
E. M. Marei
Department of Microbiology, Faculty of Agriculture, Ain Shams University, Cairo, Egypt
LiveDNA: 20.23985
ORCID: 0000-0001-7761-608X
I. Elmaghraby
Central Laboratory of Organic Agriculture, Agricultural Research Center, Giza, Egypt
Current Research in Bacteriology
Year: 2016 | Volume: 9 | Issue: 1 | Page No.: 1-8
DOI: 10.3923/crb.2016.1.8







ABSTRACT
Attempts were made to protect Bacillus subtilis, which can be used as a biofertilizer against bacteriophage infection. A bacteriophage resistant mutant of Bacillus subtilis was successfully isolated. Inocula of B. subtilis in forms of free cells, alginate immobilized cells and phage resistant mutant were prepared. Under cultivated soil conditions, inoculation of wheat plants with either immobilized cells or phage resistant mutant of B. subtilis resulted in increase in numbers of spore forming bacteria in the rhizosphere soils and significantly increased plant height (cm), fresh and dry weigh (gram per plant) as compared to plants inoculated with the free cells of B. subtilis. Presence of bacteriophage, in soil cultivated with wheat plants inoculated with free cells of B. subtilis, markedly reduced number of spore forming bacteria in rhizosphere soil and significantly reduced plant height (cm), fresh and dry weight (gram per plant) as compared to plants inoculated with free cells of B. subtilis in absence of phage. In plants inoculated with immobilized cells or phage resistant mutant of B. subtilis, no significant effect for the presence of phage was detected. The phage susceptible cells of B. subtilis and the phage resistant mutant ones were examined by scanning electron microscopy. The scanning electron micrographs indicated that the isolated phage resistant mutant of B. subtilis exhibited slight difference in the dimension of the cells and number of the pili as compared to the susceptible cells.
PDF Abstract XML References Citation
Received: November 12, 2015;
Accepted: February 18, 2016;
Published: June 15, 2016
How to cite this article
E. M. Marei and I. Elmaghraby, 2016. Protection of Bacillus subtilis Against Bacteriophage Attack. Current Research in Bacteriology, 9: 1-8.
DOI: 10.3923/crb.2016.1.8
URL: https://scialert.net/abstract/?doi=crb.2016.1.8
DOI: 10.3923/crb.2016.1.8
URL: https://scialert.net/abstract/?doi=crb.2016.1.8
INTRODUCTION
Soil fertility improvement is one of the most common tactics to increase agricultural production. Chemical fertilizer is widely used to supply essential nutrients for plant to increase yield. In fact, yield of most crop plants are increased linearly with the amount of fertilizer that they absorb (El-Haddad et al., 1993). Microorganisms employed to enhance the availability of nutrients are biofertilizers (Mohammadi and Sohrabi, 2012).
The use of biofertilizer could replace chemical fertilizers and pesticides. In addition to the immense cost of chemical fertilizers, their prevalent use in agriculture is also associated with problems such as environmental pollution, health hazards, interruption of natural ecological nutrient cycling and destruction of biological communities (Karuppiah and Rajaram, 2011). Therefore, biofertilizers are used as an alternative to chemical fertilizers for increasing soil productivity and plant growth in sustainable agriculture, thus received vast attention in recent times (Mohammadi and Sohrabi, 2012). They can dissolve insoluble phosphate and produces plant growth promoting substances such as IAA, amino acids, vitamins etc. (Zahran et al., 1995; Principe et al., 2007; Kumar et al., 2011). Bacillus species are a major component of the microbial flora, which live in close association with various types of agricultural crops. Predominance of Bacillus sp. is due to its ability to efficiently use the nutrients provided by the plant through exudates (Joshi and Bhatt, 2011). Bacillus subtilis is one such commercialized organism and it acts against a wide variety of pathogenic fungi. These bacteria competitively colonize the roots of plant and can act as biofertilizers or antagonists (biopesticides) or simultaneously both (Hurek and Reinhold-Hurek, 2003). Use of these microbes as fertilizers in the field has been reported to increase crop growth and yield.
