
Samuel Mburu Kamau
Dairy Training Institute, P.O. Box 449, Naivasha 20117, Kenya
Rong-Rong Lu
State Key Laboratory of Food Science and Technology, School of Food Science and Technology, Jiangnan University, 1800 Lihu Avenue, Wuxi, Jiangsu 214122, China
Current Research in Dairy Sciences
Year: 2011 | Volume: 3 | Issue: 1 | Page No.: 25-35







ABSTRACT
The Degree of Hydrolysis (DH) and the scavenging activity of DPPH radical of Whey Protein Hydrolysates (WPH) were investigated. The untreated commercial whey protein concentrate was hydrolyzed with trypsin, promatex, flavorzyme, protease N and crude commercial enzymes, 2709 alkaline protease and ASI.398 neutral protease for 4 h at enzymes’ optimal conditions of pH and temperature. All the enzymes yielded WPH that possessed antioxidant activity. Flavorzyme and ASI.398 neutral protease derived hydrolysates having the highest Diphenyl-picrylhydrazyl (DPPH) scavenging activity while 2709 alkaline protease yielded the highest DH. Trypsin-derived WPH was readily soluble in the DPPH assay solution compared to other WPH and was therefore used to investigate the effect of other hydrolysis conditions. Temperature, pH, substrate concentration and enzyme to substrate (E/S) ratio, were shown to influence DH and DPPH scavenging activity of the trysin-derived WPH. There was no correlation of DH and DPPH scavenging activity.
PDF Abstract XML References Citation
Received: March 31, 2010;
Accepted: May 06, 2010;
Published: April 29, 2011
How to cite this article
Samuel Mburu Kamau and Rong-Rong Lu, 2011. The Effect of Enzymes and Hydrolysis Conditions on Degree of Hydrolysis and DPPH Radical Scavenging Activity of Whey Protein Hydrolysates. Current Research in Dairy Sciences, 3: 25-35.
DOI: 10.3923/crds.2011.25.35
URL: https://scialert.net/abstract/?doi=crds.2011.25.35
DOI: 10.3923/crds.2011.25.35
URL: https://scialert.net/abstract/?doi=crds.2011.25.35
INTRODUCTION
Whey Protein Hydrolysates (WPH) represent a rich and heterogeneous protein source with wide range of attributes for nutritional, biological and functional purposes (Smithers et al., 1996). The biofunctional activities of whey proteins has gained prominence in the recent years, driven by demand from consumers, for save foods devoid of artificially synthesized ingredients. WPHs are capable of arresting radical-mediated oxidation of fats, the main cause of deterioration of oil-containing foods, during processing and storage (Pena-Ramos and Xiong, 2003; Elias et al., 2005). The utilization of protein hydrolysates or peptides to improve the antioxidant activity in foods presents additional advantages over other natural antioxidants, since they also confer nutritional value.
Bovine whey proteins, which are mainly comprised of β-lactoglobulin (55-60%) and α-lactalbumin (15-20%), are sources of antioxidative peptides (Bayram et al., 2008; Peng et al., 2009) that can be derived using different enzymes and different hydrolysis conditions. Additionally, antioxidant activity of Whey Protein Isolate (WPI) was reported by Pena-Ramos et al. (2004) in a lipid oxidation measured as thiobarbituric acid reactive substances (TBARS). However, this property is undetected in many studies, probably due to the detection methods employed. Whey Protein Concentrate (WPC) which is reported to be antioxiditive in cooked beef (Shantha and Decker, 1995) is a widely used raw material for production of bioactive peptides due to its abundance availability in different quantities and qualities.
Antioxidative peptides possessing higher radical scavenging activity than butylated hydroxyanisole have been obtained by enzymatic hydrolysis of whey proteins or purified β-lactoglobulin, the major whey protein (Hernandez-Ledesma et al., 2005). Similar antioxidative peptides have been identified in milk protein isolates (Hogan et al., 2009) and casein proteins (Lopez-Exposito et al., 2007; Gomez-Ruiz et al., 2008). The properties of the WPH are dependent on composition of amino acid residuals. Enzymes are extremely specific in the bonds they hydrolyze depending on the environment of the particular peptide bond. The extent of hydrolysis is influenced by the hydrolysis conditions (temperature, pH, E/S ratio and reaction time). Denaturation of whey proteins through heat treatment increases their susceptibility to hydrolysis by some enzymes (Rao et al., 1997), however, poor solubility and formation of aggregates impart negatively on overall reaction and may lead to low antioxidant activity of its hydrolysates owing to poor solubility and poor separation.
