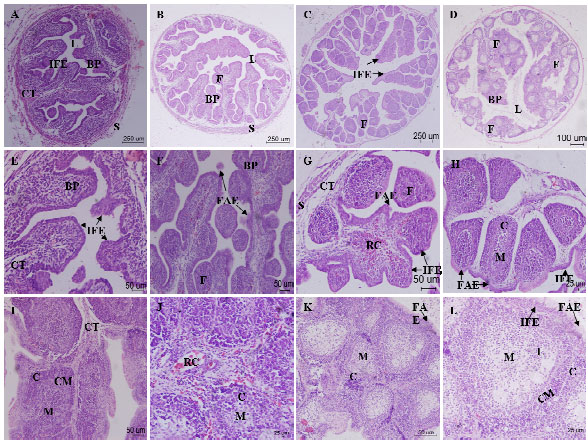 | |
| Fig. 2 (A-L): | Histological changes of bursal follicle and epithelium at different stages Goose bursal sections were stained with Hematoxylin-eosin (HE). A-D: Structure of goose bursa of fabricius at E17, E24, E28, 18d, Magnification in A-D: x40, bars 250 μm. E-K: Structure at E17, E24, E26, E28, 3d, 7d, 18d; Magnification in (E, F, G, I, K): x200, bars 50 μm; (H, J): x400, bars 25 μm. L: Structure of lymphoid follicles, Magnification in: x400, bars 25 μm. BP: Bursal plicae; F: Bursal follicles; L: Lumen; CT: Connective tissue; M: Medulla; C: Cortex; CM: Corticomedullary; IFE: Interfollicular epithelium; FAE: Follicle-associated epithelium; RC: Red cell |
E24:The bursa of fabricius became thicker. There were some elongated mucosal folds on the mucosal surface. The number of nodular lymphocyte cell masses gradually increased. The loose connective tissue in the submucosa was forming large trabecular meshwork. The muscularis layer was hypoplasia. A few follicle-associated epithelium were located on the mucosal surface and played a role in phagocytosis (Fig. 2B)
E26:Bursal mucosal epithelium became thicker increasingly and formed many leafy mucosal folds before lymphoid follicle emergence. The lymphoid follicles included cortex and medulla, whereas their boundary was not evident. There are several small lymphocytes, B lymphocytes, T lymphocytes and many macrophages existing in the follicles (Fig. 2C)
E28:The number and volume of lymphoid follicles increased significantly. The cell density in the cortex was much more than that in the medulla. Some heterophilic leukocytes and lymphocytes-like cells dispersed in the cortex. The number of follicular absorptive epithelium in the lymphoid follicle increased and follicular secretory epithelium appeared (Fig. 2D)
The morphology of goose bursa of fabricius after hatching:
3d:The bursa of fabricius developed well. The shape of most lymphoid follicles was irregular. The cortex and medulla can be easily distinguished in the submucosa. The number of follicular absorptive epithelium and follicular secretory epithelium increased (Fig. 2E)
7d:The shape of lymphoid follicles became oval. A large number of the reticular cell and reticular fiber were present in lymphoid follicles; lots of heterophilic leukocytes around follicles; many blood cell in the trabecular meshwork (Fig. 2F)
18d:The lymphoid follicles degenerated gradually. The boundary between cortex and medulla was still clear but there were only few lymphocytes in the medulla. Vacuolated structure was relatively obvious (Fig. 2G)
The comparison of the follicular number at different stages: There were approximately 3 bursal follicles at E17, 10 at E24, 17 at E26, 100 at E28, 128 at 3d, 120 at 7d and 78 at 18d (Table 1). It was clear that the volume and number of the lymphatic follicle increased from E17 to 3d and decreased after 3d (Fig. 3). The vacuolated structure was seen in lymphoid follicle after hatching.
The CD4 and CD8 expression in goose bursa of fabricius
The T lymphocyte distribution and CD4 CD8 co-expression during bursal development: The single positive CD4 (CD4+) were mainly expressed in the connective tissue around the mucosal folds and cortex in the lymphoid follicles. The single positive CD8 (CD8+) were also expressed in the connective tissue adjacent to the serosa and cortex in the follicles.
The CD4 and CD8 co-expression was predominantly in the connective tissue and rarely in the follicle at E17, also widely discovered in the connective tissue near the serosa at E24. The co-expression level increased and reached the peak in the connective tissue at E26. Although the co-expression in connective tissue gradually reduced at E28, it increased in the cortex and corticomedullary of follicles at 3d. Co-expression was more evident in the cortex at 7d while it is almost absent in the connective tissue. Then it gradually increased in the cortex and connective tissue near the junction of follicle at 18d. It is indicated that CD4 and CD8 co-expression were simultaneously distributed but more in the connective tissue and cortex during development (Fig. 4).