For such economically important microorganism (Bacillus subtilis), knowledge of factors influencing the survival, establishment and activities of these desired bacteria in the soil is of a particular interest. Therefore, presence of bacteriophages is likely to be one of the most important factors influencing the maintenance and activities of these bacteria. Presence of bacteriophages specific to Bacillus megaterium had a depressive effect on the efficiency of these bacteria in dissolving phosphate (Zayed, 1998; Hammad, 1999).
Hammad (1998) and Fathy (2004) found that immobilization system offered high protection to Bacillus megaterium against their specific phages and increased their efficiency in dissolving phosphate. In addition, Hammad (1999) isolated phage resistant mutant of Bacillus megaterium. Presence of bacteriophages did not affect the efficiency of the isolated mutant in dissolving insoluble phosphate in pure liquid cultures and under cultivated soil conditions.
Upon the above mentioned information, the presence of bacteriophages may affect the density and activity of such important bacteria (Bacillus subtilis) in the soil. Therefore, this investigation was carried out as an attempt to protect such desired bacteria against phage attack via the immobilization system and isolation of phage resistant mutant of B. subtilis. Moreover, the morphological differences between the phage susceptible cells of Bacillus subtilis and the phage resistant mutant were also studied using scanning electron microscopy.
MATERIALS AND METHODS
Phage source: A virulent phage isolate specific to Bacillus subtilis was used in this study. This phage was previously isolated by Marei (2013) from a soil sample collected from Matruh Governorate, Egypt.
Bacterial isolate: Identified Bacillus subtilis isolate which used in this study, was kindly provided by Cairo MIRCEN (Microbial Resource Center), Faculty of Agriculture, Ain Shams University, Cairo, Egypt.
Preparation of high titer phage suspension: For preparing the high titer phage suspension, agar double layer plates method of Maniatis et al. (1982) was used as described by Hammad and Dora (1993). Agar double layer plates were prepared. The top layer contained a mixture of 3 mL of semi-solid nutrient agar medium. There were 300 mL of liquid culture of indicator bacteria (Bacillus subtilis) and 50 mL of the phage suspension (107 pfu mL–1). After incubation at 30-35°C for 24-48 h plates showed almost complete lysis. Five milliliter of SM medium (general procedure for plating and titering phage stock by Maniatis et al. (1982)) which contains per liter 5.8 g NaCl, 2 g MgSO4.7H2O, 50 mL of 1 M tris-HCl (pH 7.5) and 5 mL of 2% gelatin, were added to the surface of each plate. The top agar layer of each plate was scraped off and combined in a flask together with the added SM. After 30 min of occasional shaking, agar and bacterial debris were sedimented by centrifugation at 4000 rpm for 30 min. The supernatant containing the phages was stored at 4°C over 3 mL of chloroform.
Titer estimation: Titer of the prepared phage suspension was estimated using the method described by Kiraly et al. (1970). From the phage suspension a series of tenfold dilution was prepared in sterile eppendorf tubes. Ninety milliliter of SM medium were placed in each tube. Ten milliliters of phage suspension were added to the first vial and mixed, then 10 mL from the first vial were transferred into the second one and so on until the last tube. After dilution, 300 mL of indicator bacterial liquid culture (B. subtilis) were placed in each tube. The contents of each tube were Shaken and transferred to a sterile test tube containing 3 mL of semi-solid medium (nutrient agar) which had been prepared before hand and kept at 50-55°C. Each tube was shaken separately and the contents were poured onto previously prepared solid media plates. Plates were then incubated at 30-35°C for 24-48 h. The formed plaques were counted and the titer was calculated and expressed as plaque forming unit (pfu mL–1).
Isolation of phage-resistant mutant: The method described by Adams (1966) was used. One milliliter liquid bacterial culture containing 108 cells of B. subtilis was mixed with 1 mL of phage suspension containing 1010 plaque forming unite in an eppendorf tube. The tube was incubated for 5 min at 30-35°C to ensure that all bacteria, which can adsorb phages, were infected. One hundred microliters of the adsorption mixture was placed on the surface of a plate containing nutrient agar medium and spread uniformly with a glass rod until all the liquid had been adsorbed by agar. After incubation for 24-48 h, single colonies appeared. A single colony was picked from this plate, suspended in 1 mL of nutrient broth and from this suspension a 1oopful was streaked on another plate. Two repetitions of this procedure (streaking on agar plates) were carried out to obtain a pure isolate of phage-resistant mutant free from contaminating phages. A liquid culture of the obtained pure isolate of phage resistant mutant was prepared to be used as inoculum.