In order to derive antioxidative peptides from whey proteins, the enzyme must be able to hydrolyze specific peptide bonds within the chain of individual whey protein fractions. The hydrolysis factors have an effect on both the WPC and the enzyme activity and consequently the antioxidant activity of the derived peptides. The production of antioxidative peptides, therefore, requires a careful choice of hydrolysis conditions and enzymes as investigated by present experiments. Peptides with high antioxidant activity would be preferred since they are economical owing to being effective in low concentrations and minimizes any adverse effects arising from their incorporation in foods. These peptides can be added to food to retard lipid oxidation which has been associated with deterioration of food quality during processing and storage through flavor changes, nutrients loss and occasional generation of toxic substances (Kanner and Rosenthal, 1992). The content of beneficial health amino acids from WPC hydrolysis depend on DH (Gad and Sayed, 2009).
The objectives of the present study were to investigate the effectiveness of the different enzymes in hydrolyzing untreated commercial WPC and their ability to release peptides with DPPH scavenging activity. The influence of hydrolysis parameters (independent factors) on the DPPH scavenging activity and the final DH of the trypsin-derived WPH were also investigated.
MATERIALS AND METHODS
This study was conducted in the School of Food Science and Technology of Jiangnan University, Wuxi from January 2009 to June 2009.
Materials and reagents: Whey protein concentrate (NZMP alacenTM392, 80% protein) was a product of NZMP Ltd., New Zealand. Trypsin from porcine pancreas (EC.3.4.21.4), 4x10 3 μ g-1, 2709 alkaline protease, 2x10 5 μ g-1 and ASI.398 neutral protease, 1.3x10 5 μ g-1 enzymes were obtained from Wuxi Xuemei Enzyme Technology Co. Ltd., Jiangsu, People’s Republic of China. Promatex (EC 3.4.24.28), 1x10 5 μ g-1 and flavorzyme (EC 3.4.11.1), 2x10 4 μ g-1 were from Novo Nordisk Inc., Bagsvard, Denmark while protease N (IUB 3.4.24.28), 2.5x105 was a product of Amano Enzyme Company (Nagoya, Japan). 1,1-diphenyl-2-picrylhydrazyl (DPPH) was purchased from Sigma Chemical Co. (St. Louis, MO, USA). All other chemicals and reagents used in the experiments were of analytical grade.
Hydrolysis of WPC: Enzymatic hydrolysis was performed in a thermostatically controlled 0.5 L stirred, tank-type batch reactor. Measured WPC was dispersed in deionized water and then transferred into the batch reactor. The temperature of the solution was raised to the required hydrolysis temperature and adjusted to the hydrolysis pH with 0.5 N NaOH. Hydrolysis was commenced by addition of enzyme at given enzyme to substrate ratio. The dropping hydrolysis pH was maintained at the desired value by continuous addition of 0.1 N NaOH. The reaction was allowed to proceed for 1 or 4 h after which the pH was adjusted to 7 using 10 % HCl. The enzyme was inactivated by placing the solution in boiling water for 10 min. Separation of the soluble peptide fraction from the unhydrolyzed protein was achieved through centrifugation at 8000 rpm for 30 min at 4°C. The supernatant WPH were lyophilized and used for other analysis.
Determination of degree of hydrolysis: The pH stat method was used for quantification of DH (Adler-Nissen, 1986). The amount of 0.1 N NaOH required to maintain a constant pH during the hydrolysis period was used to calculate the DH. The DH, defined as the percent ratio of the number of peptide bonds broken (h) to the total number of peptide bonds in the substrate (htot) was therefore calculated using Eq. 1.
| (1) |
where, B is the amount of NaOH consumed (mL) during the hydrolysis; Nb is the normality of NaOH; MP is the mass (g) of protein and α is the average degree of dissociation of the α-NH2 groups released during hydrolysis expressed as Eq. 2
| (2) |
where, pK is the average dissociation value for the α-amino groups liberated during hydrolysis and is dependent on temperature, peptide chain length and the nature of the terminal amino acid. The total number of peptide bonds (htot) in whey protein was assumed to be 8.8 meq g-1 (Adler-Nissen, 1986).
