
Catherine E. Stanley
Department of Biology and Center of Muscle Biology, University of Kentucky, Lexington, KY, 40506-0225, USA
Robin L. Cooper
Department of Biology and Center of Muscle Biology, University of Kentucky, Lexington, KY, 40506-0225, USA
LiveDNA: 1.523
Current Research in Neuroscience
Year: 2021 | Volume: 11 | Issue: 1 | Page No.: 1-17







ABSTRACT
Background and Objective: The effects of intracellular and extracellular pH changes on synaptic transmission have been studied for many years in different models. Intracellular acidification at the presynaptic terminal is known to occur with increased neuronal activity and can also occur in pathological conditions. This study aimed to study the effects of these pH alterations in larval Drosophila. Materials and Methods: The effects of pH on both spontaneous and evoked synaptic transmission at the Neuromuscular Junction (NMJ) in larval Drosophila were examined by altering intracellular acidification, using either propionic acid or the ammonium chloride pre-pulse technique as well as by altering extracellular pH using HCl or NaOH. This NMJ is glutamatergic and is used as a model of graded synaptic transmission. Electrophysiological recordings were obtained from muscle 6 at abdominal segment 3 or 4. Results were analyzed via Student’s t-test or repeated-measures ANOVA. Results: Propionic acid is shown to increase spontaneous quantal event frequency while also attenuating evoked transmission, enhancing high-frequency depression and depolarizing the target cell. The enhanced frequency of spontaneous events was attenuated with CdCl2 in the bath, indicating calcium influx as a mechanism. Ammonium chloride withdrawal has more diverse and complex effects that are shown to be dose and condition-dependent. Extracellular acidification depolarizes and alkalization hyperpolarizes the synaptic target. Conclusion: Thus, the effects of pH alteration on synaptic transmission are diverse in Drosophila melanogaster. These studies are significant in further developing a model of the effects of intracellular and extracellular acidification on evoked and non-evoked synaptic transmission.
PDF Abstract XML References Citation
Received: October 31, 2020;
Accepted: December 11, 2020;
Published: December 15, 2020
How to cite this article
Catherine E. Stanley and Robin L. Cooper, 2021. Effect of pH on Synaptic Transmission at the Neuromuscular Junction in Drosophila melanogaster. Current Research in Neuroscience, 11: 1-17.
DOI: 10.3923/crn.2021.1.17
URL: https://scialert.net/abstract/?doi=crn.2021.1.17
DOI: 10.3923/crn.2021.1.17
URL: https://scialert.net/abstract/?doi=crn.2021.1.17
INTRODUCTION
Factors that alter synaptic transmission are diverse, from external factors (i.e., neuromodulators, temperature) to intracellularly initiated actions such as metabolism. The increased electrical activity of a neuron is known to result in intracellular acidification, which is a decrease in intracellular pH (pHi). The decrease in pHi can then influence the local extracellular pH (pHo). Even with endogenous buffering of pHi and pHo to maintain cellular homeostasis, the cell and its environment undergo transient pH changes to cellular activity1-3.
In addressing the effects of pHo and pHi alterations on synaptic transmission, several indices are used. These include alterations in membrane potential, effects on evoked transmission, the rate of occurrence of spontaneous quantal events as well as ion and proton changes with electrical activity. Since large-scale, long-term pH changes are not generally seen during high neuronal activity, it is unlikely that pH drops enough to significantly impact the function of enzymes in the synaptic cleft that break down neurotransmitters. The transient alterations in pH, however, may impact cellular processes. The full effects of transient pH changes on synaptic transmission remain to be completely revealed.
The Drosophila Neuromuscular Junction (NMJ) is a commonly used model that is extremely useful for studies on synaptic transmission. Genetic manipulation is relatively straightforward in Drosophila, making it useful for studies involved in genetic research or for the use of such things as genetically encoded pH indicators (GEpHIs)4. The Drosophila NMJ allows for analysis of membrane potential and spontaneous as well as evoked synaptic transmission at the postsynaptic muscle, in addition to the tracking of vesicle dynamics within the presynaptic terminal5,6. Furthermore, due to the graded nature of excitatory junction potentials (EJPs) at the motor endplate, EJP amplitude can be correlated with the amount of neurotransmitter released presynaptically7,8.
Previous studies on the effects of pH in altering synaptic transmission using the Drosophila model found that modulation of pHo affects both spontaneous and evoked Excitatory Junctional Currents (EJCs). The finding that evoked rather than spontaneous EJCs were more significantly impacted by the pH change indicates that reduction in vesicular fusion and release of neurotransmitters is the main effect of acidification rather than alterations in the amount of neurotransmitter in each vesicle. Caldwell et al.9 further investigated the effects of acidification at the Drosophila NMJ by specifically targeting its effects on vesicular dynamics. Acidification was achieved by two methods: application of propionic acid and withdrawal of ammonium chloride. Propionic acid easily crosses the plasma membrane and can acidify the interior of the cell. Ammonium chloride application and incubation is known to alkalize the interior of the presynaptic terminal. Upon rapid withdrawal during washout, protons are left behind, leading to transient acidification of the intracellular space. Using propionic acid, Caldwell et al.9 found that miniature endplate potential (mEPP) frequency was significantly increased, while washout led to a decrease in mEPP frequency. The muscle membrane potential was also found to rapidly hyperpolarize upon propionic acid administration. The use of the ammonium chloride pre-pulse technique to acidify the presynaptic terminal showed more mixed results. Only some preparations showed an increase in mEPP frequency and frequencies were not affected by NHE inhibition.
In this study, the effect of altering pHi by propionic acid and ammonium chloride on spontaneous synaptic activity was investigated. The extracellular pH was also varied by HCl or NaOH to examine the effects on the target membrane potential. The effect of pHi on the rate of evoked synaptic depression was also investigated. These studies are significant in corroborating previous reported findings with similar techniques as well adding to the effect of synaptic depression with heightened activity to pH and repetitive exposures to altered pH.
METHODS AND MATERIALS
Study area: The study was carried out in the Department of Biology, University of Kentucky in Lexington, KY, from August, 2019-June, 2020.
Fly stock maintenance: Flies utilized were Canton-S (CS) strain Drosophila melanogaster obtained from Bloomington Fly Stock Center that has been maintained in the lab for many years. Early third instar larvae were used for all electrophysiological studies. Flies and larvae were fed a standard cornmeal-agar-dextrose-yeast medium diet, housed at temperatures between 20 and 22°C and maintained on a 12:12 L:D cycle.
Saline and pharmacology: For all dissected preparations, a modified hemolymph-like saline 3 (HL3) [in mM: 1.0 CaCl2·2H2O, 70 NaCl, 20 MgCl2, 5 KCl, 10 NaHCO3, 5 trehalose, 115 sucrose, 25 5N, N-bis(2-hydroxyethyl)-2-aminoethanesulfonic acid (BES)] at pH 7.1 was utilized as a bathing medium10,11.
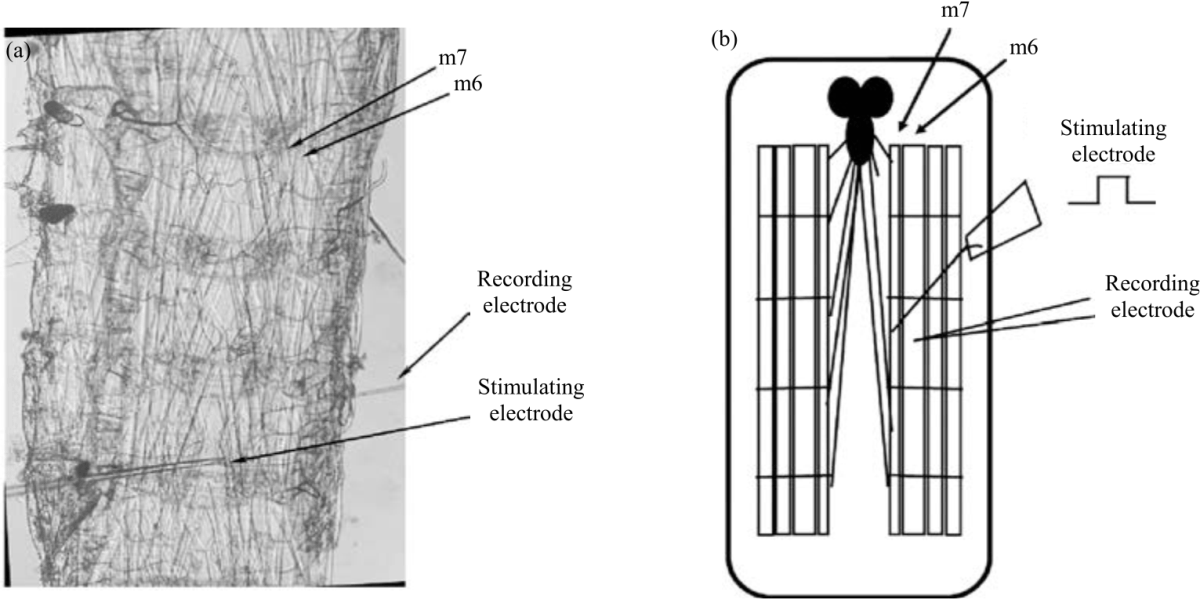 |
| Fig. 1(a-b): | Semi-intact dissection setup (a) The semi-intact dissected larval Drosophila preparation. Recordings were taken from muscle m6. In studies using stimulation, a suction electrode (labeled stimulating electrode) delivers stimulation to the appropriate segmental nerve and (b) A schematic representation of a) |
For studies using Propionic Acid (PA), a stock solution of 13.26 M/L PA was utilized and diluted with HL3 saline (20 mM PA: pH 5.70, 40 mM PA: pH 4.75). The NH4Cl (20 mM: pH 7.12, 40 mM: pH 7.15) and PA were made fresh before experimentation. Solutions were housed in closed vials during the experiments. For saline adjusted to a pH of 6.2 or 8.2, small amounts of HCl (1 M) stock or NaOH (1 M) stock was used, respectively. All compounds were obtained from Sigma-Aldrich (St. Louis MO, USA).