In summary, the co-expression clearly rised from E17 to E26, then reduced from E28 to 7d and increased at 18d. It was almost present in bursal plicae, follicle-associated epithelium, follicular secretory epithelium and lumen which were described as T-independent bursal areas.
Statistical analysis of double positive area percentage: The statistical results of CD4+CD8+ expression (double positive) in goose bursal areas during different periods were analyzed and shown in Table 2. The double positive area percentage relationship between different periods was analysed using software SSPS13.0. The results showed that it were significant differences between group (F (6,63) = 264.212, p<0.01 (p = 0.000)) using one-way ANOVA.
| Table 1: | The statistics of the bursal follicular number during different periods |
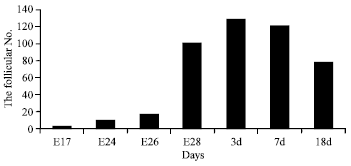 | |
| Fig. 3: | The comparison on the lymphoid follicular number during different periods |
| Table 2: | Statistical analysis showing double positive area percentage results in the goose bursa of fabricius during different periods |
| Data are expressed as Mean±SE. **Indicate that difference is extremely notable compared with 3d after hatching (p<0.01) | |
| Table 3: | The comparative analysis on goose bursal double positive area percentage during different periods |
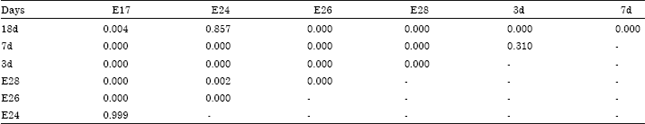 | |
| The mean difference is significant at the 0.05 level | |
It come to conclusion that it did not have the homogeneity of variance (p<0.05 (p = 0.000)) through test of homogeneity of variance (supplemental Table S1, S2, S3 in appendix). The multiple comparative analysis on goose bursal double positive area percentage during different periods was performed difference using Tamhane with non-homogeneity of variance (Table 3). The lowest was 0.000 and the highest was 0.999. The stem-and-leaf diagram showed that had significant differences during different periods (Fig. 5).
The results showed that: significant difference (p<0.01) were observed among E17 (p = 0.000), E24 (p = 0.000), E26 (p = 0.000), E28 (p = 0.000), 18d (p = 0.000) and 3d.
The apoptosis analysis on goose bursa of fabricius
Hoeschst33342 stained nuclei analysis: The small blue dot was apoptotic body of nuclei (the white arrows in Fig. 6h). At E17, nuclei were sparse in connective tissue and serosa, while they are intensive in the interfollicular epithelium and large numbers of apoptotic cells were seen in bursal follicle. At E24, nuclei arranged neatly and closely in the interfollicular epithelium and bursal follicle whereas loosely in the connective tissue connecting with serosa. At E26, E28 and 3d, the nuclei were compacted to each other but sparse in connective tissue and aligned closely in the cortex, while they scattered in the medulla at 7d and 18d.
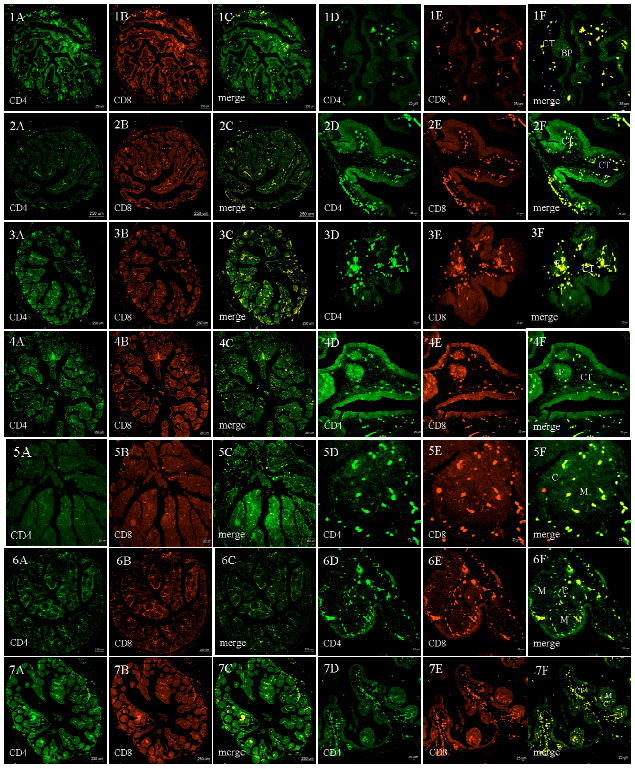 | |