Scavenging activity of DPPH radical: The antioxidant activity of the WPH was determined by its ability to scavenge DPPH radical which was measured according to the method of Shimada et al. (1992) with some modification. Lyophilized WPH was solubilized in deionized water to make a sample with a concentration of 10 mg mL-1. Two milliliter of the aliquot WPH was mixed with 2 mL DPPH solution (0.1 mM DPPH in ethanol freshly prepared). The mixture was allowed to stand for 30 min at room temperature while being kept under dark condition. The blank was conducted in the same manner, except that deionized water was used instead of the sample while a control comprising of 2 mL of the aliquot WPH mixed with 2 mL ethanol was used. The absorbance of the sample, blank and control were determined at 517 nm using a spectrophotometer (Unico, 2000, China). The radical scavenging ability of the tested samples was measured as a decrease in the absorbance of DPPH radical (Shimada et al., 1992) and was calculated by using Eq. 3.
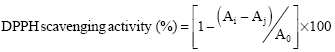 | (3) |
where, Ai, Aj and Ao represent absorbance of sample, control and blank, respectively.
Protease selection: The six proteases (trypsin EC 3.4.21.4, protease N IUB 3.4.24.28, promatex EC 3.4.24.28, flavorzyme EC 3.4.11.1 and 2709 alkaline protease and ASI.398 neutral protease) were evaluated for their suitability in hydrolyzing WPC and their ability to derive peptides with antioxidant activity. About 2709 alkaline protease and ASI.398 neutral protease are both commercial crude endopeptidases while the rest are common enzymes available in pure technical grades. The pH stat method as described by Adler-Nissen (1986) was used to screen the proteases by identifying those with better capabilities of hydrolyzing WPC resulting to high DH that is associated with production of short peptides described to have high antioxidant activity. The antioxidant activity was determined by the ability of the WPH to scavenge the DPPH radical. The solubility of the WPH in the DPPH assay solution was put into consideration.
Effects of hydrolysis variables on DH and DPPH radical scavenging: The single factor experiments were designed to estimate the effects of individual factors, namely; substrate concentration, E/S ratio, pH and temperature, on DH and DPPH radical scavenging. Each factor was varied at four levels while the other factors being held constant. Substrate concentration and E/S ratio were varied from 1 to 4%, temperature varied from 45 to 60°C and pH varied from 7.0 to 8.5 (Fig. 1). The WPC was hydrolyzed for 1 h using trypsin EC 3.4.21.4. pH stat method was used to determine the final DH. The neutralized WPH were separated and lyophilized and their DPPH scavenging activities (10 mg mL-1) were determined.
Statistical analysis: The experimental results are expressed as Mean±standard Deviation (SD) of at least three measurements. The significance (p<0.05) between the means was preformed by the one-way Analysis of Variance (ANOVA) using statistical package for social science (SPSS 13.0) for Windows® software (SPSS Inc., Chicago, IL, USA). Post-hoc analysis for significance was obtained using Tukey’s Honestly Significant Difference (HSD) test. All the curves were fitted with Microsoft® Office Excel (Microsoft Corporation, Redmond, WA, USA). The error bars on the charts were fixed as SD or at 5%.
RESULTS AND DISCUSSION
Hydrolysis of WPC: Properties of hydrolysates are mainly dependent on the primary structure of the peptide residuals as a consequence of the substrate protein, type of hydrolyzing enzyme, the DH, the environmental conditions and the substrate pretreatment. A careful choice of process parameters is prerogative to successful hydrolysis. Preparation of protein hydrolysates for food use requires the elimination of residual enzymatic activity after reaction, which is usually achieved by heat denaturation of the enzyme or by change of pH.
