
Min Chen
Institute of Natural Medicinal Chemistry and Green Chemistry, College of Light Industry and Chemical Engineering, Guangdong University of Technology, Guangzhou 510006, Guangdong, China
Shun Huang
Nanfang PET Center, Nanfang Hospital, Southern Medical University, Guangzhou, 510515 Guangdong, China
Yuan-Yuan Chang
Institute of Natural Medicinal Chemistry and Green Chemistry, College of Light Industry and Chemical Engineering, Guangdong University of Technology, Guangzhou 510006, Guangdong, China
Xi Zheng
Institute of Natural Medicinal Chemistry and Green Chemistry, College of Light Industry and Chemical Engineering, Guangdong University of Technology, Guangzhou 510006, Guangdong, China
Ren-Ping Zhou
Susan Lehman Cullman Laboratory for Cancer Research, Department of Chemical Biology, Ernest Mario School of Pharmacy, Rutgers, The State University of New Jersey, 08854 Piscataway, New Jersey, USA
Dong-Li Li
Wuyi University, Jiangmen 529020, Guangdong, China
Kun Zhang
International Healthcare Innovation Institute (Jiangmen), 529000 Jiangmen, China
LiveDNA: 86.23260
Current Research in Neuroscience
Year: 2018 | Volume: 8 | Issue: 1 | Page No.: 1-9
DOI: 10.3923/crn.2018.1.9







ABSTRACT
Background and Objective: Curcumin is a bioactive natural polyphenol, but the poor bioavailability has limited its application. To be effective as a therapeutic drug, curcumin must be combined with other drugs to improve bioavailability. The aim of the present study was to investigate the effects and mechanisms of curcumin and piperine administered individually or in combination on lipopolysaccharide (LPS)-induced neuroinflammation. Materials and Methods: Mice were treated with curcumin (100 and 200 mg kg–1, p.o.) and piperine (20 mg kg–1, p.o.) for 21 days followed by LPS (250 μg kg–1, i.p.) administration for 7 days. Results: It is found that repeat injections of LPS induced neuronal damage and increased inflammatory cytokines release, while pretreatment with curcumin reversed these changes. In addition, co-administration of piperine with curcumin potentiated their neuroprotective effects as compared to the treatment with curcumin alone. It is further examined that the molecular mechanisms by which curcumin exerts its neuroprotective effects and inhibited the Toll-like receptor 4 (TLR4)-mediated downstream signaling and effectively lowered the production of inflammatory mediators. Conclusion: These findings led us to the conclusion that the effects of curcumin on LPS-induced neuroinflammation is mediated by modulating TLR4 pathway, which is enhanced by an adjuvant, piperine.
PDF Abstract XML References Citation
How to cite this article
Min Chen, Shun Huang, Yuan-Yuan Chang, Xi Zheng, Ren-Ping Zhou, Dong-Li Li and Kun Zhang, 2018. Peperine Enhancement on Neuroinflammatory Effects of Curcumin and its Mediation via Modulating Toll like Receptor-4 Pathway. Current Research in Neuroscience, 8: 1-9.
DOI: 10.3923/crn.2018.1.9
URL: https://scialert.net/abstract/?doi=crn.2018.1.9
DOI: 10.3923/crn.2018.1.9
URL: https://scialert.net/abstract/?doi=crn.2018.1.9
INTRODUCTION
Neuroinflammation is the most common hallmark in many neurodegenerative diseases, including Alzheimer’s disease, Parkinson’s disease, amyotrophic lateral sclerosis and multiple sclerosis. Although neuroinflammation may not be an initiating factor in neurodegenerative diseases, there is emerging evidence, which indicates that it acts as an amplifier to exacerbate diseases progression. Under inflammatory conditions, microglia and astrocytes are activated extensively, promoting the release of cytokines, reactive oxygen species (ROS) and nitric oxide (NO), that further lead to the increase of neurotoxicity and neuronal death1. Since neuroinflammation usually starts from the beginning of neurodegeneration, it is likely that anti-inflammatory intervention will be long-term, imposing a critical need for a high level safety2. Recent studies suggest that anti-neuroinflammation agents from traditional herbs may be safe and effective3,4.