| Fig. 4: | CD4 and CD8 immunoreactivity in goose bursa of Fabricius during different periods 1-7 represents E17, E24, E26, E28, 3d, 7d, 18d, respectively. A, D: CD4 expression (green, 1st Ab: polyclonal rabbit anti-rat/mouse CD4 IgG 1:100; 2nd Ab: goat antirabbit IgG-FITC, 1:32); B,E: CD8 expression (red, 1st Ab: mouse monoclonal anti- CD8 IgG 1:100; 2nd Ab: goat anti-mouse IgG-TRITC 1:32) in bursal follicles. C, F is shown in yellow after merging images. Magnification in A-C: 40x, bars 250 µm; in (D-F): 400x, bars 25 µm. BP: Bursal plicae; F: Bursal follicles; CT: Connective tissue; M: Medulla; C: Cortex |
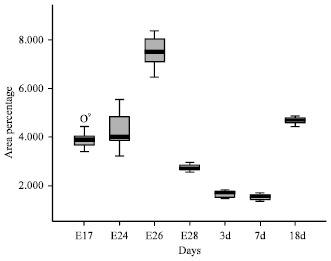 | |
| Fig. 5: | The stem-and-leaf diagram about double positive area percentage during bursal development. Rectangular box is the subject of the diagram, it shows the value of 25-75%. The thick black lines represent average value. o represents outliers; It shows 1 outliers in 70 numerical values |
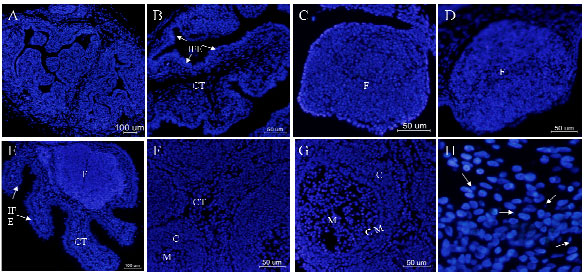 | |
| Fig. 6 (A-H): | Hoeschst33342 stained nuclei in goose bursa of fabricius during different periods A-G: separately represents E17, E24, E26, E28, 3d, 7d, 18d. Magnification in (A, E): x100, bars 100 μm; (B, C, D, F, G): x200, bars 50 μm. H: showing the apoptotic bodies of goose bursal nuclei, the white arrows pointed to apoptotic bodies. F: Bursal follicles; CT: Connective tissue; M: Medulla; C: Cortex; IFE: Interfollicular epithelium; CM: Corticomedullary |
Statistical analysis on apoptosis rate: Statistical analysis were carried on goose bursal apoptosis rate at different periods (Table 4). The stem-and-leaf diagram showed that there were significant differences among 3d and E17, E24, E28, 18d (Fig. 7).
There were significant differences between groups (F (6,63) = 5.811, p<0.01 (p = 0.000)) using one-way ANOVA and it had the homogeneity of variance (p>0.05 (p = 0.068)) through test of homogeneity of variance (supplemental Table S4, S5, S6 in appendix).
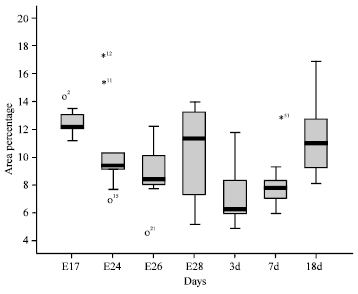 | |
| Fig. 7: | The stem-and-leaf diagram on apoptosis rate at different periods. Rectangular box is the subject of the diagram, it shows the value of 25-75%. The thick black lines represent average value. o represents outliers; * represents extreme value. It shows 3 outliers and 3 extreme values in 70 numerical values |
| Table 4: | The apoptosis rate in goose bursa of fabricius at different periods |
| Data are expressed as Mean±SE. **Indicate that difference is extremely notable compared with 3d after hatching (p<0.01) | |
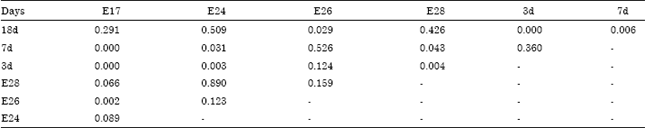
| Table 5: | The multiple comparison on the apoptosis rate the difference during different periods |
 | |
| The mean difference is significant at the 0.05 level | |
The multiple comparison on goose bursal apoptosis rates among different periods was performed difference using LSD with homogeneity of variance (Table 5). It was testified that extremely significant difference (p<0.01) were observed among E17 (p = 0.000), E24 (p = 0.003), E28 (p = 0.004), 18d (p = 0.000) and 3d.