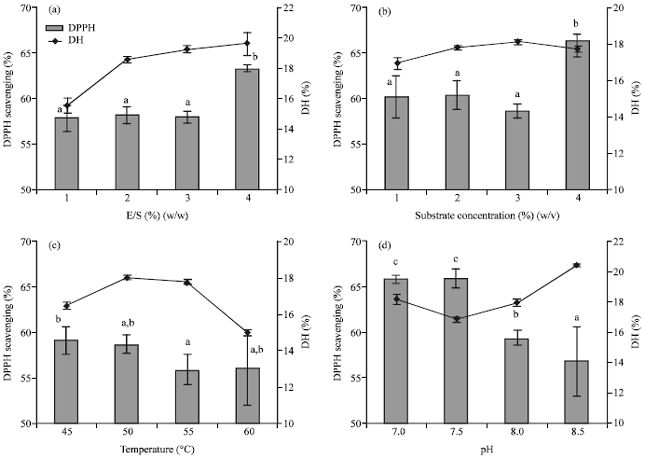 | |
| Fig. 1: | Effect of hydrolysis variables on DH and DPPH radical scavenging activity of trypsin-derived WPH (a) Enzyme to substrate ratio, Hydrolysis conditions: WPC concentration 30 mg mL-1, pH 8, temperature 50°C and hydrolysis time of 1 h, (b) Substrate concentration, Hydrolysis conditions: pH 8, temperature 50°C, enzyme to substrate ratio of 2% (w/w) and hydrolysis time of 1 h, (c) Temperature Hydrolysis conditions: WPC concentration 30 mg mL-1, pH 8, enzyme to substrate ratio of 2% (w/w) and hydrolysis time of 1 h and (d) pH, Hydrolysis conditions: WPC concentration 30 mg mL-1, temperature 50°C, enzyme to substrate ratio of 2% (w/w) and hydrolysis time of 1 h. Different letter(s) indicates activity values that are significantly different (α = 0.05) |
The inactivation of the enzymes by manipulation of the hydrolysates’ pH was not a choice because of the need to neutralize to pH 7 and therefore heat inactivation was adopted. However, this treatment can alter the structure of the peptides and change the functional properties of the hydrolysates such as the decrease in free peptides and increased formation of aggregates in the heat-treated hydrolysates (Britten et al., 1994). Aggregates were most evident in WPH obtained under high temperatures and had poor separation.
Degree of Hydrolysis (DH): The DH is a measure of the extent of hydrolytic degradation of proteins and is also the most used indicator for comparison among different proteolytic processes. The principle of the pH stat technique is that, when hydrolysis is carried out at neutral or alkaline conditions, dissociation of protons from the free amino groups released is favored (Adler-Nissen, 1986). The liberation of protons into the surrounding medium leads to a reduction in the pH of the reaction mixture. The number of peptide bonds cleaved was estimated from the amount of base required to maintain a constant pH during the reaction (Adler-Nissen, 1986). All hydrolytic curves showed a high initial rate of reaction in the first 20 min but proteolysis rate decreased gradually in the following time. Adler-Nissen (1986) attributed the reduction in hydrolysis rate to the competition between unhydrolyzed protein and the peptides being constantly formed during hydrolysis. The rate of reaction with all the enzymes leveled out after one hour owing to reduced number of susceptible peptide bonds except for the 2709 alkaline protease enzyme, which showed steady rate up to the end of the 4 h hydrolysis. It can be concluded that, the enzymatic activity of the 2709 alkaline protease was least affected by the resultant peptides and the substrate concentration changes. The enzyme, which is a commercial crude endopeptidase had a high activity of 2x105 μ g-1 and a wide pH range of 9-12 and is known to hydrolyze big molecule proteins in to peptides and amino acids (Feng et al., 2001) hence the observed high DH. At the same, it was interesting to note that high declared activities of the enzymes did not obviously correspond to a high DH. Trypsin which had the lowest declared activity of 4x10 3 μ g-1 gave a high DH. After the 4 h hydrolysis, final DH showed marked differences among the six enzymes. Other experiments showed that substrate concentration, temperature, pH and E/S ratio affected the DH (Fig. 1). Similar results have been reported by Kanu et al. (2009), when they hydrolyzed sesame flour by different proteases.
Scavenging activity of DPPH radical: Most free radicals are highly reactive and exist only for a short time. DPPH is one of the free radicals which are relatively stable at room temperature. The DPPH has commonly been used in the analysis of antioxidant activity and the test system can be used for the primary characterization of the scavenging potential of peptides. The DPPH radical has a single electron and shows strong absorbance at 517 nm. The absorbance of ethanolic DPPH solution at 517 nm reduces gradually as the free radicals are scavenged and the intensity of the color of the solution changes (Yamagushi et al., 1998) owing to the formation of the diamagnetic molecule by accepting an electron or hydrogen radical (Xie et al., 2008).
The DPPH radical scavenging activity of the WPH was dependent on enzyme used as well as the hydrolysis conditions (T, pH, E/S) that also had influence on DH. However, no relationship was established between DH and DPPH radical scavenging activity implying that, the activities of the WPH was inherent to the characteristic amino acid sequences of peptides derived, depending on the protease specificity (Chen et al., 1995). Similar results were reported for wheat germ protein hydrolysates and alfalfa leaf protein hydrolysates (Xie et al., 2008). The WPH possibly contained amino acids and peptides that were electron and hydrogen donors, or possessed radical trapping ability.