Curcumin is a bioactive natural polyphenol, which is widely used as herbal medicine in Asian countries5. It has been shown to have beneficial effects on various chronic conditions, including autoimmune, cardiovascular, neurodegenerative and psychological diseases, diabetes and cancer6,7. Furthermore, curcumin is safe and well-tolerated even at high dose up to 8 g/day. However, the major limitation of curcumin is poor bioavailability, which embodies in its low solubility, poor absorption, rapid metabolism and elimination8. Therefore, strategies to promote curcumin bioavailability are highly desirable. Piperine is a natural alkaloid in black pepper, which inhibits the hepatic and intestinal glucuronidation and enhances the bioavailability of many drugs9. In the present study, we employed a classical LPS model to evaluate the possible neuroprotective effects of curcumin alone and in combination with piperine and investigated its mechanisms of action.
MATERIALS AND METHODS
This study was performed from February, 2016 to December, 2016 in the laboratories of the Institute of Natural Medicinal Chemistry and Green Chemistry in Guangdong University of Technology, Guangzhou, China.
Materials: Curcumin and piperine were obtained from Sinopharm Chemical Reagent Company. The LPS (serotype O55:B5) was purchased from Sigma-Aldrich. Antibodies against TLR4, MAPK, Myd88, IL-1β and β-actin were purchased from Abcam (Cambridge, UK). Antibodies against NF-κB p65 and TNF-α were obtained from Cell Signaling Technology Inc (Boston, USA).
Animals and treatment: The C57 mice (4-5 weeks) were purchased from Animal Center of Sun Yat-Sen University (Guangzhou, China). All mice were housed in a pathogen-free animal room under controlled temperature (23±2°C) and humidity (60±5%) with a 12/12 h light/dark cycle and provided with standard laboratory diet and water. Mice were acclimatized to the facility for 4 weeks before experimental procedures were initiated. All animal care and experimental procedures were approved by the Laboratory Animal Ethics Committee of Guangdong University of Technology (SCXK2011-0029). All studies were conducted on the basis of the guidelines set by the National Institute of Health (the 7th edition, USA).
Animals were randomly divided into 7 groups with 6 mice in each group: Control, LPS, Cur-L (LPS+100 mg kg–1/day curcumin), Cur-H (LPS+200 mg kg–1/day curcumin), Pip (LPS+20 mg kg–1/day piperine), Cur-L+Pip (LPS+100 mg kg–1/day curcumin+20 mg kg–1/day piperine) and Cur-H+Pip (LPS+200 mg kg–1/day curcumin+ 20 mg kg–1/day piperine). Mice were intraperitoneal (i.p.) administered with LPS (250 μg kg–1) once daily for 7 days after the drugs treatment. Control mice were injected with PBS. Curcumin and piperine were orally administered to mice for 21 days (once a day) while other groups received PBS only.
Nissl staining and cell counting: For histologic analysis, tissue samples were washed twice with 0.01 M PBS and then fixed in 10% buffered formalin. Paraffin-embedded brain specimens were sectioned at 5 μm in a rotary microtome. Slices were stained with 0.5% cresyl violet solution for 10 min at 50°C, rinsed with distilled water and dehydrated through an alcohol series (70, 95 and 100%). Tissue slices were permeabilized with xylene and mounted with neutral resin. Neuronal density was evaluated by counting the number of normal neurons, that had prominent Nissl bodies and distinct nucleus, in three fields (400×magnification) per slide. Immunohistochemistry staining: In situ expression of TNF-α in the brain was examined using immunohistochemistry as follows. Sections were first treated with TBS+0.4% Triton+1% FBS for 1 h at room temperature. The samples were then incubated overnight at 4°C with primary anti-TNF-α antibody, washed twice with TBS+0.4% Triton for 5 min each at room temperature and followed by 2 h incubation at room temperature with horseradish peroxidase-conjugated secondary antibodies in TBS+0.4% Triton. The sections were then counterstained with hematoxylin for 2 min to stain cell nuclei. The stained sections were visualized using a microscope.