Electrophysiological recordings: Larvae were cut longitudinally on the dorsal side and the cuticle was peeled back and pinned on four corners. Internal organs were removed to expose the body wall muscles of the preparations. Segmental nerves were cut near the base of the brain and the nerves were maintained in the preparation to keep their connections to the muscle fibers. Immediately following dissection, postsynaptic recordings were taken with a sharp glass intracellular recording electrode filled with 3M KCl (40 mega ohm resistance).
Muscle 6 at abdominal segment 3 or 4 was used for electrophysiological recordings. Recordings were performed at 20-21°C with standard procedures12. An AxoClamp-2 B amplifier (Molecular Devices, LLC. 1311 Orleans Drive, Sunnyvale CA, USA) was used to obtain membrane potentials. PowerLab/4s interface (ADI Instruments, Colorado Springs, CO, USA) was used to digitize the electrical signals. A suction electrode was used for segmental nerve stimulation (S88 Stimulator, Astro-Med, Inc., GRASS Co., USA). Evoked excitatory junction potentials (EJPs), spontaneous miniature excitatory junction potentials (mEJPs) and membrane potentials were observed and analyzed with LabChart 7.0 (ADInstruments, USA) (Fig. 1a, b).
Measures of synaptic transmission and membrane potentials
Propionic acid: Spontaneous mEJPs were recorded for 5 min while bathing the dissected preparation in HL3 saline. The saline was then switched out with either 20 or 40 mM PA and allowed to bathe for another 5 min. The PA was then removed and the preparation was flushed with fresh saline twice. After recording for 5 min following washout, the saline was switched out with PA of the same concentration a second time and allowed to bathe the preparation for 5 min. The PA was removed and the preparation was flushed with saline two more times. The mEJPs were recorded for 5 more min following the final flushing (Fig. 2a).
Studies were also performed utilizing cadmium (CdCl2, 1 mM), a calcium channel blocker. Recordings were taken for 5 min while bathing the dissected preparation in HL3 saline. Total 1 mM CdCl2, was added to the preparation and allowed to bathe for 5 min. The solution was then removed and the preparation flushed with 20 mM PA and 1 mM CdCl2 combined and recorded for an additional 5 min (Fig. 2b).
Ammonium chloride: Spontaneous mEJPs were recorded for 5 min from the dissected preparation bathed in HL3 saline. The preparation was then incubated for 20 min in 20 mM NH4Cl. Spontaneous events were recorded for an additional 10 min following the rapid removal of NH4Cl (Fig. 2c).
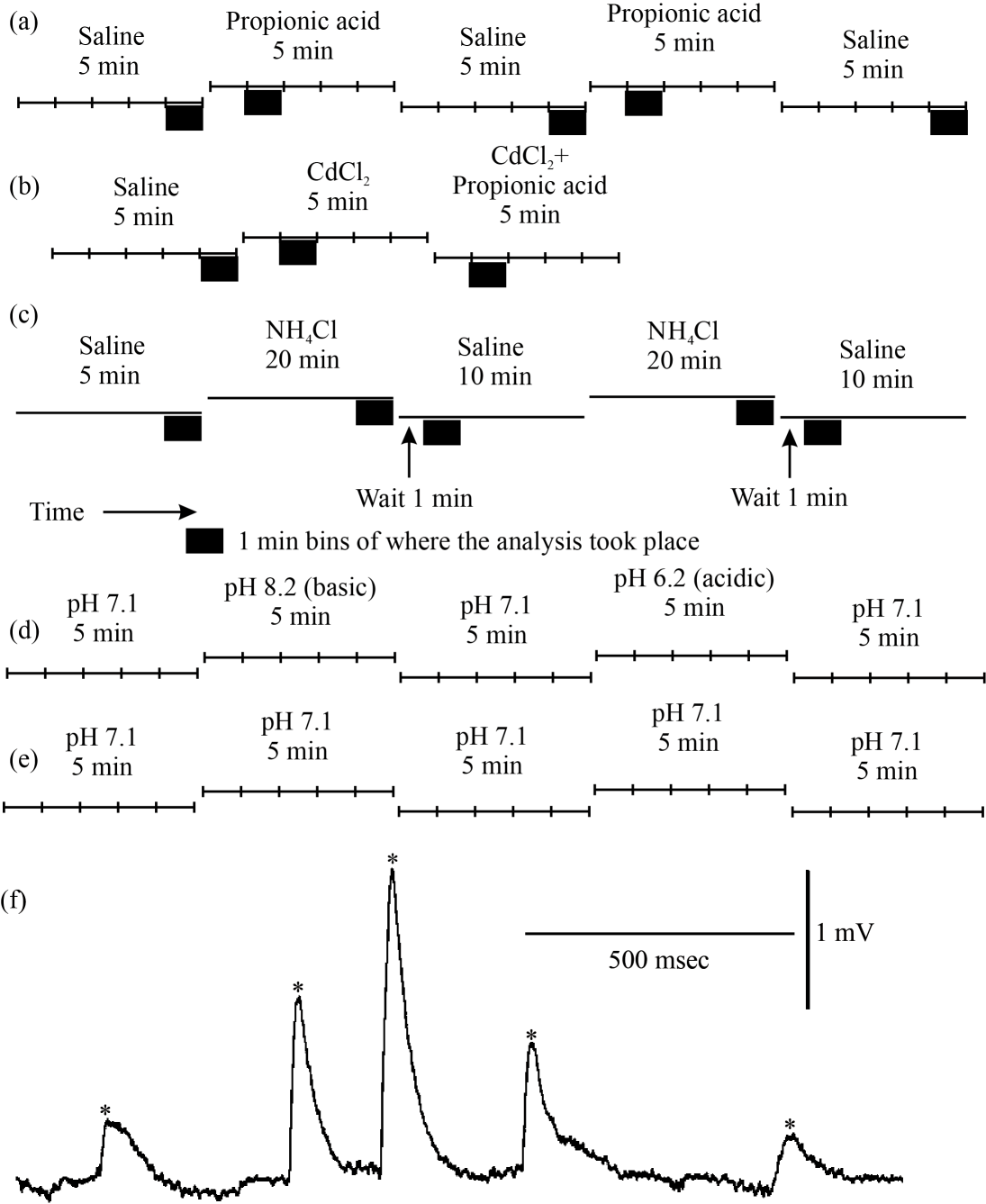 |
| Fig. 2(a-f): | Timelines for experimental protocols and analysis windows of spontaneous mEJP studies with alteration in intracellular and extracellular pH The timelines for each experiment in the mEJP studies are shown. The black boxes indicate the 1 min windows in which mEJP frequency was calculated. (a) Protocol for the effect of propionic acid on spontaneous mEJPs, (b) Protocol for the effect of CdCl2 in combination with propionic acid on spontaneous mEJP frequency, (c) Protocol for the effect of ammonium chloride on spontaneous mEJP frequency, (d) Protocol for the effect of extracellular basic or acidic pH, (e) protocol for the control condition in which saline of the same pH was exchanged and (f) An example of spontaneous quantal events. A representative trace of spontaneous quantal events, were seen as rapid deflections, versus the baseline recording. Quantal events are marked by asterisks (*). The deflections have a characteristic fast rise followed by slow decay |
Altering extracellular pH environment: To replicate the results of Caldwell et al.9 and to add experimentation as to the effects on membrane potential with changes in extracellular pH, a series of experiments were performed in which the resting membrane potential was monitored while altering extracellular pH. The paradigm used was 5 min in saline at pH 7.1, 8.2, 6.2, followed by 5 min at pH 7.1 (Fig. 2d).
Control experiments for changing the bathing media were performed by exchanging the saline at pH 7.1 for fresh media at the same pH. Five periods of 5 min of bathing between changes was used (Fig. 2e). This mimicked the timing for the exposure to basic and acidic environments.
Quantal analysis: Quantal events were considered as synaptic activity above two times the average background noise level and with a sharp rise time. The frequency of the mEJPs was measured by manually identifying the events and by placing a cursor at the base of each event in set time windows (Fig. 2f).
Membrane potentials: Resting membrane potentials (RPs) were analyzed for the various paradigms along with studies on the frequency of spontaneous mEJPs. Membrane potential values were taken from the end of each saline or treatment condition and compared across conditions.
Evoked EJPs: The evoked EJPs were induced by stimulating the segmental nerve with a fire-polished glass suction electrode and Grass S-88 stimulator. Amplitudes were measured manually by placing a cursor at the base and peak of the EJP using the LabChart software. EJP amplitudes were corrected for non-linear summation using Martin's correction factor, for the EJP amplitudes larger than 15% of the resting potential by assuming a reversal potential of 0 mV and a membrane capacitance factor of 0.5513,14.
 |
| Fig. 3: | Protocol timeline for 50% synaptic depression Using an intracellular recording electrode, the resting membrane potential of the muscle fiber is determined. Half Hz stimulation frequency is applied to the associated segmental nerve through a suction electrode to determine the minimum voltage necessary for the maximal response of the muscle such that all nerves are recruited. Stimulation frequency is then increased to 5 Hz and the initial EJP amplitude is determined from the average of the first five full EJPs. Stimulation at 5 Hz is continued until the average amplitude of EJPs is 50% of the initial EJP amplitude |
Depression studies: Third instar larvae were utilized to determine the amount of time it takes for 50% depression of EJPs amplitudes upon continuous stimulation. To determine the minimum voltage necessary to produce maximal EJP response upon stimulation, the segmental nerve was stimulated at 0.5 Hz while continually adjusting the voltage supplied. Upon determining the minimum voltage to generate evoked EJPs, 20 mM PA was applied to the preparation and allowed to bathe for 1 minute while stimulated at 0.5 Hz. The stimulation rate was then increased to 5 Hz to produce continuous EJPs at a high rate. As the membrane potential rapidly depolarizes upon the addition of PA, the initial EJP amplitude was determined as the amplitude of the first EJP following stabilization of the membrane potential. Continuous stimulation was applied until the average amplitude of the EJPs being produced was 50% of the initial EJP amplitude. The time it took from the start of stimulation to 50% depression was recorded. A timeline of the protocol can be seen in Fig. 3.