Given that, the response of peptides can be influenced by the accessibility to the oxidant-antioxidant test systems, solubility in assay solvents as well as their purity, other antioxidant activity tests are required to better characterize the WPH.
Protease selection: The DPPH scavenging activities and the final DH of the WPH (Table 1) attained with different enzymes were used for selection of enzyme with which to carry other investigations. The highest DH of 29.53±0.59% was achieved with the 2709 alkaline protease enzyme, which also had the least DPPH scavenging activity of 62.77±0.36%. This low activity was probably due to extensive hydrolysis resulting to free amino acids that have been reported to have low antioxidant activity compared to peptides (Zhou and Decker, 1999; Hernandez-Ledesma et al., 2005).
| Table 1: | The DH and DPPH scavenging activity of WPH (concentration of 10 mg mL-1) obtained with various enzymes |
 | |
| Hydrolysis conditions: WPC concentration 30 mg mL-1, enzymes to substrate ratio of 2% (w/w) and hydrolysis time of 4 h. pH and temperature (°C) of hydrolysis are indicated in brackets respectively. Different letters indicate values that are significantly different (α = 0.05) | |
Flavourzyme® showed the lowest ability to hydrolyze the WPC, yielding a DH value of only 9.06%, although it gave a comparatively high DPPH scavenging activity of 72.60±0.34%. Flavourzyme EC 3.4.11.1, a fungal protease from Aspergillus oryzae with both endo-and exo-peptidase activities was recently reported to derive hydrolysates from canola protein that were effective as antioxidants in model systems, mainly by scavenging of free radicals and acting as reducing agents (Cumby et al., 2008). Flavourzyme and Protamex EC 3.4.24.28, another endo protease used in our experiments have been used to hydrolyze WPI for 1 h producing fractions with antioxidant activities that were measured with a liposome-oxidizing system (Pena-Ramos et al., 2004).
The highest DPPH scavenging activity (73.05±0.61%) was attained by the enzyme ASI.398 neutral protease, a crude commercial preparation with endoproteolytic properties (Shen et al., 2002). ASI.398 neutral protease has been used to produce hydrolysates from fish protein (Wang et al., 1996) and used to hydrolyze snail foot protein (Xia et al., 2007). Protease N (IUB 3.4.24.28, Bacillus subtilis) enzyme was previously used to hydrolyze WPI in a tangential flow filter membrane reactor (Cheison et al., 2006). The desalted fractions of WPH were reported to possess a high DPPH radical scavenging and were largely composed of low amounts of hydrophilic amino acids ca. 45 mg of amino acids per 100 g of protein (Cheison et al., 2007). The protease N was also recently reported to derive hydrolysates from royal jelly proteins that showed a strong antioxidative activity against peroxidation of linoleic acid (Guo et al., 2009) and the hydrplysates’ low molecular weight fraction (<1 kDa) had the greatest antioxidant activity.
Trypsin EC 3.4.21.4, a serine endoprotease readily available in high purity as well as in technical grade and quantities showed both high DH and high DPPH scavenging activity (Table 1). Trypsin-derived antioxidative peptides from whey proteins have been previously reported (Bayram et al., 2008). All the WPH except that of trypsin were difficult to separate from undigested WPC by using centrifugation and their lyophilized WPH were unclear on dissolution with deionized water and subsequently had poor solubility in ethanol and DPPH solution. For this reason and considering the need for high DH, trypsin was chosen for hydrolysis parameter tests although it did not have the highest DPPH scavenging activity. High solubility of trypsin WPH was earlier reported (Mutilangi et al., 1996; Svenning et al., 2000). The ease of separation and high solubility of trypsin WPH may be accounted for by presence of small size peptides and higher hydrophilicity, a property that was associated with high solubility of hydrolysates (Mahmoud, 1994). Trypsin hydrolyzes hydrophilic sites (lysine or arginine residues) on the protein to form hydrophilic peptides.