The correlation analysis on co-expression area percentage and apoptosis: The statistical analysis between double positive area percentage and apoptosis rate also showed that there were significant differences among 3d and E17, E24, E28, 18d. Spearman’s rho was adopted to analyze the correlation between double positive area percentage and apoptosis rate. The results showed that: their correlation coefficient is 0.242, p = 0.043 (p<0.05). It is indicated that the correlation had significant difference between them (Table 6). It is suggested that there is cooperative relationship between them in bursal development.
| Table 6: | The correlations analysis between double positive area percentage and apoptosis rate |
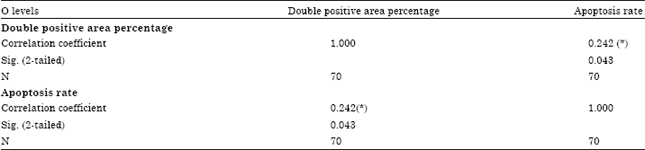 | |
| *Correlation is significant at the 0.05 level (2-tailed) | |
DISCUSSION
The bursa of fabricius is responsible for B cell maturation and antigen specific IgM-IgG switch (Nagy and Olah, 2009; Nagy and Olah, 2010; Ricci et al., 1996) or peptides (Garcia-Espinosa et al., 2008). The goose bursal development in this study was similar to chicken bursa of fabricius (Glick et al., 1956; Nagy and Olah, 2009). Except for different incubation period, the size and developing pattern were consistent to those of the chicken during embryogenesis and early weeks after hatching (Luna et al., 2005). It is reported that the morphology of human intestinal Peyer patches, rabbit appendices, Sheep Ileal Peyer Patches (SIPP) and the avian bursa of fabricius are similar (Dasso et al., 2000). In mammals and birds, B-cell differentiation starts from VDJ recombination in central lymphoid organs (Butler, 1997; Griebel and Hein, 1996). Historically, there is Germinal Center (GC) structure in the rabbit appendix, follicular structure in the avian bursa of fabricius and SIPP (Hodges, 1974; Reynolds and Morris, 1983). It was also be revealed that the avian bursal follicle can be divided into cortex and medulla. In present study, the vacuolated structure appeared after hatching, however, SIPP not were found. It was shown that there were structural differences in avian bursa of fabricius, Peyer patches and appendix in mammals. It is known that continuing T-cell can activated B cells and initiate the GC reaction, or differentiate into short-lived plasma cells (Larosa and Orange, 2008). Our viewpoints are in agreement with previous reports (Odend’hal and Breazile, 1980; Dolfi et al., 1988) that goose had T-dependent area in the bursa of fabricius. We come to conclusion that connective tissue and cortex were T-dependent bursal areas in the goose. This research provides theoretical basis and evidences for avian embryology and immunology researches.
T cells in both the human and rabbit appendix are fewer than B cells. T cells in the rabbit appendix range from 7 to 40% (Dasso et al., 2000; Hanaoka et al., 1977; Bast et al., 1979) and in human from 19 to 50% (Kawanishi, 1987; Mizumoto, 1976; Neiburger et al., 1976; Alexopoulos et al., 1976). The majority of T cells lie in T-cell rich interfollicular region and a few are also found in B-cell rich follicular region. But peripheral CD4+CD8+ T-cells are reported in partially inbred and MHC-homozygous H.B15 chickens by Luhtala et al. (1997). Cell-mediated immunity plays a useful role in laying hens (Babu et al., 2005). In this study for the first time it was proved that CD4 or CD8 expression in goose bursa of fabricius is consistent with process in chicken thymus: co-negative CD4¯CD8¯ firstly emergence, then co-positive CD4+CD8+ formation, single-positive CD4+ or CD8+ appearance at last (Oguma et al., 2009; Parel and Chizzolini, 2004). The most interesting thing of all is T Lymphocyte selection in goose bursa of fabricius was unexpectedly similar to T cell development in thymus.
The results are approximately in accordance with that bursal CD4+, CD8+ T cells and macrophages were observed in the MDV-infected and controlled chicken. In addition, MDV-infected cells up-regulate MHC II molecules in bursa of fabricius (Niikura et al., 2007) and down-regulate MHC I after MDV infection (Gimeno et al., 2001; Hunt et al., 2001; Levy et al., 2003). Therefore, it can be concluded that T lymphocytes existed in avian bursa of fabricius and exerted an important role in B cell proliferation and immune system maintenance.