Cell culture: Immortalized murine microglial (BV2) cells were grown in DMEM (Gibco, California, USA) containing 10% FBS (Gibco, California, USA) in a humidified atmosphere containing 5% CO2. When 80% confluence was reached, cells were trypsinized and seeded into cell culture dishes (100×20 mm) at 2×105 cells mL–1 and grown to 70-80% confluence, cells were then pre-incubated for 1 h in the absence or presence of curcumin (2.5, 5 and 10 μM) before addition of LPS (100 ng mL–1) for 24 h. The conditioned medium was analyzed immediately to determine the levels of secreted TNF-α and IL-1β using Quantikine mouse TNF-α or IL-1β ELISA kits (CUSABIO, Wuhan, China).
Western blot analysis: Treated cells were lysed in lysis buffer (Beyotime Institute of Biotechnology, Shanghai, China) and protein contents were determined with Pierce BCA protein assay kit (Thermo Fisher Scientific, Maryland, USA). About 20 μg of protein lysates were subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to PVDF membranes. The membranes were blocked in 5% BSA in TBST for 1.5 h and then incubated overnight at 4°C with primary anti-NF-κB p65, TNF-α, IL-1β, TLR4, MAPK and Myd88 antibodies. Bound primary antibody was detected by horseradish peroxidase conjugated secondary antibody followed by chemiluminescence detection (MultiSciences Biotech, Hangzhou, China). The bands density was quantified using the Quantity One analysis software (Bio-Rad, California, USA) via calculating the average optical density in each field.
Short interfering RNA (siRNA) transfection: The TLR4 siRNAs were purchased from Ribo Biotechnology (Ribo Bio, Guangzhou, China). The TLR4 siRNAs were transfected into BV2 cells according to a siRNA transfection protocol provided by Ribo Biotechnology. Briefly, after culturing BV2 cells in antibiotic-free DMEM at 37°C in a humidified atmosphere of 5% CO2 for 24 h, the siRNA/ lipofectamine 2000 (Invitrogen, Carlsbad, California, USA) complexes, which were diluted in OPTIMEM medium (Gibco, California, USA), were added to the BV2 cells. After transfection, the BV2 cells were subjected to LPS stimulation for 24 h and the conditioned medium was analyzed immediately to determine the levels of secreted TNF-α and IL-1β using Quantikine mouse TNF-α or IL-1β ELISA kits (CUSABIO, Wuhan, China). A non-targeting siRNA (si-NT, Ribo Bio) was used as negative control. Statistical Analysis: The data are presented as means±standard deviation (SD). Statistical comparisons of the data were performed using the one-way ANOVA of the SPSS 19.0 system for multiple comparisons with Tukey post-hoc test to determine statistical significance. Differences were considered as statistically significant at p<0.05.
RESULTS
Effects of curcumin and piperine alone or in combination on LPS-elicited neuronal damage: Systemic LPS administration has been known to cause chronic neuroinflammation, which has been implicated in neuronal damage10. The neuroprotective effect of curcumin was investigated morphologically using Nissl staining in the cortex and hippocampus of adult mice. As shown in Fig. 1, a clear reduction in Nissl substance was observed in LPS-treated mice compared to the control group (LPS vs. Control: p<0.01). Curcumin increased the number of Nissl bodies and neurons (In the hippocampus: Cur-H vs. LPS: p<0.05; in the cortex: Cur-L vs. LPS: p<0.05.). The number of normal neurons was significantly increased in the combination group (In the cortex: Cur-L+Pip vs. Cur-L: p<0.05), although piperine alone did not produce any distinct effect on histopathologic changes.
Effects of curcumin and piperine alone or in combination on LPS-induced TNF-α and IL-1β expressions: Robust cytokine production and excessive inflammatory infiltrates are the major factors of neuronal damage after LPS injection11. In the present study, the expression levels of inflammatory cytokines were determined by immunohistochemistry and ELISA analysis. As shown in Fig. 2, a significant increase in the protein levels of TNF-α and IL-1β upon LPS injections was observed when compared with the control group (LPS vs. Control: p<0.01).