Effects of hydrolysis variables on DH and DPPH scavenging: A relationship between DH and E/S ratio was observed. The DH increased significantly when the E/S ratio was doubled to 2%, however, further increase in this ratio up to 4% did not yield significant increase in DH (Fig. 1a) which may be attributed to either enzyme being saturated with the substrate or occurrence of enzyme inhibition by the derived peptides. The low DH of 15.52%, observed at E/S 1% may have been caused by lack of substrate saturation or was probably due to the presence of soluble peptides that irreversibly inhibited the enzyme proportionately more than at the higher E/S ratios. A high E/S ratio was necessary as native whey proteins particularly α-lactalbumin are resistant to hydrolysis by trypsin (Jean-Marc et al., 2006). Pellegrini et al. (1999) reported higher rates of tryptic hydrolysis when using high E/S ratio (10%) compared to lower ratios. The resistance of α-lactalbumin to tryptic hydrolysis is attributed to the compact conformation of the native α-lactalbumin molecule stabilized by the presence of bound calcium at the pH of reaction (pH 8). The analysis of the hydrolysis curve revealed a linear relationship between initial rate of reaction and E/S ratio. Higher concentrations gave higher initial rates of reaction. This was similar to results reported by Rao et al. (1997), when they hydrolyzed whey proteins with trypsin. A high DPPH scavenging activity value of 63.19%, at E/S ratio of 4% (Fig. 1a) may have been due to increased rate of splitting certain bonds which resulted to antioxidative peptides. At low E/S only few peptide bonds are hydrolyzed (Adler-Nissen, 1986) giving long peptides that poorly dissolve in DPPH solution. Low E/S ratio has also been associated with hydrolysates having properties different from those obtained using medium and high ratios (Adler-Nissen, 1986).
Substrate concentration had a significant influence on DH; showing a large increase in DH at concentrations above 2%, which did not increase with subsequent increase in concentration of up to 4% (Fig. 1b). The addition of NaOH to maintain pH caused further dilution of the low substrate concentration and could have negatively impacted on the stabilizing effect of substrate on the enzyme. This could have led to autodigestion of the unfolded enzyme and hence its inactivation resulting to a low DH at substrate concentration of 1%. Pace and McGrath (1980) reported that substrates stabilise and protect enzymes from the effects of hydrolysis conditions such as temperature, pH and hydrolysate inhibition as well as self-hydrolysis. The range of substrate concentrations used in our experiments was expected to produce same rate of reaction with only minor differences in overall reaction owing to minor viscosity differences. A high DPPH scavenging activity of 66.16%, at substrate concentration of 4% (Fig. 1b) was attributed to the increased specificity of the enzyme to cleave certain bonds owing to better enzyme stabilization by the substrate at that concentration.
The variation of hydrolysis temperature led to change in reaction rate. High temperatures correspondingly gave high initial rate of reaction. The increase in DH at higher temperatures, in present study observed at 50°C (17.74%) and 55°C (17.98%) (Fig. 1c), has been reported when wheat proteins were hydrolyzed by fungal protease (Bombara et al., 1992). A decline in DH to 14.96% observed at 60°C was most likely caused by heat inactivation of the enzyme in course of the 1 h hydrolysis. This effect offset the desirable whey protein denaturation that would have increased susceptibility of the protein to enzyme digestion. The DPPH scavenging activities of the WPH were in the same range (Fig. 1c), although, more errors were encountered in WPH produced at higher temperature probably due to separation problems emanating from aggregations (Britten et al., 1994).
A high DH at a particular pH is typical of many enzyme catalyzed reactions, where pH causes changes in the biological or chemical nature of reactants. Among these changes, possible denaturation of the protein structure of the enzyme or the disturbance of the ionic character of the substrate would in turn affect the ability of the substrate to bind to the enzyme (Boikess et al., 1986). The change in pH affects both the substrate and the enzyme by changing the charge distribution and conformation of the molecules. Consequently, WPH obtained at different pH possessed different properties. The highest DH of 20.53% was reached at pH 8.5 (Fig. 1d). Increase in DH at pH above 7.5 was attributed to the enzyme’s higher catalytic effectiveness at its optimum pH. Interestingly, pH 7 had also a high DH of 18.12% that was unexpected, given that, the recommended pH for the enzyme was pH 7.5-8.5. This high value could have been as a result of conformational changes of the more resistant á-lactalbumin and hence its vulnerability to trypsin. Indeed, this assumption was supported by the high DPPH scavenging activity of 65.70% (Fig. 1d), implying possible presence of antioxidative peptides derived from α-lactalbumin. Verification of this assumption by identification of the peptides is required. In addition high DPPH scavenging activities obtained when the hydrolysis was conducted at near neutral pH may be attributed to the high solubility of the peptides formed. Solubility of WPH has been reported to be close to a 100% at neutral pH (Britten et al., 1994).