In this study, it was demonstrated that T Lymphocyte completely involved in the development of goose bursa of fabricius. It is well known that T cell function can be divided into three categories according to TCRαβ: T helper (Th1, Th2) and T suppressor (Ts); cytotoxic T lymphocyte (Tc). Th2 cells specialize in facilitating B-cell antibody responses, produce IL-4(drive B-cell proliferation), IL-5 and IL-13, while IL-4 and IL-5 enable IgE production (Larosa and Orange, 2008). It is well known that the surface receptor (CD28, CD40L and MHC etc.) on T lymphocytes is also surface antigen on B cell. The CD28 family ligand is B7-1 (CD80) and B7-2 (CD86). Previous studies showed that combination between IL-4 and CD40L:CD40 sustained B-cell activation and differentiation, promoted the memory B cell’s production and immunoglobulin class switch recombination (Larosa and Orange, 2008; Lafrenz and Feldbush, 1981; Miller and Sprent, 1971). Under these conditions the present study demonstrate that T lymphocytes is essential for B cell activation differentiation and maturation in the goose bursa of fabricius. Understanding the process of T lymphocyte selection in bursa of fabricius will greatly help study the immune response in goose.
The bursa of fabricius is one of the most important lymphoid system during cell apoptosis and its apoptosis rate is several fold higher than that of the thymus (Luna et al., 2005; Paramithiotis et al., 1995). This study uncovered that the distribution of the nucleus at different periods. A significant finding in the study was that goose bursal apoptosis decreased between E17 and 3d and increased after hatching. The apoptosis rate is very low while CD4+CD8+ co-expression level became higher. It was coincided with the fact that co-positive thymocytes were more sensitive than single-positive thymocytes via various reagents to induce apoptosis (Oguma et al., 2009). The apoptotic cells were also increased by virus inoculation (Wang et al., 2011). B-lymphocytes in the bursa of fabricius undergo proliferation, differentiation and apoptosis (Garcia-Espinosa et al., 2003). It has been estimated that only about 5% of these juvenile B-cells emigrate successfully, while the rest of the cells die in situ via apoptosis (Lassila, 1989). T-lymphocytes subsets may play a role in cellular immune response of colon cancer (Attallah et al., 2006). It was found that lymphocyte accompanied by extensive apoptotic cell death during avian development. The results indicated that T lymphocyte selection and apoptosis were correlated with B-cell development in goose bursa of fabricius, the concrete function is yet unclear and further study is needed to clarify the underlying sense.
CONCLUSION