To determine the possible effects of the changing membrane potential on the initial depression study recordings, the experiment was repeated while voltage clamping the membrane potential at -60 mV. Voltage was maintained by injecting current during all periods in which it is clamped at -60 mV.
Statistical analysis: A paired Student's t-test or repeated-measures ANOVA was used to compare the difference in the number of spontaneous quantal events before and after exchanging solution with saline containing PA, NH4Cl, NaOH, or HCl. Differences in the resting membrane potentials were measured before and after added these same compounds and analyzed by a repeated-measures ANOVA.
A repeated-measures ANOVA was used to examine the before and after data and if the data sets were normally distributed. A Bonferroni test was used as a post hoc test following the ANOVA to compare the relative changes induced by the compounds within an experimental paradigm to determine significant differences. This analysis was performed with Sigma Stat software. A p<0.05 is considered as statistically significant with an asterisk (*).
RESULTS
Propionic acid: A representative trace for the effect of 20 mM propionic acid on spontaneous quantal events illustrates a notable pattern of depolarization upon application of propionic acid, along with an increase in mEJP frequency (Fig. 4a). Subsequent washout with fresh saline was shown to re-polarize the membrane potential and reduce mEJP frequency. The second application of propionic acid and washout with saline repeated the pattern of depolarization and the increase in the frequency of the mEJPs followed by repolarization and reduction in the mEJP frequency. A similar pattern was observed when using 40 mM propionic acid (Fig. 4b). It is important to note, however, that in many of the preparations with 40 mM PA it was difficult to maintain an intracellular recording in the muscle fiber due to contractions. The use of 1 mM CdCl2 to block voltage-gated calcium channels in the presynaptic membrane reduced the occurrence of spontaneous quantal events without affected the membrane potential of the muscle fiber. The following application of 1 mM CdCl2 with 20 mM propionic acid increased the frequency in some of the preparations but not at all. However, the membrane did depolarize in all trials (Fig. 4c).
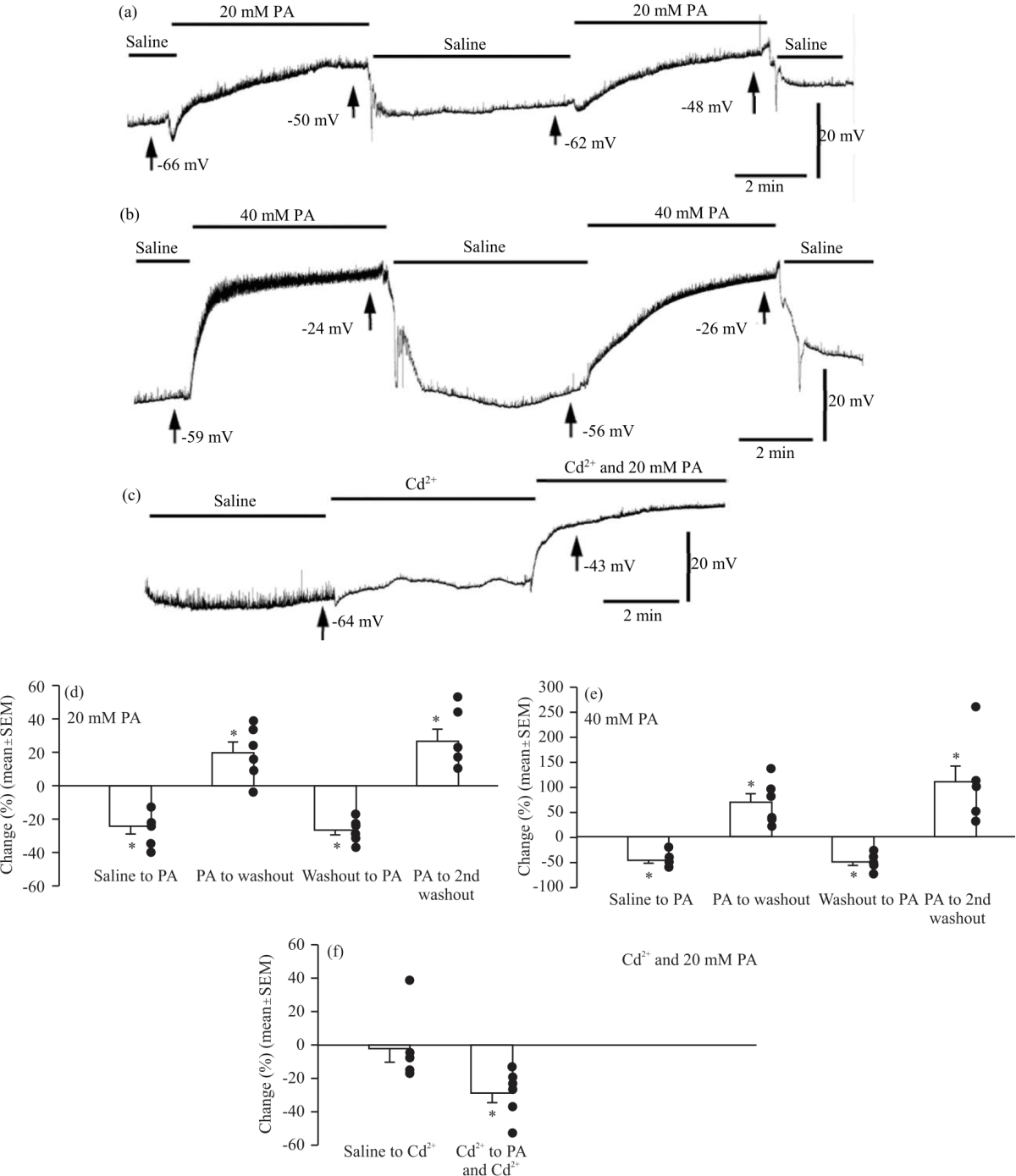 |
| Fig. 4(a-f): | Representative traces for the effect of propionic acid on spontaneous quantal events and membrane potentials (a) Representative traces of spontaneous quantal events during 5 min of saline, 5 min after application of 20 mM propionic acid, 5 min after washout with saline, 5 min after a second treatment with 20 mM propionic acid and 5 min after a final washout with saline, (b) Representative trace of spontaneous quantal events using the same protocol as for A) but with 40 mM propionic acid, (c) Representative trace of spontaneous quantal events during 5 min of saline, 5 min after application of 1 mM CdCl2 and 5 min after application of 1 mM CdCl2 and 20 mM propionic acid. Percent change in membrane potential values with propionic acid was calculated between each changing condition. Membrane potential values were taken from the end of each condition, (d) The average±SEM percent change in membrane potentials between the initial saline condition and after application of 20 mM propionic acid, between the first application of 20 mM propionic acid and the first washout, between the first washout and the second application of 20 mM propionic acid, and between the second application of 20 mM propionic acid and the final washout, (e) The average±SEM percent change in membrane potentials using the same protocol as (A) but with 40 mM propionic acid and (f) The average±SEM percent change in membrane potentials between the initial saline condition and after application of 1 mM CdCl2 and between the 1 mM CdCl2 condition and after application of 1 mM CdCl2 with 20 mM propionic acid. Significance of p<0.05 is denoted by an *. The closed circles represent individual preparations. Significance values are calculated based on raw data |
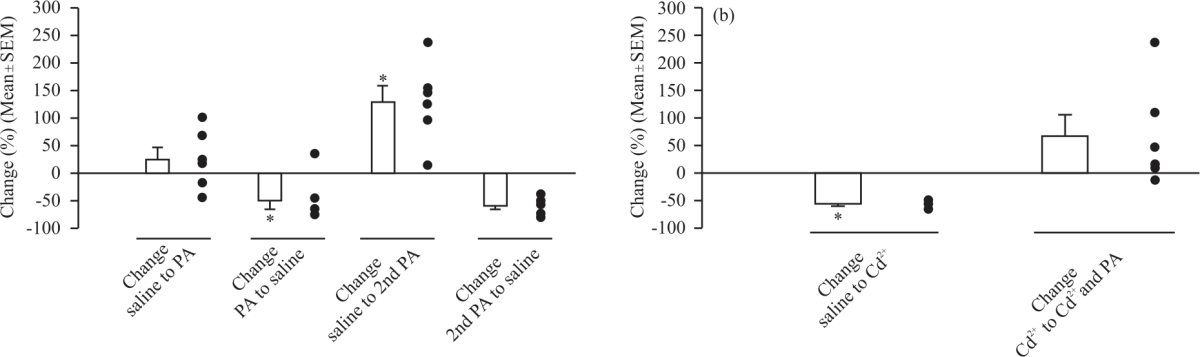 |
| Fig. 5: | Percent change in spontaneous quantal event frequency with propionic acid Percent change values in spontaneous quantal event frequency were calculated between each changing condition, (a) The average±SEM percent change in spontaneous quantal event frequency between the initial saline condition and after application of 20 mM propionic acid, between the first application of 20 mM propionic acid and the first washout, between the first washout and the second application of 20 mM propionic acid, and between the second application of 20 mM propionic acid and the final washout and (b) The average±SEM percent change in spontaneous quantal event frequency between the initial saline condition and after application of 1 mM CdCl2 and between the 1 mM CdCl2 condition and after application of 1 mM CdCl2 with 20 mM propionic acid. Significance of p<0.05 is denoted by an *. The closed circles represent individual preparations. Significance values are calculated based on raw data |
The mean RP was significantly depolarized after the first application of 20 mM propionic acid (-43.67±2.23 mV, one-way ANOVA with a Bonferroni post hoc analysis, N = 6) compared to the initial saline condition (-58.33±2.64 mV, p<0.001). RP became significantly more negative following washout (-51.67±2.32 mV, p = 0.029). RP was then significantly depolarized after the second application of 20 mM propionic acid (-37.67±2.03 mV, p<0.001). The second and final washout made the RP significantly more negative (-47.17±2.32 mV, p = 0.007). Thus, application of 20 mM propionic acid depolarized the RP but washout with saline was able to repolarize the RP. It is important to note that the final washout RP was still significantly depolarized compared to the initial saline value (p = 0.001). The trends in percent change of the RP between each condition were shown in Fig. 4d.