CONCLUSIONS
The extent of hydrolysis of untreated WPC as determined by DH and the potency of antioxidative peptides in the WPH is influenced by the hydrolysis conditions namely; substrate concentration, temperature, pH, E/S ratio and hydrolysis time. A high DH is attained when conditions render the protein susceptible to enzyme attack while at the same time offering optimal enzyme activity. A high DH, though beneficial where low molecular weight fractions are required, does not necessarily result to a high antioxidant activity in WPH. Pure and crude enzymes that are commercially available can derive antioxidative peptides from whey proteins.
ACKNOWLEDGMENT
This study was supported by both Government of Kenya and Government of People’s Republic of China through Chinaese Government Scholarship Scheme. The authors are very grateful to the two governments.
REFERENCES
- Bayram, T., M. Pekmez, N. Arda and A.S. Yalcin, 2008. Antioxidant activity of whey protein fractions isolated by gel exclusion chromatography and protease treatment. Talanta, 75: 705-709.
CrossRef - Britten, M., H.J. Giroux and V. Gaudin, 1994. Effect of pH during heat processing of partially hydrolyzed whey protein. J. Dairy Sci., 77: 676-684.
Direct Link - Cheison, S.C., Z. Wang and S.Y. Xu, 2006. Hydrolysis of whey protein isolate in a tangential flow filter membrane reactor II. Characterisation for the fate of the enzyme by multivariate data analysis. J. Membrane Sci., 286: 322-332.
CrossRef - Cheison, S.C., Z. Wang and S.Y. Xu, 2007. Preparation of whey protein hydrolysates using a single and two stage enzymatic membrane reactor and their immunological and antioxidant properties: Characterization by multivariate data analysis. J. Agric. Food Chem., 55: 3896-3904.
PubMedDirect Link - Chen, H.M., K. Muramoto and F. Yamauchi, 1995. Structural analysis of antioxidative peptides from soybean β-conglycinin. J. Agric. Food Chem., 43: 574-578.
CrossRefDirect Link - Cumby, N., Y. Zhong, M. Naczk and F. Shahidi, 2008. Antioxidant activity and water-holding capacity of canola protein hydrolysates. Food Chem., 109: 144-148.
CrossRef - Elias, R.J., D.J. Mcclements and E.A. Decker, 2005. Antioxidant activity of cysteine, tryptophan and methionine residues in continuous phase beta-lactoglobulin in oil-in-water emulsions. J. Agric. Food Chem., 53: 10248-10253.
CrossRefDirect Link - Feng, Y.Y., W.B. Yang, S.L. Ong, J.Y. Hu and W.J. Nig, 2001. Fermentation of starch for enhanced alkaline protease production by constructing an alkalophilic Bacillus pumilus strain. Applied Microbiol. Biotechnol., 57: 153-160.
Direct Link - Gad, A.S. and A.F. Sayed, 2009. Evaluation the degree of whey protein concentrate 80 hydrolysis on the health benefits amino acids content need of the human body. Int. J. Dairy Sci., 4: 91-99.
CrossRefDirect Link - Gomez-Ruiz, J., I. Lopez-Exposito, A. Pihlanto, M. Ramos and I. Recio, 2008. Antioxidant activity of ovine casein hydrolysates: Identification of active peptides by HPLC-MS/MS. Eur. Food Res. Technol., 227: 1061-1067.
CrossRefDirect Link - Guo, H., Y. Kouzuma and M. Yonekura, 2009. Structures and properties of antioxidative peptides derived from royal jelly protein. Food Chem., 113: 238-245.
CrossRefDirect Link - Hernandez-Ledesma, B., A. Davalos, B. Bartolome and L. Amigo, 2005. Preparation of antioxidant enzymatic hydrolysates from α-lactalbumin and β-lactoglobulin. Identification of active peptides by HPLC-MS/MS. J. Agric. Food Chem., 53: 588-593.
CrossRefDirect Link - Hogan, S., L. Zhang, J. Li, H. Wang and K. Zhou, 2009. Development of antioxidant rich peptides from milk protein by microbial proteases and analysis of their effects on lipid peroxidation in cooked beef. Food Chem., 117: 438-443.