It was proved that T-dependent area also existed in goose bursa of fabricius, involved B cell differentiation and maturation and shamed the same feature as the T lymphocyte selection in thymus. T lymphocytes are indispensible for the development of goose bursa of fabricius.
ACKNOWLEDGMENT
This study was supported by “863” Science and Technology Research Program of China (2007AA100504) and Anhui Natural Science Foundation (10020303043 and KJ2009A039).
APPENDIX
 | |
| Fig. S1: | B-H represents E17, E24, E26, E28, 3d, 7d, 18d, respectively. A represents goose incubation in oven at 37°C; I represent mature White goose |
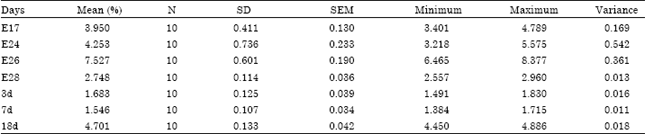
| Table S1: | The descriptions on double positive area percentage of geese bursa |
 | |
| Table S2: | The result of one-way ANOVA on double positive area percentage of geese bursa |
 | |
| Table S3: | The multiple comparisons on double positive area percentage of geese bursa with LSD |
 | |
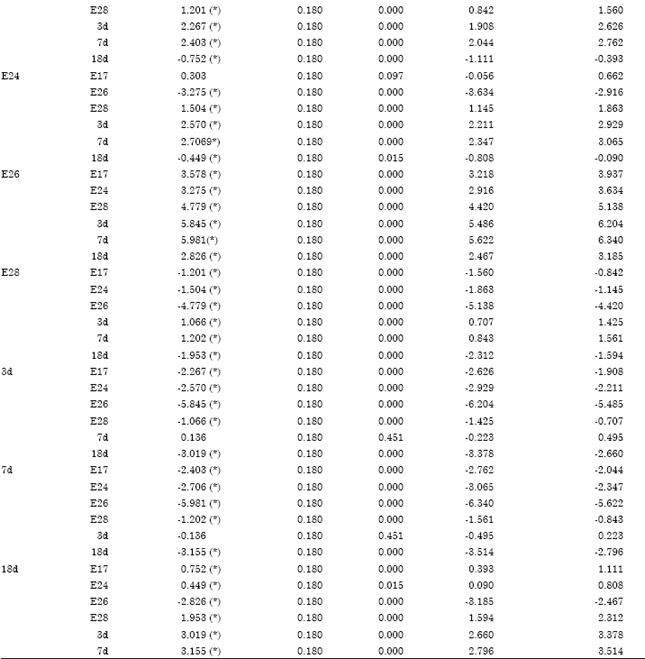 | |
| * The mean difference is significant at the 0.05 level | |
| Table S4: | The descriptions on apoptosis rate of geese bursa |
 | |
 | |
| Table S5: | The result of one-way ANOVA on apoptosis rate of geese bursa |
 | |
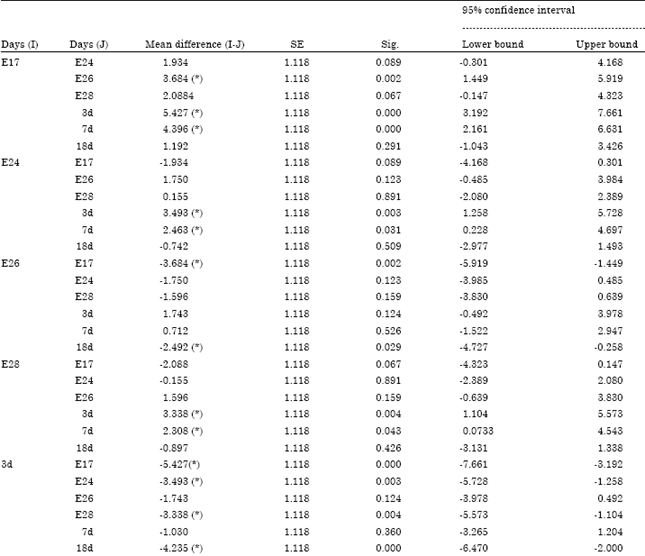
| Table S6: | The multiple Comparisons on apoptosis rate of geese bursa with LSD |
 | |
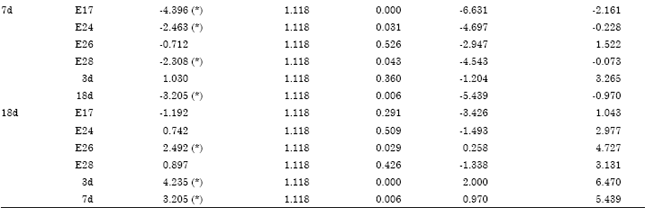 | |
| * The mean difference is significant at the 0.05 level | |
REFERENCES
- Attallah, A.M., A.A. Tabll, M.F. Ismail, A.S. Ibrahim and I. El-Dosoky, 2006. Flow cytometric analysis of peripheral blood T-lymphocyte subsets in colon cancer. Int. J. Cancer Res., 2: 409-414.
CrossRefDirect Link - Ackerman, G.A. and R.A. Knouff, 1959. Lymphocytopoiesis in the bursa of Fabricius. Am. J. Anat., 104: 163-205.
CrossRef - Alexopoulos, C., A.G. Papayannis and C. Gardikas, 1976. Increased proportion of B lymphocytes in human tonsils and appendices. Acta Haemat., 55: 95-98.
CrossRef - Wang, A., F. Liu, Z. Wang, X. Jiang, W. Wang, K. Teng and J. Xu, 2011. Pathological study of SPF chickens experimentally infected with a chinese IBDV strain BC6/85. Asian J. Anim. Vet. Adv., 6: 36-50.
CrossRefDirect Link - Babu, U.S., P.L. Wiesenfeld, R.B. Raybourne, M.J. Myers and D. Gaines, 2005. Effect of dietary fishmeal on cell-mediated immune response of laying hens. Int. J. Poult. Sci., 4: 652-656.
CrossRefDirect Link - Bast, B.J.E.G., D. Catty, R. Manten-Slingerland, J.T.G. Jansen, D.H. Veldhuis, P. Roholl and R.E. Ballieux, 1979. Surface Ig on rabbit lymphocytes, rabbit B and T-cells are distinct populations. Eur. J. Immunol., 9: 997-1003.
CrossRef - Brown, C.Y., S.J. Bowers, G. Loring, C. Heberden, R.M. Lee, P.E. Neiman, 2004. Role of Mtd/Bok in normal and neoplastic B-cell development in the bursa of Fabricius. Dev. Comp. Immunol., 28: 619-634.