Curcumin effectively lowered TNF-α and IL-1β protein levels (In the hippocampus: The Cur-H vs. LPS: p<0.05; in the cortex: Cur-L vs. LPS: p<0.05 and Cur-H vs. LPS: p<0.05.). In addition, co-administration of curcumin with piperine significantly potentiated the anti-inflammatory effect as compared to the treatment alone (In the hippocampus: Cur-L+Pip vs. Cur-L: p<0.05, in the cortex: Cur-L+Pip vs. Cur-L: p<0.05 and Cur-H+Pip vs. Cur-H: p<0.05.). However, piperine alone did not produce any significant effect on the release of inflammatory cytokines as compared to LPS group. These observations indicate that curcumin is the main biologically active compound in the combination treatment and piperine significantly increased the curcumin anti-inflammatory effects.
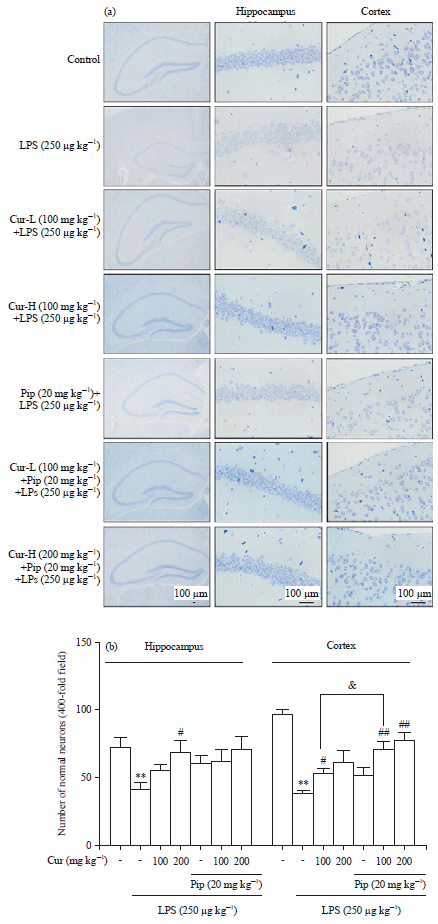 | |
| Fig. 1(a-b): | Effects of curcumin and piperine alone or in combination on LPS-elicited neuronal damage. Nissl staining and normal neuronal counts in hippocampus and cortex of each group, scale bar = 100 μm |
| Data represent the Mean±SD (n = 6 mice per group). The significance of differences from control group is at **p<0.01, from LPS group at #p<0.05 and ##p<0.01, from Cur-L group at &p<0.05. Cur: Curcumin, Pip: Piperine | |
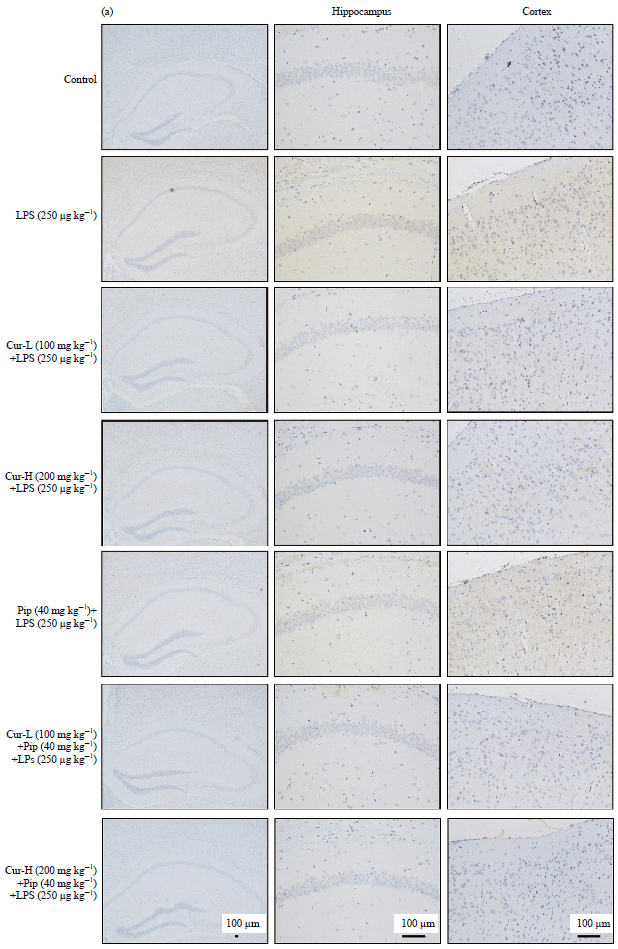 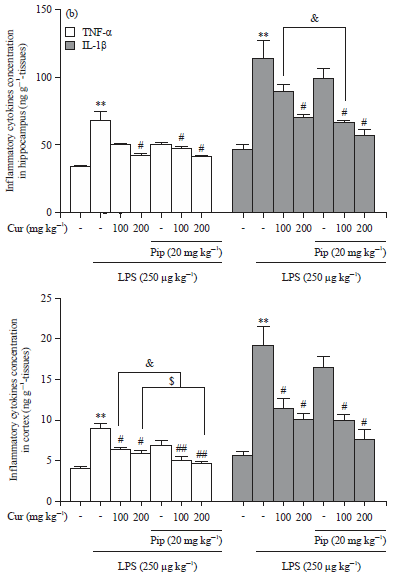 | |
| Fig. 2(a-b): | Effects of curcumin, piperine alone or in combination on LPS-induced TNF-α and IL-1β expressions, (a) Immunostaining of TNF-α in cortex and hippocampus, scale bar = 100 μm and (b) ELISA was conducted to determine the TNF-α and IL-1β levels in the brain homogenates of mice |