The mean RP was also significantly depolarized after the first application of 40 mM propionic acid (-34.17±3.60 mV) compared to the initial saline condition (-64.33±4.04 mV, p<0.001, one-way repeated measures ANOVA and the Bonferroni post hoc analysis, Fig. 4e). The RP then became significantly more negative following the first washout (-54.50±2.25 mV, p<0.001). The application of 40 mM propionic acid for a second time significantly depolarized the RP (-27.17±3.61 mV, p<0.001). The second and final washout made the RP significantly more negative (-51.67±1.78 mV, p<0.001, Fig. 4e). It is important to note that the final washout RP is still significantly depolarized compared to the initial saline value (p = 0.017). Therefore, similar to treatment with 20 mM propionic acid, 40 mM propionic acid application depolarized RP but washout was effective in repolarizing the RP, although not necessarily to initial RP values. The Fig. 4e depicts the trends in percent change of the RP between each condition. It does not appear that Ca2+ influx is responsible for the depolarization induced by PA (20 mM) as preincubation with Cd2+ and then PA with Cd still results in depolarization (Fig. 4c-f). A comparison of the percent change in membrane potential between saline and propionic acid application for 20 mM versus 40 mM propionic acid indicates that 40 mM propionic acid depolarized RP to a significantly greater extent than 20 mM propionic acid (N = 6, p = 0.019, Student’s t-test).
There was a significant decrease in mEJP frequency between the first 20 mM propionic acid treatment and the first washout (0.72±0.15 Hz, N = 6, p = 0.010). There was then a significant increase in mEJP frequency after the second application of 20 mM propionic acid (1.60±0.40 Hz, N = 6, p<0.05, paired t-test). There was no significant difference in mEJP frequency after the final washout (0.65±0.20 Hz, N = 6, p>0.05) (Fig. 5a). To determine if the increase in the frequency of mEJPs was due to propionic acid promoting an influx of extracellular calcium, a separate set of preparations were exposed to CdCl2 (1 mM) to block voltage-gated calcium channels on the plasma membrane. There was no significant difference in the mean RP between the initial saline condition (-64.50±3.53 mV) and after application of 1 mM CdCl2 (-61.67±1.33 mV, repeated measures one-way ANOVA and the Bonferroni post hoc analysis). Application of 1 mM CdCl2 in combination with 20 mM propionic acid led to significant depolarization (-43.67±3.23 mV) compared to both the initial saline condition and the 1 mM CdCl2 condition (Fig. 4c-f).
The mean frequency of mEJPs was significantly decreased between the initial saline condition (2.98±0.19 Hz) and treatment with CdCl2 (1.31±0.11 Hz, N = 6, p<0.05, paired Student's t-test). For a comparison of mEJP frequency between the CdCl2 condition and CdCl2 combined with 20 mM propionic acid, there was not a significant difference (N = 6, p = 0.063, Wilcoxon Signed Rank test was used as the Shapiro-Wilk test of normal distribution failed) (Fig. 5b).
Time to depression of the EJP amplitude to 50% was utilized as a means of examining the effect of propionic acid on evoked synaptic transmission at a high frequency. A representative trace of the depression in the EJP amplitude in saline as a control depicts a lengthy rundown time (Fig. 6a). In contrast, a representative trace for 50% EJP amplitude depression following administration of 20 mM propionic acid in a non-voltage-clamped preparation illustrates a rapid rundown to 50% of the initial EJP amplitude (Fig. 6b). This pattern is repeated in voltage-clamped preparations that are clamped to -60 mV (Fig. 6c).
The mean time to depression of 50% in the EJP amplitude was significantly decreased in the non-voltage clamped condition (N = 6, p<0.001, Kruskal-Wallis one way of variance on ranks, all pairwise comparisons Dunn's method) compared to the saline condition (2,962±400 sec, ±SEM). There was also a significant decrease in time between the saline and voltage-clamped condition (56±45 sec, ±SEM, N = 5, p<0.007, Kruskal-Wallis one way of variance on ranks, all pairwise comparisons Dunn's method). There was no significant difference in the time between the non-voltage clamped and voltage-clamped 20 mM propionic acid preparations (N = 6, p = 0.792, Mann-Whitney rank-sum test was used as the Shapiro-Wilk test of normal distribution failed) (Fig. 6d).
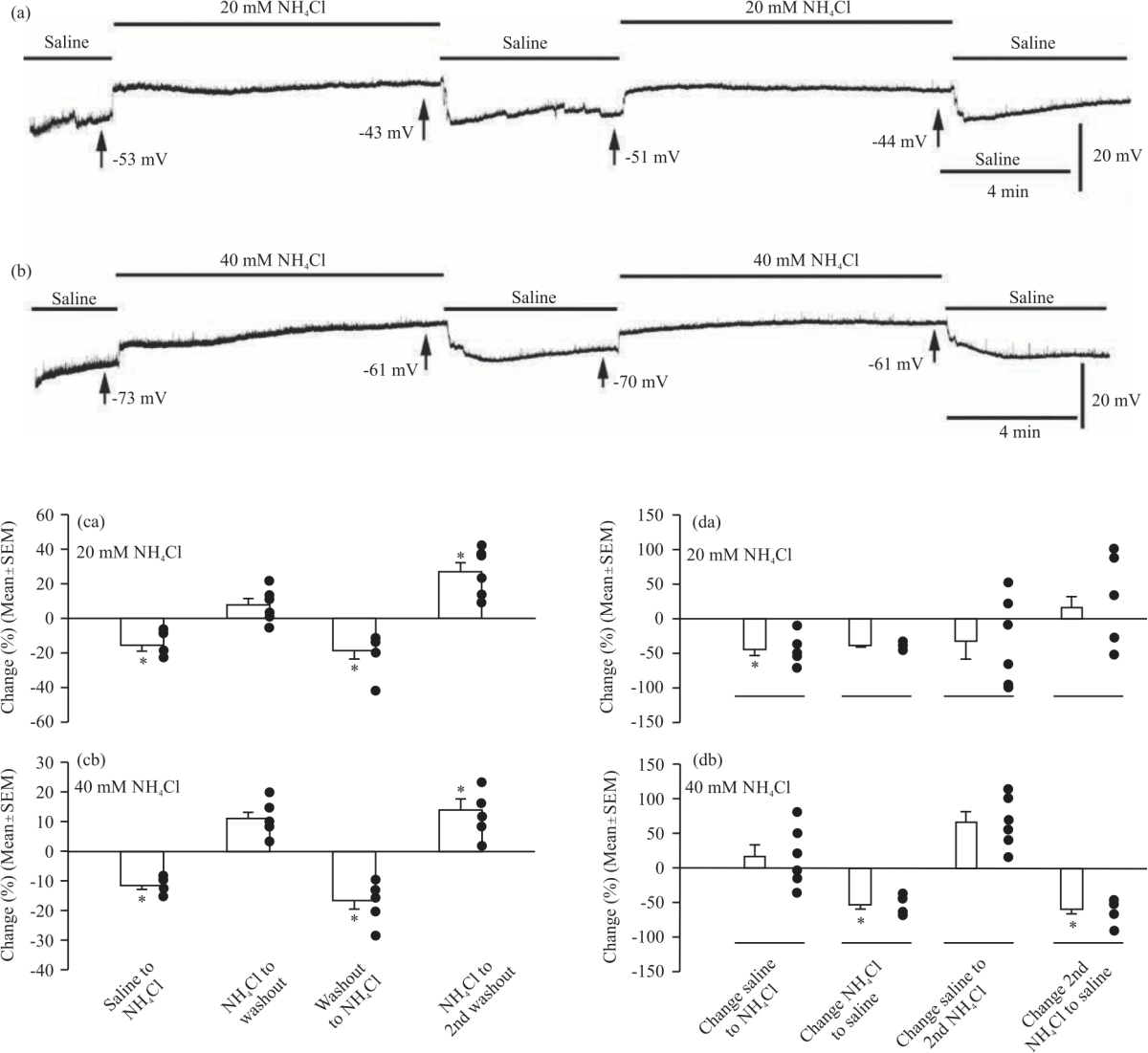
Ammonium chloride: A representative trace for spontaneous quantal events shows depolarization and a reduction in the frequency of mEJPs after the addition of ammonium chloride (20 mM), whereas its removal causes repolarization and a further reduction in the frequency of mEJPs (Fig. 7a). A similar pattern is seen for the 40 mM ammonium chloride condition (Fig. 7b).
The RP was significantly increased after the first application of 20 mM ammonium chloride (-51.50±3.42 mV) compared to the initial saline condition (-61.00±2.76 mV, p = 0.028, a one-way ANOVA and the Bonferroni post hoc analysis). Analysis by ANOVA was unable to compare the difference in mean RP between the first application of ammonium chloride and the first washout due to non-normality. Analysis by paired Student's t-test indicated no significant difference between the two conditions (p = 0.452). RP was significantly increased after the second application of 20 mM ammonium chloride (-44.17±2.32 mV, p = 0.012). The second and final washout significantly hyperpolarized the RP (-55.50±2.09 mV, p = 0.006). Thus, application of 20 mM ammonium chloride depolarized the RP but washout with saline was able to repolarize the RP. The Fig. 7ca depicts the trends in percent change of the RP between each condition.