Preparation of inocula
Phage resistant mutant and wild type of B. subtilis inocula: The used bacterial isolate (wild type of B. subtilis) and the isolated phage resistant mutant were grown in Erlenmeyer flasks, each containing 100 mL of nutrient broth medium (Allen, 1959) and incubated in a shaker at 30-35°C for 96 h. (giving 25-30×108 cell per milliliter). These liquid cultures were used as inocula.
Sodium alginate-immobilized cells inoculums: One hundred milliliter of a sterile solution of sodium alginate (2% w/v) was mixed with an equal volume of the prepared liquid culture of phage susceptible B. subtilis. The mixture was added drop-wise into 200 mL of 2% CaCl2 sterile solution using a sterile Pasteur pipette. Beads of approximately 2 mm in diameter were formed and hardened in 2% CaCl2 solution for 2 h before washing. The beads were then washed with sterilized water and stored at 4°C. All steps were carried out under aseptic conditions.
Scanning electron microscopy: Differences between susceptible and resistant cells of B. subtilis to phage infection were investigated using scanning electron microscopy. The phage susceptible B. subtilis and the isolated phage resistant mutant were examined by scanning electron microscope (Jeol-SEM) in Center of Minute Analysis and Environmental Research, Faculty of Science, Beni-Souf University. The steps of Jeol-SEM protocol were carried out for preparing the samples.
Soil used: A clay loam soil was collected from the surface 15 cm layer of the Experimental Farm of Faculty of Agriculture, Ain Shams University, Shoubra, Cairo, Egypt. The mechanical and chemical analysis of the soil used is presented in Table 1. The collected soil was used for cultivation of wheat plants.
Experimental design and treatments: A pots experiment was carried out to evaluate the efficiency of free, immobilized cells and phage resistant mutants of B. subtilis as biofertilizers in presence and absence of phages. Fired clay pots containing 1 kg soil/pot were prepared and autoclaved at 121°C for 60 min. The pots were planted with wheat grains (Gemaza 11) supplied by Agriculture Research Center (ARC) Cairo, Egypt. Pots were subjected to the following treatments:
| • | Inoculation with free cells of phage susceptible B. subtilis |
| • | Inoculation with free cells of phage susceptible B. subtilis plus phage suspension |
| • | Inoculation with immobilized cells of phage susceptible B. subtilis |
| • | Inoculation with immobilized cells of phage susceptible B. subtilis plus phage suspension |
| • | Inoculation with phage resistant mutant of B. subtilis |
| • | Inoculation with phage resistant mutant of B. subtilis plus phage suspension |
| • | Control (uninoculated plants) |
Five replicates for each treatment were employed and plants were thinned to seven plants in each pot.
| Table 1: | Mechanical and chemical properties of the used soil |
| The mechanical and chemical analysis of the used soil were carried out at the Central Lab, Faculty of Agriculture, Ain Shams University | |
In the treatments inoculated with free cells of either the phage susceptible or phage resistant mutant of B. subtilis, 5 mL of the prepared liquid cultures inocula were added to each pot. In case of inoculation with the immobilized cells, a calculated weight of beads containing the same number of bacterial cells (in the 5 mL of free cells inoculum) was added to each pot. For inoculation with phage, 5 mL of the high titre phage suspension were added to each pot.
Determinations: Numbers of spore forming bacteria were determined in rhizosphere soils of different treatments at intervals of 15 days up to 75 days. The standard plate method was used for estimating the number of spore forming bacteria. A serial dilution for each rhizosphere soil sample was prepared in test tubes. The tubes containing dilutions were pasteurized for 15 min at 80°C. Nutrient agar medium (Allen, 1959) was used. The prepared petri plates were incubated at 30°C for 6 days then the colonies were counted and expressed as CFU g–1 (Hammad, 1983).