| Data represent the Mean±SD (n = 6 mice per group). The significance of differences from control group is at **p<0.01, from LPS group at #p<0.05 and ##p<0.01, from Cur-L group at &p<0.05, from Cur-H group at $p<0.05. Cur: Curcumin, Pip: Piperine | |
Anti-neuroinflammatory mechanism of curcumin against LPS-induced neuroinflammation in BV2 cells: Since the in vivo data have suggested a major role of curcumin in the combination, we subsequently examined the molecular mechanism of curcumin in vitro using cultured BV2 microglial cells. As shown in Fig. 3a, treatment with curcumin within 20 μM for 24 h showed no evident effect on the cell viability. However, the viability of BV2 cells treated with LPS showed a significant, dose-dependent decline (p<0.01). Importantly, curcumin (2.5, 5 and 10 μM) effectively prevented cytotoxicity of LPS of BV2 cells (p<0.01).
The LPS activates microglia to release proinflammatory mediators, which may further damage neuronal cells, stimulate toxic substances synthesis and increase microglial activation through a positive feedback loop12. Thus, we examined whether curcumin inhibited the production of proinflammatory mediators induced by LPS in cultured BV2 microglia cells. As shown in Fig. 3b, both TNF-α and IL-1β expressions in LPS-stimulated BV2 cells were decreased by curcumin.
The TLR4 is one of the cell surface receptors, which mediates LPS effects. Activated TLR4/MD-2 complex recruits Myd88 to activate MAPK and NF-κB which subsequently induce the production of inflammatory cytokines. As shown in Fig. 3c, curcumin significantly decreased the expressions of Myd88, MAPK and NF-κB induced by LPS, while treatment with curcumin showed no evident effect on the expression of TLR4.