The RP was significantly depolarized after the first application of 40 mM ammonium chloride (-57.00±2.88 mV) compared to the initial saline condition (-64.50±3.41 mV, p = 0.011, one-way repeated measures ANOVA and the Bonferroni post hoc analysis). The change in RP between the first application of 40 mM ammonium chloride and the first washout (-63.17±3.10 mV) was very near significance (p = 0.051). Application of 40 mM ammonium chloride for a second time significantly depolarized the RP (-52.67±3.08 mV, p<0.001). The second and final washout made the RP significantly more negative (-59.83±3.06 mV, p = 0.016). Thus, there was a similar pattern of depolarization after the application of ammonium chloride and repolarization upon washout for both 20 mM and 40 mM concentrations of ammonium chloride. A comparison of the percent change in membrane potential between saline and ammonium chloride application for 20 mM versus 40 mM ammonium chloride indicates that there was no significant difference in the extent of RP depolarization between doses (p = 0.18). The Fig. 7cb depicts the trends in percent change of the RP between each condition.
In analyzing the effects of 20 mM ammonium chloride on spontaneous quantal events, the mean frequency of mEJPs was found to be significantly decreased between the initial saline condition (2.61±0.29 Hz, ±SEM) and after application of 20 mM ammonium chloride (1.43±0.26 Hz, N = 6, p<0.05, a one-way ANOVA and the Bonferroni post hoc analysis). There was no significant change in mEJP frequency following the first removal of 20 mM ammonium chloride (0.85±0.14 Hz, N = 6, p>0.05). There was also no significant difference in mEJP frequency upon the second application of 20 mM ammonium chloride (0.62±0.29 Hz, N = 6, p>0.05, paired t-test) or after the final washout (0.49±0.12 Hz, N = 6, p>0.05, paired t-test).
The effects of 40 mM ammonium chloride application and withdrawal showed slightly different trends to that seen in the preparations exposed to the lower concentration. The mean frequency of mEJPs was not significantly different between the initial saline condition (3.69±0.52 Hz) and the first application of 40 mM NH4Cl (3.97±0.43 Hz, N = 6, p>0.05, paired t-test). However, there was a significant decrease in mEJP frequency following ammonium chloride removal (1.79±0.22 Hz, N = 6, p<0.05, a one-way ANOVA and the Bonferroni post hoc analysis).
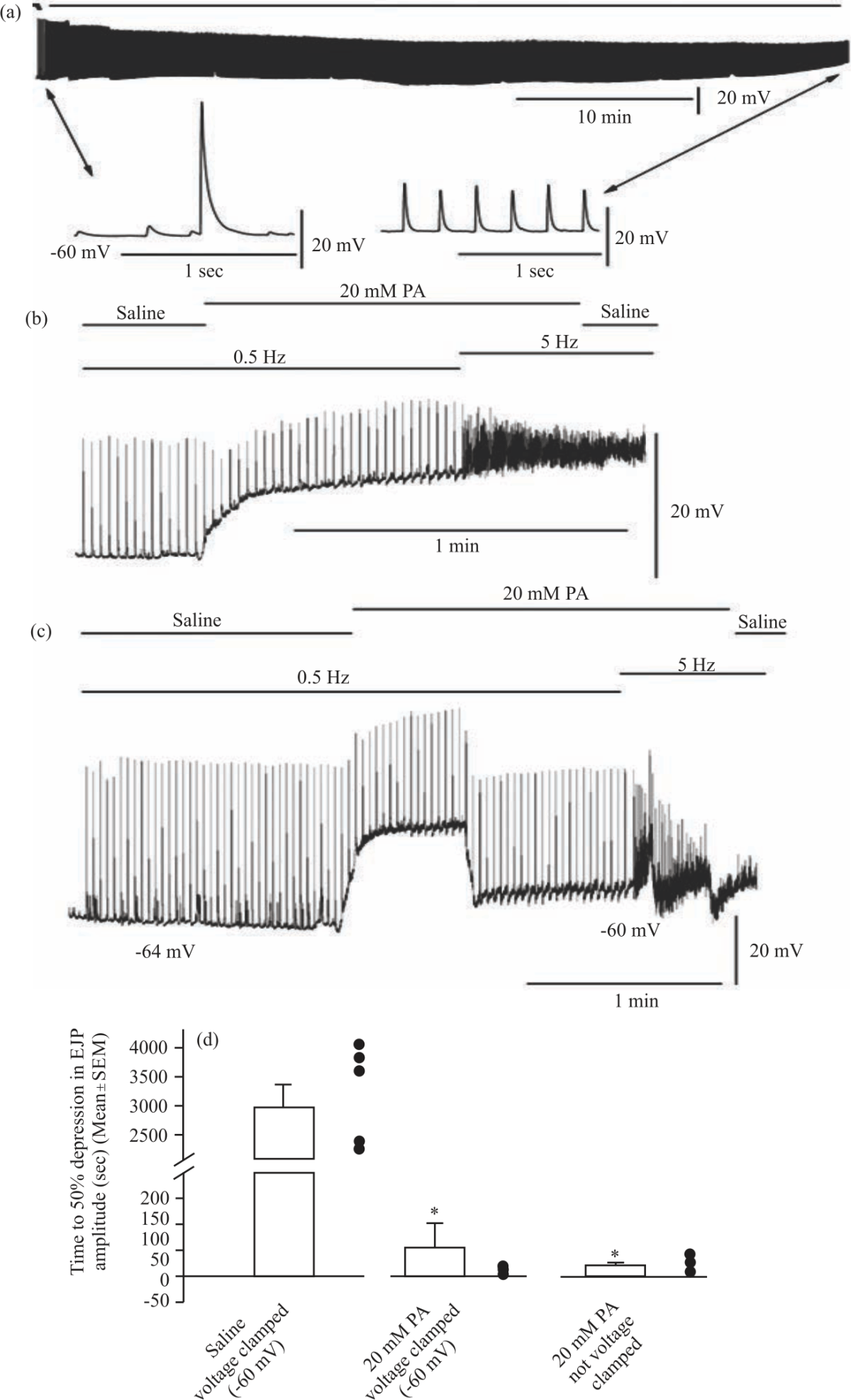 |
| Fig. 6(a-d): | Representative traces for time to 50% EJP amplitude depression For each preparation, the initial EJP amplitude was determined from the first full EJP upon the start of 5 Hz stimulation. Each preparation was observed until the average amplitudes of the evoked EJPs were 50% of the initial EJP amplitude from that preparation. Note the difference in scale bars for each trace, (a) Representative trace of the time to 50% EJP amplitude depression in saline only, (b) Representative trace of the time to 50% EJP amplitude depression in a non-voltage clamped preparation after the addition of 20 mM propionic acid, (c) Representative trace of the time to 50% EJP amplitude depression in a voltage-clamped preparation after the addition of 20 mM propionic acid and (d) Time to 50% EJP depression using 5 Hz stimulation. The Mean±SEM time (in seconds) for 50% EJP amplitude depression during 5 Hz stimulation for saline controls, after the addition of 20 mM propionic acid for non-voltage-clamped preparations and after addition of 20 mM propionic acid for voltage-clamped preparations. *Indicates a significant difference (p<0.05) from the saline condition. The closed circles represent individual preparations |
 |
| Fig. 7(a-d): | Representative traces for the effect of ammonium chloride on spontaneous quantal events (a) Representative traces of spontaneous quantal events during 5 min of saline, 20 min after application of 20 mM ammonium chloride, 10 min after washout with saline, 20 min after a second treatment with 20 mM ammonium chloride, and 10 min after a final washout with saline, (b) Representative trace of spontaneous quantal events using the same protocol as for A) but with 40 mM ammonium chloride. Percent change values in membrane potentials were calculated between each changing condition. Membrane potential values were taken from the end of each condition, (ca) The average±SEM percent change in membrane potentials between the initial saline condition and after application of 20 mM ammonium chloride, between the first application of 20 mM ammonium chloride and the first washout, between the first washout and the second application of 20 mM ammonium chloride, and between the second application of 20 mM ammonium chloride and the final washout, (cb) The average±SEM percent change in membrane potentials using the same protocol as (ca) but with 40 mM ammonium chloride. Significance of p<0.05 is denoted by an *. The closed circles represent individual preparations. Percent change values in spontaneous quantal event frequency were calculated between each changing condition and (da) The average±SEM percent change in spontaneous quantal event frequency between the initial saline condition and after application of 20 mM ammonium chloride, between the first application of 20 mM ammonium chloride and the first washout, between the first washout and the second application of 20 mM ammonium chloride, and between the second application of 20 mM ammonium chloride and the final washout, (db) The average±SEM percent change in spontaneous quantal event frequency using the same protocol as (da) but with 40 mM ammonium chloride. Significance of p<0.05 is denoted by an *. Significance values are calculated based on the raw data |
There was no significant change in mEJP frequency after a second application of ammonium chloride (2.89±0.30 Hz, N = 6, p>0.05). Removal of ammonium chloride for the second time resulted in a significant reduction in mEJP frequency (1.19±0.26 Hz, N = 6, p<0.05). The Fig. 7da-db depicts the trends for percent change in mEJP frequency across conditions.