Plant height (cm), fresh and dry weight (gram per plant) were determined when plants were 75 days old.
Statistical analysis was carried out according to Gomez and Gomez (1976) using L.S.D parameter at 5%.
RESULTS
Titer of the prepared phage suspension: Two hundred milliliter of high titer phage suspension were prepared to be used in this study. Titer of the prepared suspension was estimated and was found to be 8×1010 pfu mL–1.
Phage resistant mutant of B. subtilis: As shown in Fig. 1b, the isolated mutant of B. subtilis exhibited resistance to the bacteriophage of the wild type (phage susceptible B. subtilis) i.e., no lysis was detected on the plate seeded with the mutant and spotted with the phage suspension. Whereas, lysis of the wild type can be clearly seen (Fig. 1a).
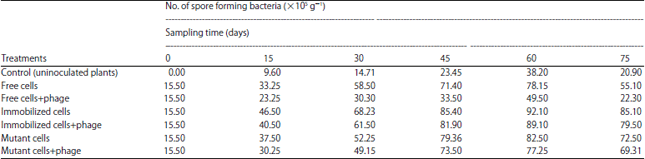
Spore forming bacteria in rhizosphere soil of wheat plants: Data presented in Table 2 indicates that under any inoculation treatment number of spore forming bacteria in rhizosphere soil of wheat plants tended to increase progressively and reached their maximum when plants were 60 days old, then decreased.
| Table 2: | Densities of spore forming bacteria in rhizosphere soil of wheat plants treated with different forms of Bacillus subtilis inoculation in presence and absence of phages |
 | |
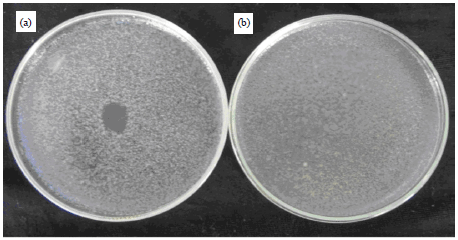 | |
| Fig. 1(a-b): | Bacterial lawns of the wild type (a) and the phage resistant mutant (b) of B. subtilis, spotted with the phage lysate. Susceptibility of the wild type and resistance of the mutant can be clearly seen |
At any sampling time, the rhizosphere soil of wheat plants inoculated with the immobilized cells of B. subtilis contained much higher number of spore forming bacteria than those inoculated with the free cells.
Presence of phages markedly reduced the number of spore forming bacteria in the rhizosphere soil of wheat plants inoculated with free cells of B. subtilis as compared to those inoculated with free cells in the absence of phages. On the other hand, the presence of phages had no pronounced effect on densities of spore forming bacteria in rhizosphere soil of wheat plants inoculated with immobilized cells.
Moreover, presence of phages had no pronounced effect on the density of spore forming bacteria in rhizosphere soil of wheat plants which were inoculated with the phage resistant mutant.
Growth measurements of wheat plants: Data presented in Table 3 indicate that, the plant height (cm), fresh and dry weight (gram per plant) of wheat plants inoculated with free cells of phage susceptible B. subtilis plus phages were lower than in the other inoculation treatments. Inoculation of wheat plants with the immobilized cells of phage susceptible B. subtilis significantly increased the studied measurements as compared to the other treatments, even in the presence of the phage. No significant differences were detected in the studied measurements of the plants inoculated with phage resistant mutant and those inoculated with the mutant plus phage.
Scanning electron microscopy of phage susceptible and phage resistant cells of B. subtilis: The scanning electron micrographs of phage susceptible cells and the phage resistant mutant of B. subtilis (Fig. 2a-b) show normal size and tubes shaped cells.