CrossRef - Jean-Marc, C., S. Mahmoud, E.Z. Khaled, D. Michele, C. Yvan and H. Thomas, 2006. Study of conformational changes of ewe's holo (native) and apo-α-lactalbumin by spectroscopy and trypsinolysis. J. Food Biochem., 30: 390-404.
CrossRefDirect Link - Kanner, J. and I. Rosenthal, 1992. An assessment of lipid oxidation in foods. Pure Applied Chem., 64: 1959-1964.
Direct Link - Kanu, P.J., J.B. Kanu, E.H. Sandy, J.B.A. Kandeh, P.M.P. Mornya and Z. Huiming, 2009. Optimization of enzymatic hydrolysis of defatted sesame flour by different proteases and their effect on the functional properties of the resulting protein hydrolysate. Am. J. Food Technol., 4: 226-240.
CrossRefDirect Link - Lopez-Exposito, I., A. Quiros, L. Amigo and I. Recio, 2007. Casein hydrolysates as a source of antimicrobial, antioxidant and antihypertensive peptides. Le Lait, 87: 241-249.
CrossRefDirect Link - Mahmoud, M.I., 1994. Physicochemical and functional properties of protein hydrolysates in nutritional products. Food Technol., 48: 89-113.
Direct Link - Pace, C.N. and T. Mcgrath, 1980. Substrate stabilization of lysozyme to thermal and guanidine hydrochloride denaturation. J. Biol. Chem., 255: 3862-3865.
Direct Link - Pellegrini, A., U. Thomas, N. Bramaz, P. Hunziker and R.V. Fellenberg, 1999. Isolation and identification of three bactericidal domains in the bovine α-lactalbumin molecule. Biochim. Biophys. Acta, 1426: 439-448.
Direct Link - Pena-Ramos, E.A. and Y.I. Xiong, 2003. Whey and soy protein hydrolysates inhibit lipid oxidation in cooked pork patties. Meat Sci., 64: 259-263.
Direct Link - Pena-Ramos, E.A., L.X. Youling and E.A. Guillermo, 2004. Fractionation and characterisation for antioxidant activity of hydrolysed whey protein. J. Sci. Food Agric., 84: 1908-1918.
Direct Link - Peng, X., Y.L. Xiong and B. Kong, 2009. Antioxidant activity of peptide fractions from whey protein hydrolysates as measured by electron spin resonance. Food Chem., 113: 196-201.
CrossRefDirect Link - Rao, P.R., P.V. Diwan, A. Margot, E. Flaschel and A. Renken, 1997. Empirical kinetic models for tryptic whey protein hydrolysis. Process Biochem., 32: 217-223.
CrossRefDirect Link - Shantha, N.C. and E. Decker, 1995. Conjugated linoleic acid concentrations in cooked beef containing antioxidants and hydrogen donors. J. Food Lipids, 2: 57-64.
CrossRefDirect Link - Shimada, K., K. Fujikawa, K. Yahara and T. Nakamura, 1992. Antioxidative properties of xanthan on the autoxidation of soybean oil in cyclodextrin emulsion. J. Agric. Food Chem., 40: 945-948.
CrossRefDirect Link - Smithers, G.W., F.J. Ballard, A.D. Copeland, K.J. De Silva and D.A. Dionysius et al., 1996. New opportunities from the isolation and utilization of whey proteins. J. Dairy Sci., 79: 1454-1459.
Direct Link - Svenning, C., J. Brynhildsvold, T. Molland, T. Langsrud and G.E. Vegarud, 2000. Antigenic response of whey proteins and genetic variants of [beta]-lactoglobulin-the effect of proteolysis and processing. Int. Dairy J., 10: 699-711.
Direct Link - Xia, S.H., Z. Wang and S.Y. Xu, 2007. Characteristics of Bellamya purificata snail foot protein and enzymatic hydrolysates. Food Chem., 101: 1188-1196.
CrossRef - Yamaguchi, T., H. Takamura, T. Matoba and J. Terao, 1998. HPLC method for evaluation of the free radical-scavenging activity of foods by using 1,1-diphenyl-2-picrylhydrazyl. Biosci. Biotechnol. Biochem., 62: 1201-1204.
CrossRefPubMedDirect Link - Zhou, S. and E.A. Decker, 1999. Ability of amino acids, dipeptides, polyamines and sulfhydryls to quench hexanal, a saturated aldehydic lipid oxidation product. J. Agric. Food Chem., 47: 1932-1936.
CrossRefDirect Link