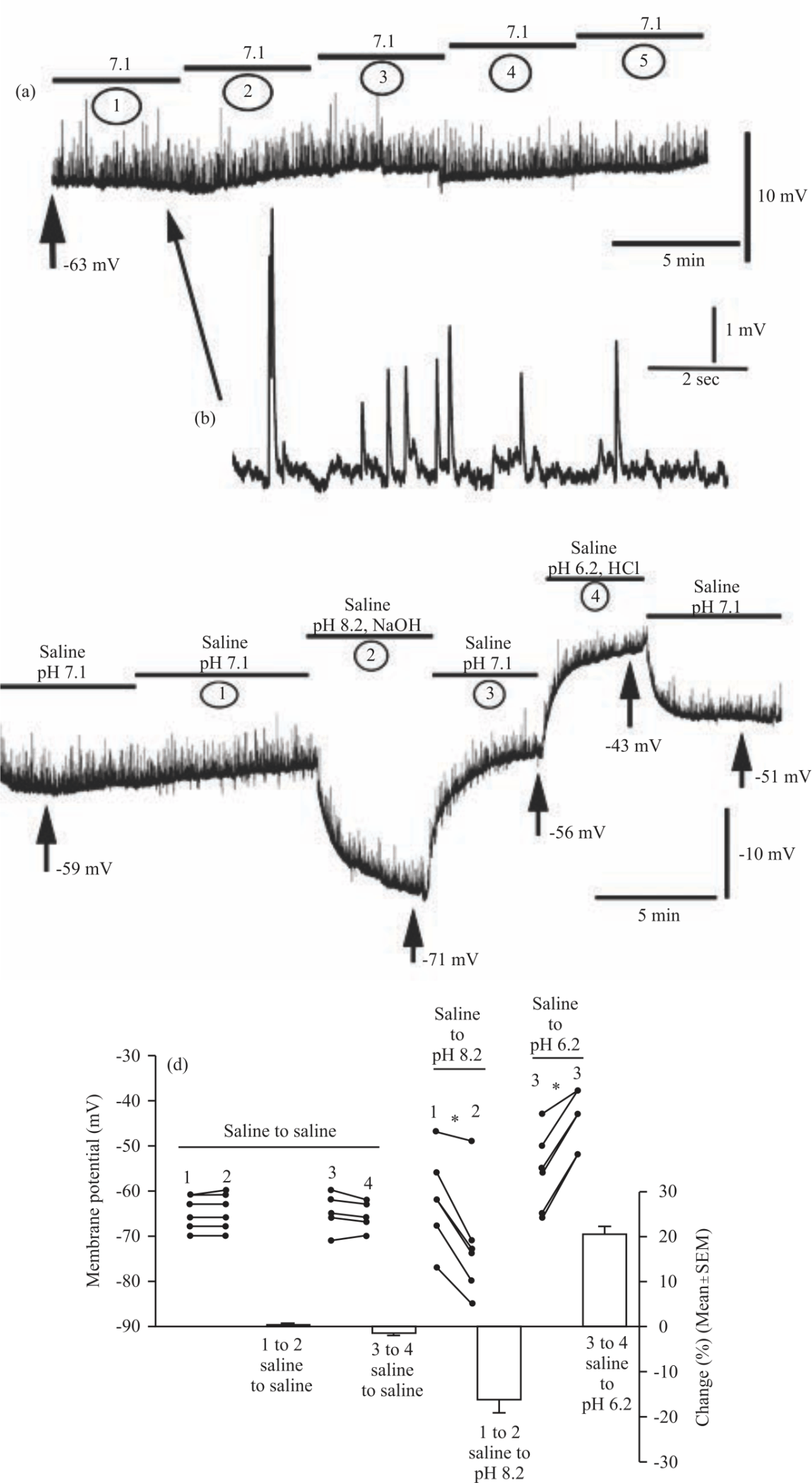 |
| Fig. 8(a-d): | A representative trace for the saline control paradigm in which saline at a pH of 7.1 was exchanged four times every 5 min (a) A full trace with the five saline exchanges marked as bars, (b) An enlarged trace depicting mEJP frequency and amplitude before the first exchange, (c) A pH of 8.2 hyperpolarizes the RP whereas a pH of 6.2 depolarizes the RP. Both effects on the RR were able to be reversed upon exchange with fresh saline at a pH of 7.1 and (d) The changes in membrane potential between saline controls and before and after changes in pH from 7.1-8.2 or from 7.1-6.2. Raw values of membrane potential from individual traces are shown as closed circles. Mean percent change values between the changing conditions are shown as solid bars. Significance of p<0.05 is denoted by an * |
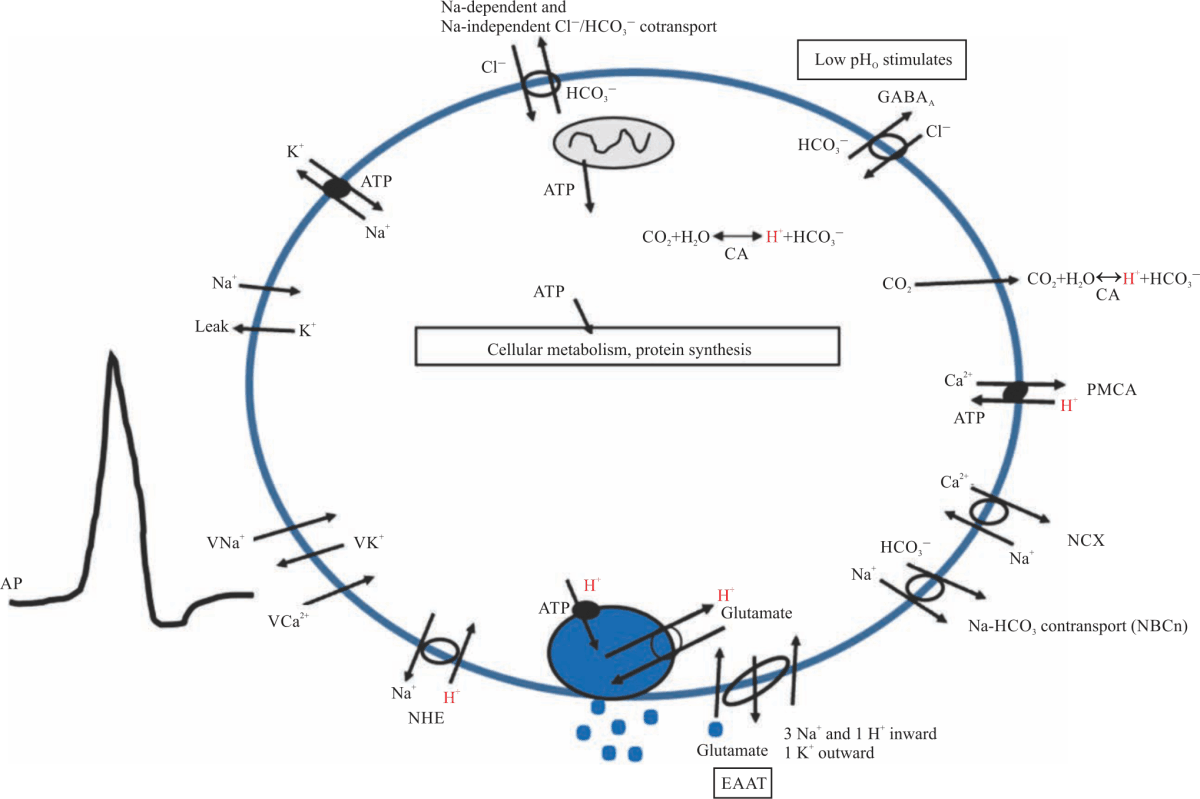 |
| Fig. 9: | A model of a presynaptic bouton A simplified model of a presynaptic bouton of a motor neuron with some of the many transporters, exchangers, channels and ions that may be implicated in the effects of pH on synaptic transmission. Included is a vesicle (shown as a filled blue circle) to depict the fusion and release of glutamate into the synaptic cleft. Pictured are aspects in the plasma membrane such as the sodium-hydrogen exchanger (NHE), the plasma membrane calcium ATPase (PMCA), the sodium-calcium exchanger (NCX) as well as multiple cotransporters, antiporters, leak channels, and voltage-gated channels for various ions |
Extracellular pH effects on RP: The change in extracellular pH with compounds not readily diffusible across the membrane also revealed alterations in the RP. As a control for the effects of exchanging the media multiple times, saline at 7.1 was exchanged four times with 5 min maintained between changes (Fig. 8a). An enlarged portion of the synaptic activity for this control is also shown (Fig. 8b). There were no significant differences in mean RP in this control condition.
The effect of a changing environment from relatively neutral to basic then back to neutral and then acidic for short acute periods revealed an effect on RP (Fig. 8c). The mean RP was significantly more negative at a pH of 8.2 (-72.00±5.06 mV, one-way ANOVA with a Bonferroni post hoc analysis, N = 6) compared to the initial pH of 7.1 (-62.00±4.17 mV, p = 0.010). RP then depolarized after switching back to saline at a pH of 7.1 (-55.83±3.59 mV, p<0.001). RP was significantly depolarized further when pH was decreased to 6.2 (-44.33±2.59 mV, p = 0.003). Switching back to a pH of 7.1 for the third time then caused the RP to become significantly more negative (-53.00±3.27 mV, p = 0.034). Thus, extracellular alkalization is shown to hyperpolarize the RP whereas acidification causes depolarization. It is important to note that the final RP in saline at pH 7.1 was still significantly depolarized compared to the initial RP value at pH 7.1 (p = 0.025). The Fig. 8d depicts the trends in percent change of the RP between each condition.
These studies are important in developing a cellular model of the potential effects of intracellular and extracellular acidification on evoked and non-evoked synaptic transmission (Fig. 9). The aspects of the model are addressed in the discussion below.
DISCUSSION
There are several important findings from this study to be addressed. Propionic acid was shown to depolarize the membrane potential of the cell, with repolarization occurring following flushing with fresh saline. The extent of depolarization was found to be greater with a higher concentration of propionic acid, indicating a dose-dependent effect. Analysis of spontaneous quantal events before and after propionic acid administration indicates that, at a 20 mM concentration, propionic acid can cause an increase in mEJP frequency which is decreased following washout. The inhibition of calcium channels with CdCl2 prevents the increase in mEJP frequency during the exposure to propionic acid. At 40 mM propionic acid, the resultant frequency of mEJPs was so high as to prevent accurate analysis; while the data could not be quantified, there was a notable increase in mEJP frequency in all preparations observed. This effect was also reversed upon removal of propionic acid. These findings support the earlier report by Caldwell et al.9 with measures in the frequency of mEJPs.
The time to 50% EJP amplitude depression provided insight into the presynaptic effects of pH through alterations in vesicular packaging and fusion. Due to the graded nature of EJPs at the Drosophila NMJ, EJP amplitude is dependent upon the amount of neurotransmitter released from the presynaptic cell to act at the glutamatergic receptors at the postsynaptic membrane. The 5 Hz stimulation was applied to the preparations to simulate the high activity of synaptic function, which over time leads to depletion of vesicles available in the readily releasable pool (RRP) and recruitment of vesicles from the recycling and reserve pools. It is also important to note the effect of resting membrane potential (RP) changes in the postsynaptic cell for this experimental paradigm. As RP changes, the driving gradient of ions is altered. This altered driving gradient itself can affect the amplitude of EJPs independent of the effects involving vesicular dynamics at the presynaptic membrane. Compared to the saline conditions, 20 mM propionic acid application led to extremely rapid rundown of EJP amplitude. This was shown for both the voltage-clamped and non-voltage-clamped preparations. This indicates that low intracellular pH likely has some effect on vesicular dynamics, such that vesicles are unable to be repackaged or recycled and high stimulation rapidly depletes the vesicle pools available.