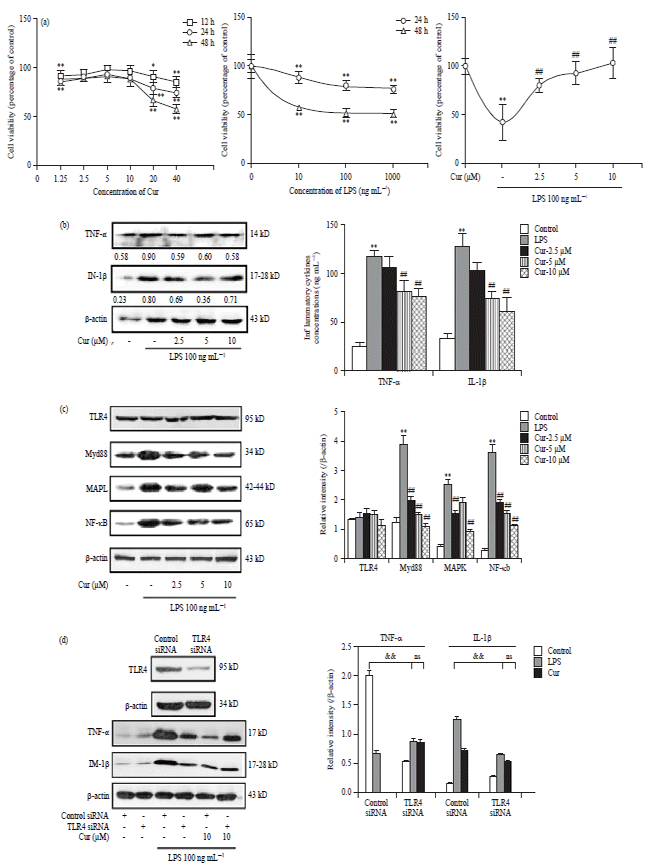 | |
| Fig. 3(a-d): | Anti-neuroinflammatory mechanism of curcumin against LPS-induced neuroinflammation in BV2 cells, (a) Curcumin prevents the cytotoxicity of LPS in BV2 cells, (b) Western blotting and ELISA measurement of TNF-α and IL-1β in BV2 cells treated with LPS (100 ng mL–1) for 24 h with or without curcumin pretreatment, (c) Curcumin regulates TLR4 downstream Signaling pathway. TLR4, Myd88, MAPK and NF-κB protein expressions were determined by western blotting and normalized by β-actin and (d) Effect of curcumin on TLR4-mediated TNF-α and IL-1β production |
| The significance of differences from control group is at *p<0.05 and **p<0.01, from LPS group at #p<0.05 and ##p<0.01, from control siRNA group at &&p<0.01, ns: Not significant, Cur: Curcumin | |
To examine whether TLR4 mediates curcumin effects, TLR4 protein expression was down-regulated using a siRNA against the receptor. The TLR4 protein level was significantly reduced in BV2 cells after transfection with TLR4 siRNA. As expected, down-regulation of TLR4 resulted in a clear reduction in TNF-α and IL-1β production (p<0.01, p<0.01). In addition, the levels of TNF-α and IL-1β were not decreased in TLR4 siRNA transfected cells (ns, ns), which had been pretreated with curcumin (Fig. 3d), indicating that curcumin inhibits TLR4-mediated cytokines production.
DISCUSSION
In accordance with our previous report13, cognitive impairment and abnormal cerebral metabolism, two markers of brain injury, were observed in mice after repeated LPS injections. Meanwhile, pathological damage, including extensive loss of Nissl substance and normal neuronal cells, was also observed in the brain of LPS-treated mice14,15. Neuroinflammation, the inflammation in the brain, has been postulated to play a key role in the mechanisms responsible for brain damage. In the present study, mice subjected to LPS (250 μg kg–1, i.p.) administration for 7 days showed a marked reduction in the number of normal neurons and a significant increase in inflammatory cytokines in cortex and hippocampus of adult mice (Fig. 1, 2), which supports previous reports that systemic LPS administration induces chronic neuroinflammation and triggers neuropathological change16,17.
Curcumin is one of the most studied phytochemical agents in the spice turmeric, which displays complex and multifaceted activities18. There have been many reports about curcumin and its roles in inflammation. However, its insolubility in water and poor bioavailability have limited its therapeutic applications. To be effective as a therapeutic drug, curcumin must be combined with other drugs to improve bioavailability8. For these reasons, we assessed the effects of using curcumin alone and in combination with piperine in LPS model of neuroinflammation. We found that curcumin treatment alleviated the LPS-induced neuropathological change as evidenced by elevated Nissl substance in cells and increased number of normal neurons. The effects were further enhanced when piperine was co-administered with curcumin. Neuroinflammation has been proposed to be a key contributor to neuronal damage19,20. Therefore, we assayed the changes of protein expression levels of inflammatory cytokines in this study. The results showed that curcumin treatment suppressed the inflammatory processes, reflected by decreased protein expressions of TNF-α and IL-1β. Moreover, co-administration of curcumin with piperine reduced the cytokines release more effectively as compared to curcumin treatment alone (Fig. 2), suggesting that the protective effect of curcumin was more pronounced when given in the combination with piperine.