| Table 3: | Height, fresh and dry weight of wheat plants inoculated with free, immobilized or phage resistant mutant of Bacillus subtilis in presence and absence of phages, after 75 days from planting |
 | |
 | |
| Fig. 2(a-b): | Scanning electron micrograph of (a) Phage susceptible cells and (b) Phage resistant mutant cells of Bacillus subtilis, Magnification = 10,000x |
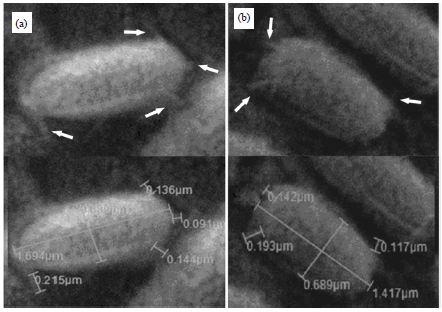 | |
| Fig. 3(a-b): | Scanning electron micrograph of (a) Phage susceptible cells and (b) Phage resistant mutant cells of Bacillus subtilis. The pili can be clearly seen on the surface of the cells, Magnification = 20,000x |
The average dimensions of the single rod shaped susceptible cells were found to be 1.529 μm in length and 0.611 μm in diameter. Whereas, the single cells of the phage resistant mutant were found to be 1.718 μm in length and 0.675 μm in diameter.
Moreover, as shown in Fig. 3a-b four pili were observed on the surface of the phage susceptible cell of B. subtilis. The lengths of the observed pili were found to be 0.091, 0.136, 0.144 and 0.215 μm. Whereas, three pili were detected on the cell surface of phage resistant mutant. These pili were 0.11, 0.14 and 0.19 μm in length.
DISCUSSION
The phage suspension used in this study was prepared using agar double layer plates showing almost complete lysis. The titer of this phage suspension was found to be (8×1010 pfu mLf). Such high concentration of phages were not surprising, since a single plaque of 2 mm in diameter may contain between 107 and 108 recoverable phage particles (Adams, 1966; Hammad, 1998; El-Balkhi et al., 2012).
A phage resistant mutant of B. subtilis was successfully isolated. Similarly, Defives et al. (1996), Coakley et al. (1997), Hammad (1999) and El-Balkhi et al. (2012) isolated phage resistant mutants of B. megaterium and Azospirillum sp.
Under any inoculation treatment numbers of spore forming bacteria in rhizosphere soil of wheat tended to increase progressively and reached their maximum when plants were 60 days old, then decreased. This may be due to the multiplication rate of these bacteria as a result of qualitative changes in the nature of root exudates of the plants during the different growth stages (Abdel-Ati et al., 1996; Hammad, 1999; Fathy, 2004; El-Balkhi et al., 2012).
The immobilization system provides suitable conditions for growth and multiplication of the cells inside the beads and bacterial cells may liberate into the surrounding soil at high population levels exceeding those applied as free cells. Therefore, number of spore forming bacteria in rhizosphere soil of wheat plants which received immobilized cells of B. subtilis was much higher than in case of inoculation with the free cells. Similar results were obtained by Van Elsas et al. (1991), Fathy (2004) and El-Balkhi et al. (2012).
Moreover, the immobilization system may protect the cells against phage attack. This may be due to the presence of the host cells inside alginate beads, which may prevent the direct adsorption of phage particles on the bacterial surface and hence no infection can be occurred. Therefore, the presence of phages had no pronounced effect on densities of spore forming bacteria in rhizosphere soil of wheat plants which inoculated with immobilized cells. Similar results were obtained by Hammad (1998), Fathy (2004) and El-Balkhi et al. (2012).
The isolated mutant of B. subtilis exhibited high resistance to phage infection. Therefore, presence of phages had no pronounced effect on the density of spore forming bacteria in rhizosphere soil of wheat plants inoculated with the phage resistant mutant. These results are in agreement with those obtained by Hammad (1999) and Fathy (2004).
Biofertilizers are used as an alternative to chemical fertilizers for increasing soil productivity and plant growth in sustainable agriculture. Bacillus species is a major component of the microbial flora, which live in close association with various types of agricultural crops. They can dissolve insoluble phosphate and produce plant growth promoting substances such as IAA, amino acids, vitamins etc. Bacillus subtilis is one of these commercialized organisms which can be used as a biofertilizer. These microbes as fertilizers in the field have been reported to increase crop growth and yield (Zahran et al., 1995; Principe et al., 2007; Kumar et al., 2011). Therefore, this may explain the high growth values (height, dry and fresh weight/plant) in wheat plants inoculated with Bacillus subtilis. Moreover, bacteriophages had a depressive effect on their bacterial host (Bacillus subtilis). Therefore, low values of plant growth were recorded in plants inoculated with free cells of (Bacillus subtilis) plus phages. Whereas, no pronounced effect for phages was detected in plants inoculated with the immobilized cells of Bacillus subtilis. This may indicate that the immobilization system provides these bacteria with high resistance against phages. Similar results were obtained by Hammad (1998), Fathy (2004) and El-Balkhi et al. (2012).