As a second mechanism of intracellular acidification, a pre-pulse of ammonium chloride and rapid removal with washout was used. Application of ammonium chloride at both 20 and 40 mM concentrations was shown to depolarize the cell, whereas its rapid removal causing transient acidification led to repolarization towards the initial RP. Thus, both alkalization by ammonium chloride and acidification with propionic acid were associated with membrane depolarization, but acidification after ammonium chloride washout was associated with repolarization. This may be explained by the transient nature of the acidification; it is possible that restoration to pH equilibrium occurred so rapidly as to not affect the RP.
The effects of ammonium chloride on spontaneous quantal events are more complex. At the lower concentration, mEJP frequency is only decreased after application of ammonium chloride. However, a second application and washout had no significant effects on mEJP frequency. At the 40 mM concentration, the first application of ammonium chloride had no effect, but washout led to a reduction in mEJP frequency. Interestingly, however, in comparison to the lower concentration, the second application of ammonium chloride showed a non-significant trend towards an increase in mEJP frequency. A second washout was able to reverse this effect. This indicates that the direction of the effect of pHi through this method of acidification also differs based on the concentration utilized. A possible explanation for this could be that, in the case of 40 mM ammonium chloride application, a second exposure may affect the intracellular environment had not returned to a normal status in regards to the effects on channels and/or protein structure. The second dose may then act as somewhat of an additive effect, although the RP was able to be recovered before the second application of ammonium chloride. There may be a bell curve effect on proteins and vesicle dynamics by ammonium chloride at different concentrations; it has been shown previously that both 5 mM and 50 mM ammonium chloride application in rat hippocampal cultures inhibits vesicle release15. Ammonium chloride was also shown to elevate intracellular calcium levels, potentially due to blocking of mitochondrial calcium uptake; these effects were long-lasting and independent of the alterations in pH and membrane potential15.
A difference from previous studies9,16,17 is the use and analysis of a second bout of acidification within each preparation. Such a method can further elucidate the potential compounding effects of acidification and the reversibility of the effects with multiple applications. It also provides information and insight into how preparations respond differently to the first compared to the second administration as well as how these patterns may differ based on the concentration of the compound used. While acidification via propionic acid leads to a consistent pattern of RP depolarization and hyperpolarization along with mEJP frequency increase and reduction following washout at both 20 and 40 mM concentrations, ammonium chloride seems to differ in its responses based on concentration as well as whether it is the first or second application. This may indicate potential issues that arise in the use of ammonium chloride for such an assay.
Repetitive respiratory acidosis occurs with humans with sleep apneas18,19 as well as repetitive metabolic acidosis with some sporting events20. There have yet to be neurological examinations in reflexes or NMJ transmission reported in such cases, but such incidences could lead to synaptic impairments and health risks.
Altering extracellular pH rapidly with HCl or NaOH is sufficient to quickly change the membrane potential without drastic alterations in intracellular p H. Thus, when using PA or NH4Cl, the effect may initially be due to the extracellular effect followed by an intracellular action on spontaneous and evoked EJPs. The effects of both intracellular and extracellular alteration in pH on the RP of the cell may indicate pH sensitivity of sodium and potassium leak channels or potentially the sodium-potassium ATPase pump. Both intracellular and extracellular pH has been shown to alter the activity of the sodium-potassium ATPase pump21-24. In the squid axon, it was found that the pump functions maximally at a pH between 7.2 and 7.4, whereas both acidification and alkalization led to a reduction in pump activity21. This was also seen in rabbit urinary bladder cells22 and sheep erythrocytes24. Furthermore, intracellular acidification has been shown to increase potassium selectivity on the cytoplasmic side of the pump, potentially increasing competition for sodium binding25. Possibly, intracellular acidification through propionic acid administration or removal of ammonium chloride inhibits the function of the pump enough to cause depolarization as the ionic gradients of sodium and potassium are slowly lost. Additionally, pH may affect sodium and potassium leak channels themselves that are crucial for the maintenance of the RP. In rat hepatocytes, it was found that intracellular acidification led to depolarization of the RP, a result attributed to the accompanied reduction in potassium flux observed26. Another effect to consider is the status of the NHE that is thought to be involved in the modulation of intracellular pH. At acidic intracellular pH, the NHE is implicated in restoring intracellular pH as it pumps out protons in exchange for extracellular sodium27,28. The influx of sodium may affect the RP by altering the ionic gradient across the membrane. In Drosophila, it has been shown that mEJP frequency increase upon addition of propionic acid is enhanced by the use of the NHE blocker EIPA, demonstrating that slowing the restoration of intracellular pH to equilibrium prolongs the effects of acidification9. Other methods of pH regulation in the cell may also be at play, buffers and other transporters such as those for bicarbonate can also help restore pH after perturbation from equilibrium. The complete mechanisms that the cell employs to sense and restore pH alterations are highly complex and not yet fully identified.
Of course, many other proteins may be affected by the drop in pH such as the sodium-calcium exchanger (NCX), calcium pumps, or the Na+/K+ ATPase pump, it has not yet been fully investigated how extracellular pH as well as intracellular pH, fully affects these proteins and the complete interactions involving all of their individual effects. The NCX is inhibited during both extracellular acidification and alkalization29 and inhibited during intracellular acidification30. The large number of proteins involved in Ca2+ sensing, vesicle docking and vesicle fusion have yet to be investigated for the interaction of various intracellular pH conditions that are associated with physiological or pathological neuronal activity. The lack of research in addressing the effect of pH on the SNARE-SNAP protein interaction is an area ripe for further studies.
Given that the frequency of spontaneous vesicular fusion increases upon acidification of the intracellular compartments of the presynaptic neuron using propionic acid, this suggests a possible increase in calcium entry and possibly enhanced SNARE-SNAP or even Ca2+ sensitivity of synaptotagmin. In the preparations involving the use of CdCl2, a calcium channel blocker, the effect of 20 mM propionic acid on mEJP frequency was decreased. Evoked synaptic transmission is depressed very rapidly under intracellular acidic conditions with propionic acid. This may be due to the depletion of readily releasable vesicles as a result of the high rate of spontaneous fusion events.
Both ammonia and propionate can increase in the blood of humans and other mammals under pathological conditions and rare diseases31. Thus, the use of ammonium chloride and propionic acid to experimentally manipulate pHi offers a useful approach to address the effects of pHi on cellular function in addition to the use of CO2. However, the effects of CO2 may have additional actions besides altering extracellular and intracellular pH and can be preparation-dependent. It was reported that the presence of CO2 blocks glutamatergic synapses at the larval Drosophila NMJ and crayfish NMJs independent of the decrease in pH32,33. While Caldwell et al.9 found that the application of 24 mM HCO3–/5% CO2 did not affect mEJP frequency, this disparity may be in part explained by the lower concentration used compared to the previous studies. The direct effect of CO2 on glutamatergic synapses in mammalian CNS preparations remains to be investigated.
This study served to address and expand upon findings from previous research in the study of pHi effects during acidification of the presynaptic nerve terminal. While many of the results corroborate previous findings, there are some differences to be addressed. In the study by Caldwell et al.9 that examined the effects of both propionic acid and the ammonium chloride pre-pulse technique on synaptic transmission, they found that propionic acid administration led to hyperpolarization of the RP, contrary to what was found in the current study. This can be partially explained by the use of the saline buffer BES. The study herein used a concentration of 25 mM whereas Caldwell et al.9 used 15 mM BES. Additionally, the RP was more negative for the preparations in the current study compared to those in Caldwell et al.9. Caldwell et al.9 incubated in ammonium chloride for 10 min compared to the 20 min used in the current study and there is no report on the timings of experimental dissection and the time between dissection and recording. Due to the rundown and degradation of preparations, it is possible that differences in such timings can affect results. Another point to consider is that they recorded from both muscle 4 and muscle 6 in their analysis of mEJP frequencies, with no distinction or separation in the results; the current study only analyzed mEJP frequencies from muscle 6. It is possible that differences in the muscles used caused disparities in the results obtained. Previous studies in Drosophila9 did not address the effect of acidification on evoked synaptic transmission. This is relevant in the case of modeling the effects of acidification during high neural activity, such as in pathological conditions involving seizures or slightly enhanced neural activity. The finding that acidification greatly decreases the time to synaptic depression indicates alteration of vesicular dynamics. This is in line with the findings that propionic acid administration increases mEJP frequency as well; if acidification increases the rate of vesicular fusion and impedes vesicular recycling and repackaging, mEJPs would be expected to increase in frequency at low or no stimulation as fusion is increased whereas high stimulation would result in greater numbers of fusion events and rapid depletion of the vesicle pools. This is supported by the findings at the lizard NMJ showing that acidification with propionic acid inhibits vesicular recycling by blocking endocytosis16. However, it has been reported that the Drosophila form of excitatory amino acid transporter (i.e., dEAAT1), is expressed in the larval brain and adult NMJ; it is not expressed in the larval periphery34.
While this study addresses the acute effects of altered pH on synaptic transmission, there are likely long-term effects as well. While this study has contributed to the development of a working model for the effects of pH on synaptic transmission, there are many further avenues of experimentation to address. These include a more in-depth analysis of pH effects for presynaptic function, such as through effects on vesicle loading, repackaging, recycling, docking and fusion and sensitivity to Ca2+ binding to synaptotagmin. There are also potential effects on voltage sensitivity of ion channels with evoked events, Ca2+ buffering, the NCX, calcium pumps, ATP production related to mitochondrial function and the potential interaction of autoreceptors (see model in Fig. 9 and further details35.