These analysis of the anti-neuroinflammation mechanism of curcumin indicates that curcumin is able to inhibit the LPS-induced secretion of TNF-α and IL-1β. Several lines of evidence suggest that anti-inflammatory effect of curcumin occurs by the down-regulation of TNF-α and IL-1β secretion, via interfering with TLR4 mediated NF-κB and MAPK signaling pathways21. Since it is observed that no difference in TLR4 expression levels when pretreated with curcumin, we propose that curcumin exerts its effects on TLR4 downstream signals. Indeed, these results showed that curcumin suppressed LPS-induced TLR4 downstream signaling, including elevated protein expressions of Myd88, MAPK and NF-κB and the release of inflammatory mediators, such as TNF-α and IL-1β, which were consistent with previous studies22. In this study, we have gone further to confirm the role of TLR4-dependent signaling by investigating the effects of TLR4 knockdown. Curcumin treatment reduced TNF-α and IL-1β levels compared to vehicle-treated normal cells in the presence of LPS. In parallel, the TNF-α and IL-1β levels in TLR4 knockdown cells were not affected. Thus, we can infer that curcumin can protect neurons from LPS-induced inflammation via TLR4-mediated downstream signaling.
CONCLUSION
The present work suggested that curcumin could ameliorate LPS-induced brain injury through suppressing inflammatory response, which may be related to TLR4-mediated signaling. Further, piperine, as bioavailability enhancer, improved curcumin performance.
SIGNIFICANCE STATEMENTS
This study investigated the synergistic inhibitory effects of curcumin in combination with piperine on neuroinflammation mice model, which may be beneficial for development of inflammatory disease drug combinations. Furthermore, this study uncovered the molecular mechanism of curcumin by suppressing TLR4 downstream signaling and decreasing inflammatory mediators in BV2 cells, providing a therapeutic target for neuroinflammation-related diseases.
ACKNOWLEDGMENT
This study was supported by Department of Education of Guangdong Province (No. 2016KCXTD005).
REFERENCES
- Glass, C.K., K. Saijo, B. Winner, M.C. Marchetto and F.H. Gage, 2010. Mechanisms underlying inflammation in neurodegeneration. Cell, 140: 918-934.
CrossRefPubMedDirect Link - Eisen, A., M. Kiernan, H. Mitsumoto and M. Swash, 2014. Amyotrophic lateral sclerosis: A long preclinical period? J. Neurol. Neurosurg. Psychiatry, 85: 1232-1238.
CrossRefDirect Link - Spencer, J.P.E., K. Vafeiadou, R.J. Williams and D. Vauzour, 2012. Neuroinflammation: Modulation by flavonoids and mechanisms of action. Mol. Aspects Med., 33: 83-97.
CrossRefDirect Link - Bhullar, K.S. and H.P. Rupasinghe, 2013. Polyphenols: Multipotent therapeutic agents in neurodegenerative diseases. Oxid. Med. Cell. Longev., Vol. 2013.
CrossRefDirect Link - Goel, A., A.B. Kunnumakkara and B.B. Aggarwal, 2008. Curcumin as "Curecumin": From kitchen to clinic. Biochem. Pharmacol., 75: 787-809.
CrossRefPubMedDirect Link - He, Y., Y. Yue, X. Zheng, K. Zhang, S. Chen and Z. Du, 2015. Curcumin, inflammation and chronic diseases: How are they linked? Molecules, 20: 9183-9213.
CrossRefPubMedDirect Link - Wanninger, S., V. Lorenz, A. Subhan and F.T. Edelmann, 2015. Metal complexes of curcumin-synthetic strategies, structures and medicinal applications. Chem. Soc. Rev., 44: 4986-5002.