CrossRef - Butler, J.E., 1997. Immunoglobulin gene organization and the mechanism of repertoire development. Scand. J. Immunol., 45: 455-462.
CrossRef - Cooper, M.D., W.A. Cain, P. Van Alten and R.A. Good, 1969. Development and function of the immunoglobulin producing system. Int. Arch. Allergy, 35: 242-252.
CrossRef - Cortes, A., J. Fonfria, A. Vicente, A. Varas, J. Moreno and A.G. Zapata, 1995. T-dependent areas in the chicken bursa of Fabricius: An immunohistological study. Anat. Rec., 242: 91-95.
PubMed - Dasso, J.F., H. Obiakor, H. Bach, A.O. Anderson and R.G. Mage, 2000. A morphological and immunohistological study of the human and rabbit appendix for comparison with the avian bursa. Dev. Comp. Immunol., 24: 797-814.
CrossRef - Dolfi, A., M. Lupetti, F. Bianchi and S. Michelucci, 1988. Diffusely infiltrated lymphoid areas of the bursa of Fabricius (DIA) and of the cloaca: An embryological study with morphological analogies. J. Anat., 156: 17-26.
CrossRef - Erf, G.F., W.G. Bottje and T.K. Bersi, 1998. CD4, CD8 and TCR defined T-cell subsets in thymus and spleen of 2- and 7-week old commercial broiler chickens. Vet. Immunol. Immunop., 62: 339-348.
CrossRef - Garcia-Espinosa, G., LA. Newberry, L.R. Berghman, B.M. Hargis, 2003. Differential cytoplasmic and whole cellular expression of histone H1 within the avian bursa of Fabricius. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol., 134: 505-509.
CrossRefPubMedDirect Link - Garcia-Espinosa, G., S. Clerens, L. Arckens, G.F. Erf, G. Tellez and B.M. Hargis, 2008. Peptides from the bursa of fabricius associated with suppression of mitogen stimulated DNA-synthesis in bursa of fabricius cells belong to intracellular proteins. Int. J. Poult. Sci., 7: 125-128.
CrossRefDirect Link - Gimeno, I.M., R.L. Witter, H.D. Hunt, L.F. Lee, S.M. Reddy and U. Neumann, 2001. Marek's disease virus infection in the brain:Virus replication, cellular infiltration, and major histocompatibility complex antigen expression. Vet. Pathol., 38: 491-503.
CrossRef - Glick, B., T.S. Chang and R.G. Jaap, 1956. The bursa of fabricius and antibody production. Poult. Sci., 35: 224-225.
CrossRefDirect Link - Griebel, P.J. and W.R. Hein, 1996. Expanding the role of Peyer's patches in B-cell ontogeny. Immunol. Today, 1: 30-39.
CrossRef - Hanaoka, M., R. Mizumoto and M. Takigawa, 1977. Regulation of IgM antibovine γ-globulin antibody formation by helper and suppressor T-cells in rabbits. Cellular. Immunol., 31: 1-12.
PubMed - Hunt, H.D., B. Lupiani, M.M. Miller, I. Gimeno, L.F. Lee and M.S. Parcells, 2001. Marek's disease virus down-regulates surface expression of MHC (B complex) class I (BF) glycoproteins during active but not latent infection of chicken cells. Viroly, 282: 198-205.
CrossRefDirect Link - Kawanishi, H., 1987. Immunocompetence of normal human appendiceal lymphoid cells: In vitro studies. Immunol., 60: 19-28.
PubMed - Lafrenz, D.E. and T.L. Feldbush, 1981. Role of T cells in the development of memory B cells. Quantitative and qualitative analysis. Immunol., 44: 177-186.
PubMed - Lassila, O., 1989. Emigration of B cells from chicken bursa of Fabricius. Eur. J. Immunol., 19: 955-958.
CrossRef - Levy, A.M., S.C. Burgess, I. Davidson, G. Underwood, G. Leitner, E.D. Heller, 2003. Interferon-containing supernatants increase Marek's disease herpesvirus genomes and gene transcription levels, but not virion replication In vitro. Viral. Immunol., 16: 501-509.
CrossRef - Luhtala, M., O. Lassila, P. Toivanen, O. Vainio, 1997. A novel CD4+CD8+ T-cell population: Inheritance of CD8a expression on CD4+ T-cells. Eur. J. Immunol., 27: 189-193.