CONCLUSION
In this study, propionic acid was shown to increase spontaneous quantal event frequency, reduce evoked transmission, enhance high-frequency depression and depolarize the target cell. The enhanced frequency of spontaneous events was attenuated with CdCl2 in the bath, indicating that calcium influx is involved in the resultant effects. The effects of ammonium chloride withdrawal were shown to be dose and condition-dependent. It was also shown that extracellular acidification depolarizes whereas alkalization hyperpolarizes the synaptic target. These studies are important in further developing a model of the effects of intracellular and extracellular acidification on evoked and non-evoked synaptic transmission.
SIGNIFICANCE STATEMENT
This study discovered and expanded upon previous findings for the effects of intracellular pH alteration on both spontaneous and evoked synaptic transmission in Drosophila melanogaster. These findings may be translationally important for uncovering the impact of such effects in cases where humans encounter transient pH alterations, such as in several disease states. Thus, this study contributes to the model of pH effects on synaptic transmission.
ACKNOWLEDGMENT
We thank Dr. Sandra L. E. Blümich (Veterinärmedizinische Fakultät, Universität Leipzig, Leipzig, Germany) for initial research on this project in our lab during the summer of 2014 while on a fellowship by Deutscher Akademischer Austausch Dienst (DAAD) German Academic Exchange Service (RISE-Program (Research Internships in Science and Engineering). Chellgren Endowed Professor fund from the University of Kentucky to R.L.C. for supplies.
REFERENCES
- Boron, W.F., 2004. Regulation of intracellular pH. Adv. Physiol. Educ., 28: 160-179.
CrossRefDirect Link - Casey, J.R., S. Grinstein and J. Orlowski, 2010. Sensors and regulators of intracellular pH. Nat. Rev. Mol. Cell Biol., 11: 50-61.
CrossRefDirect Link - Willoughby, D. and C.J. Schwiening, 2002. Electrically evoked dendritic pH transients in rat cerebellar Purkinje cells. J. Physiol., 544: 487-499.
CrossRefDirect Link - Rossano, A.J., A.K. Chouhan and G.T. Macleod, 2013. Genetically encoded pH-indicators reveal activity-dependent cytosolic acidification of Drosophila motor nerve termini in vivo. J. Physiol., 591: 1691-1706.
CrossRefDirect Link - Kuromi, H. and Y. Kidokoro, 2004. Two distinct pools of synaptic vesicles in single presynaptic boutons in a temperature-sensitive Drosophila Mutant, shibire Neuron, 20: 917-925.
CrossRefDirect Link - Ugur, B., K. Chen and H.J. Bellen, 2016. Drosophila tools and assays for the study of human diseases. Dis. Model. Mech., 9: 235-244.
CrossRefDirect Link - Cooper, R.L., B.A. Stewart, J.M. Wojtowicz, S. Wang and H.L. Atwood, 1995. Quantal measurement and analysis methods compared for crayfish and Drosophila neuromuscular junctions and rat hippocampus. J. Neurosci. Methods, 61: 67-78.
CrossRefDirect Link - Kurdyak, P., H.L. Atwood, B.A. Stewart and C.F. Wu, 1994. Differential physiology and morphology of motor axons to ventral longitudinal muscles in larval Drosophila. J. Compa. Neurol., 350: 463-472.
CrossRefDirect Link - Caldwell, L., P. Harries, S. Sydlik and C.J. Schwiening, 2013. Presynaptic pH and vesicle fusion in Drosophila larvae neurones. Synapse, 67: 729-740.
CrossRefDirect Link - De Castro, C., J. Titlow, Z.R. Majeed and R.L. Cooper, 2014. Analysis of various physiological salines for heart rate, CNS function and synaptic transmission at neuromuscular junctions in Drosophila melanogaster larvae. J. Comp. Physiol. A, 200: 83-92.
CrossRefDirect Link - Stewart, B.A., H.L. Atwood, J.J. Renger, J. Wang and C.F. Wu, 1994. Improved stability of Drosophila larval neuromuscular preparations in haemolymph-like physiological solutions. J. Comp. Physiol. A., 175: 179-191.
CrossRefDirect Link - Lee, J.Y., D. Bhatt, D. Bhatt, W.Y. Chung and R.L. Cooper, 2009. Furthering pharmacological and physiological assessment of the glutamatergic receptors at the Drosophila neuromuscular junction. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol., 150: 546-557.
CrossRef - Kim, S.M., V. Kumar, Y.Q. Lin, S. Karunanithi and M. Ramaswami, 2009. Fos and Jun potentiate individual release sites and mobilize the reserve synaptic-vesicle pool at the Drosophila larval motor synapse. PNAS., 106: 4000-4005.
CrossRefDirect Link - McLachlan, E.M. and A.R. Martin, 2014. Non-linear summation of end-plate potentials in the frog and mouse. J. Physiol., 311: 307-324.
CrossRefDirect Link - Lazarenko, R.M., C.E. DelBove, C.E. Strothman and Q. Zhang, 2017. Ammonium chloride alters neuronal excitability and synaptic vesicle release. Sci. Rep.
CrossRefDirect Link - Lindgren, C.A., D.G. Emery and P.G. Haydon, 2018. Intracellular acidification reversibly reduces endocytosis at the neuromuscular junction. J. Neurosci., 17: 3074-3084.
CrossRefDirect Link - Sandstrom, D.J., 2011. Extracellular protons reduce quantal content and prolong synaptic currents at the Drosophila larval neuromuscular junction. J. Neurogenet., 25: 104-114.
CrossRefDirect Link - Baekey, D.M., P. Feng, M. J. Decker and K.P. Strohl, 2013. Breathing and sleep: Measurement methods, genetic influences, and developmental Impacts. ILAR J., 50: 248-261.
CrossRefDirect Link - Popescu, N.A., M.D. Ionescu, G. Balan, S. Visan and E. Cinteza et al., 2018. Central sleep apnea – A rare cause for acute respiratory insufficiency in children. Case Report. MAEDICA (Buchar), 13: 66-70.
Direct Link - Knechtle, B., M. Stjepanovic, C. Knechtle, T. Rosemann, C.V. Sousa and P.T. Nikolaidis, 2018. Physiological responses to swimming repetitive “Ice Miles”. J. Strength Conditioning Res.
CrossRefDirect Link - Breitwieser, G.E., A.A. Altamirano and J.M. Russell, 2017. Effects of pH changes on sodium pump fluxes in squid giant axon. Am. J. Physiology-Cell Physiol., 253: C547-C554.
CrossRefDirect Link - Eaton, D.C., K.L. Hamilton and K.E. Johnson, 2017. Intracellular acidosis blocks the basolateral Na-K pump in rabbit urinary bladder. Am. J. Physiology-Renal Physiol., 247: F946-F954.
CrossRefDirect Link - Salonikidis, P.S., S.N. Kirichenko, L.V. Tatjanenko, W.Schwarz and L.A. Vasilets, 2002. Extracellular pH modulates kinetics of the Na+, K+-ATPase. Biochim. Biophys. Acta (BBA)-Biomembranes, 1509: 496-504.
CrossRefDirect Link - Zade-Oppen, A.M.M., J.M. Schooler, P. Cook and D.C. Tosteson, 2003. Effect of membrane potential and internal pH on active sodium-potassium transport and on ATP content in high-potassium sheep erythrocytes. Biochim. Biophys. Acta (BBA) - Biomembranes, 555: 285-298.
CrossRefDirect Link - Cornelius, F., N. Tsunekawa and C. Toyoshima, 2017. Distinct pH dependencies of Na+/K+ selectivity at the two faces of Na, K-ATPase. J. Biol. Chem., 293: 2195-2205.
CrossRefDirect Link - Fitz, J.G., T.E. Trouillot and B.F. Scharschmidt, 2017. Effect of pH on membrane potential and K+ conductance in cultured rat hepatocytes. Am. J. Physiol.-Gastrointestinal and Liver Physiology, 257: G961-G968.
CrossRefDirect Link - Cha, C.Y., C. Oka, Y.E. Earm, S. Wakabayashi and A. Noma, 2009. A Model of Na+/H+ Exchanger and Its Central Role in Regulation of pH and Na+ in Cardiac Myocytes. Biophys. J., 97: 2674-2683.
CrossRefDirect Link - Koliakos, G., K. Paletas and M. Kaloyianni, 2008. NHE-1: A molecular target for signalling and cell matrix interactions. Connect. Tissue Res., 49: 157-161.
CrossRefDirect Link - Egger, M. and E. Niggli, 2004. Paradoxical block of the Na+‐Ca2+ exchanger by extracellular protons in guinea‐pig ventricular myocytes. J. Physiol., 523: 353-366.
CrossRefDirect Link - Doering, A.E., D.A. Eisner and W.J. Lederer, 2006. Cardiac Na-Ca exchange and pH. Ann. N. Y. Acad. Sci., 779: 182-198.
CrossRefDirect Link - Jafari, P., O. Braissant, P. Zavadakova, H. Henry, L. Bonafé and D. Ballhausen, 2013. Brain damage in methylmalonic aciduria: 2-methylcitrate induces cerebral ammonium accumulation and apoptosis in 3D organotypic brain cell cultures. Orphanet J. Rare Dis., 8:4.
CrossRefDirect Link - Badre, N.H., M.E. Martin and R.L. Cooper, 2005. The physiological and behavioral effects of carbon dioxide on Drosophila melanogaster larvae. Comp. Biochem. Physiol. A Mol. Integr. Physiol., 140: 363-376.
CrossRef - Bierbower, S.M. and R.L. Cooper, 2013. The mechanistic action of carbon dioxide on a neural circuit and NMJ communication. J. Exp. Zool., 319: 340-354.
CrossRef - Rival, T., L. Soustelle, D. Cattaert, C. Strambi, M. Iché and S. Birman, 2006. Physiological requirement for the glutamate transporter dEAAT1 at the adult Drosophila neuromuscular junction. J. Neurobiol., 66: 1061-1074.
CrossRefDirect Link