CrossRefPubMedDirect Link - Belkacemi, A., S. Doggui, L. Dao and C. Ramassamy, 2011. Challenges associated with curcumin therapy in Alzheimer disease. Expert Rev. Mol. Med., Vol. 13.
CrossRefDirect Link - Atal, C.K., R.K. Dubey and J. Singh, 1985. Biochemical basis of enhanced drug bioavailability by piperine: Evidence that piperine is a potent inhibitor of drug metabolism. J. Pharmacol. Exp. Ther., 232: 258-262.
PubMedDirect Link - Perry, V.H., C. Cunningham and C. Holmes, 2007. Systemic infections and inflammation affect chronic neurodegeneration. Nat. Rev. Immunol., 7: 161-167.
CrossRefDirect Link - Erickson, M.A. and W.A. Banks, 2011. Cytokine and chemokine responses in serum and brain after single and repeated injections of lipopolysaccharide: Multiplex quantification with path analysis. Brain Behav. Immun., 25: 1637-1648.
CrossRefPubMedDirect Link - Streit, W.J., R.E. Mrak and W.S.T. Griffen, 2004. Microglia and neuroinflammation: A pathological perspective. J. Neuroinflamm., Vol. 1.
CrossRefDirect Link - Chen, M., Y.Y. Chang, S. Huang, L.H. Xiao and W. Zhou et al., 2018. Aromatic‐turmerone attenuates LPS‐induced neuroinflammation and consequent memory impairment by targeting TLR4‐dependent signaling pathway. Mol. Nutr. Food Res., Vol. 62, No. 2.
CrossRefDirect Link - Badshah, H., T. Ali and M.O. Kim, 2016. Osmotin attenuates LPS-induced neuroinflammation and memory impairments via the TLR4/NFκB signaling pathway. Scient. Rep., Vol. 6.
CrossRefDirect Link - Choi, D.Y., M. Liu, R.L. Hunter, W.A. Cass and J.D. Pandya et al., 2009. Striatal neuroinflammation promotes Parkinsonism in rats. PLoS One, Vol. 4, No. 5.
CrossRefDirect Link - Semmler, A., S. Hermann, F. Mormann, M. Weberpals and S.A. Paxian et al., 2008. Sepsis causes neuroinflammation and concomitant decrease of cerebral metabolism. J. Neuroinflamm., Vol. 5.
CrossRefDirect Link - Qin, L., X. Wu, M.L. Block, Y. Liu and G.R. Breese et al., 2007. Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia, 55: 453-462.
CrossRefPubMedDirect Link - Gupta, S.C., B. Sung, J.H. Kim, S. Prasad, S.Y. Li and B.B. Aggarwal, 2013. Multitargeting by turmeric, the golden spice: From kitchen to clinic. Mol. Nutr. Food Res., 57: 1510-1528.
CrossRefDirect Link - Lee, J.W., Y.K. Lee, D.Y. Yuk, D.Y. Choi, S.B. Ban, K.W. Oh and J.T. Hong, 2008. Neuro-inflammation induced by lipopolysaccharide causes cognitive impairment through enhancement of beta-amyloid generation. J. Neuroinflammation, Vol. 5.
CrossRefDirect Link - Lee, Y.J., D.Y. Choi, Y.P. Yun, S.B. Han, K.W. Oh and J.T. Hong, 2013. Epigallocatechin-3-gallate prevents systemic inflammation-induced memory deficiency and amyloidogenesis via its anti-neuroinflammatory properties. J. Nutr. Biochem., 24: 298-310.
CrossRefPubMedDirect Link - Anthwal, A., B.K. Thakur, M.S.M. Rawat, D.S. Rawat, A.K. Tyagi and B.B. Aggarwal, 2014. Synthesis, characterization and in vitro anticancer activity of C-5 curcumin analogues with potential to inhibit TNF-α-induced NF-κB activation. BioMed Res. Int.
CrossRefDirect Link - Youn, H.S., S.I. Saitoh, K. Miyake and D.H. Hwang, 2006. Inhibition of homodimerization of Toll-like receptor 4 by curcumin. Biochem. Pharmacol., 72: 62-69.
CrossRefPubMedDirect Link