CrossRef - Luna, M., N. Barraza, L. Berumen, M. Carranza, E. Pedernera, S. Harvey and C. Aramburo, 2005. Heterogeneity of growth hormone immunoreactivity in lymphoid tissues and changes during ontogeny in domestic fowl. Gen. Comp. Endocrinol., 144: 28-37.
CrossRef - Luna, M., A.J. Rodriguez-Mendez, L. Berumen, M. Carranza, J. Riesgo-Escovar, M.L. Baudet, S. Harvey and C. Aramburo, 2008. Immune growth hormone (GH): Localization of GH and GH mRNA in the bursa of Fabricius. Dev. Comp. Immunol., 32: 1313-1325.
CrossRef - Miller, J.F. and J. Sprent, 1971. Cell-to-cell interactions in the immune response VI. Contributions of thymusderived cells and antibody-forming cell precursors to immunological memory. J. Exp. Med., 134: 66-82.
CrossRef - Mizumoto, T., 1976. B and T-cells in lymphoid tissues of human appendix. Int. Arch. Allergy Appl. Immunol., 51: 80-93.
CrossRef - Nagy, N. and I. Olah, 2009. Locally applied testosterone is a novel method to influence the development of the avian bursa of Fabricius. J. Immunol. Methods., 343: 79-102.
CrossRef - Nagy, N. and I. Olah, 2010. Experimental evidence for the ectodermal origin of the epithelial anlage of the bursa of Fabricius. Development., 137: 3019-3023.
CrossRef - Neiburger, J.B., R.G. Neiburger, S.T. Richardson, J.L. Grosfeld, R.L. Baehner, 1976. Distribution of T and B lymphocytes in lymphoid tissue of infants and children. Infect. Immun., 14: 118-121.
PubMed - Niikura, M., T. Kim, H.D. Hunt, J. Burnside, R.W. Morgan, J.B. Dodgson and H.H. Cheng, 2007. Marek's disease virus up-regulates major histocompatibility complex class II cell surface expression in infected cells. Virology, 359: 212-219.
CrossRef - Odend'hal, S. and J.F. Breazile, 1980. An area of T cell localization in the cloacal bursa of White Leghorn chickens. Am. J. Vet. Res., 41: 225-228.
PubMed - Oguma, T., T. Ono, T. Kajiwara, M. Sato and Y. Miyahira et al., 2009. CD4+CD8+ thymocytes are induced to cell death by a small dose of puromycin via ER stress. Cellular. Immunol., 260: 21-27.
CrossRef - Olah, I. and B. Glick, 1978. The number and size of the follicular epithelium (FE) and follicles in the bursa of Fabricius. Poult. Sci., 57: 1445-1450.
PubMed - Paramithiotis, E., K.A. Jacobsen and M.J. Ratcliffe, 1995. Loss of surface immunoglobulin expression precedes B cell death by apoptosis in the bursa of Fabricius. J. Exp. Med., 181: 105-113.
CrossRef - Parel, Y. and C. Chizzolini, 2004. CD4+ CD8+ double positive (DP) T cells in health and disease. Autoimmun. Rev., 3: 215-220.
CrossRef - Reynolds, J.D. and B. Morris, 1983. The evolution and involution of Peyer's patches in fetal and postnatal sheep. Eur. J. Immunol., 13: 627-635.
CrossRef - Reynaud, C.A., C.R. Mackay, R.G. Muller and J.C. Weill, 1991. Somatic generation of diversity in a mammalian primary lymphoid organ: the sheep ileal Peyer's patches. Cell, 64: 995-1005.
CrossRef - Ricci, A., E. Bronzetti, L. Felici, E. Ciriaco, J.A. Vega and G. Germana, 1996. Muscarinic cholinergic receptor subtypes in the pigeon bursa of Fabricius: A radioligand binding and autoradiographic study. J. Neuroimmunol., 66: 23-28.
CrossRef - Rodriguez-Mendez, A.J., J.L., Luna-Acosta, M. Carranza, S. Harvey, C. Aramburo and M. Luna, 2010. Growth hormone expression in stromal and non-stromal cells in the bursa of Fabricius during bursal development and involution: Causal relationships. Gen. Comp. Endocrinol., 167: 297-307.
PubMed - Yamamoto, H., M. Hattori, K. Ohashi, C. Sugimoto and M. Onuma, 1996. Characterization of extrathymic T cells of chickens. Vet. Immunol. Immunopathol., 49: 375-386.
CrossRef - Liu, Y., X. Qiu, J. Xu, F. Hu and Y. Li et al., 2011. Association analysis between the polymorphism of the SLC11A1 gene and immune response traits in pigs. Asian J. Anim. Vet. Adv., 6: 580-586.
CrossRefDirect Link