The isolated phage resistant mutant of B. subtilis exhibited slight difference in the dimensions of the cells and number of the pili as compared to the susceptible cells as indicated by the scanning electron micrographs. This may indicate that the mutation process altered the morphological properties of the cell. Similarly, Barnet (1972) reported that symbiotic-defective mutants of root nodule bacteria were isolated as surviving cells after exposure to specific virulent phages. The isolated mutants exhibited changes in morphological, physiological and symbiotic properties as compared to the susceptible cells. In addition, Kay (1972) stated that the specific receptors sites on the bacteria, to which specific attachment proteins bind. These receptor sites may be lipopolysaccharides, cell wall proteins, teichoic acid, or flagellar or pilus protein.
CONCLUSION
Generally, on the basis of the obtained results it can be concluded that, presence of a bacteriophage specific to B. subtilis in the soil is one of the most important environmental factors influencing the activity and maintenance of such desired microorganisms in the soil. Whereas, application of such bacteria as a biofertilizer for the growing plants, in alginate immobilized form or phage resistant mutant may provide these bacteria with high resistance against phages and promote their efficiency as a biofertilizer. Therefore, application of B. subtilis in alginate immobilized form or phage resistant mutant as a biofertilizer is highly recommended to promote their efficiency and to avoid the phage attack.
ACKNOWLEDGMENT
The authors would like to express their sincere thanks, special gratitude and appreciation to Prof. Dr. Adel M. Hammad, Prof. of Agriculture Viruses, Faculty of Agriculture, Minia University for his valuable help, supporting, giving advices and fruitful discussions during this investigation. Thanks also to Ahmed A. Abd-Elwahab, Assistant Lecturer of Agriculture Viruses, Faculty of Agriculture, Ain Shams University for his help during this investigation.
REFERENCES
- Barnet, Y.M., 1972. Bacteriophages of Rhizobium trifolii L. morphology and host range. J. Gen. Virol., 15: 1-15.
CrossRefDirect Link - Coakley, M., G. Fitzgerald and R.P. Ros, 1997. Application and evaluation of the phage resistance- and bacteriocin-encoding plasmid pMRC01 for the improvement of dairy starter cultures. Applied Environ. Microbiol., 63: 1434-1440.
Direct Link - Defives, C., M. Werquin, P. Mary and J.P. Hornez, 1996. Roles of exopolysaccharides and lipopolysaccharides in the adsorption of the Siphovirus phage NM8 to Rhizobium meliloti M11S cells. Curr. Microbiol., 33: 371-376.
CrossRefDirect Link - El-Balkhi, M.A., O.A.O. Saad and A.M.M. Hammad, 2012. Protection of phosphate dissolving bacteria against bacteriophage attack. J. Basic Sci., 22: 43-57.
Direct Link - El-Haddad, M.E., Y.Z. Ishac and M.I. Mostafa, 1993. The role of biofertilizers in reducing agricultural costs, decreasing environmental pollution and raising crop yield. Arab. Univ. J. Agric. Sci. Ain Shams Univ. Cairo, 1: 147-195.
Direct Link - Hammad, A.M.M., 1998. Evaluation of alginate-encapsulated Azotobacter chroococcum as a phage-resistant and an effective inoculum. J. Basic Microbiol., 38: 9-16.
CrossRefDirect Link - Hammad, A.M.M., 1999. Induction of bacteriophage-resistant mutants of nitrogen fixing and phosphate dissolving bacteria. Ann. Agric. Sci. Cairo, 44: 479-493.
Direct Link - Hurek, T. and B. Reinhold-Hurek, 2003. Azoarcus sp. strain BH72 as a model for nitrogen-fixing grass endophytes. J. Biotechnol., 106: 169-178.
CrossRefDirect Link - Joshi, P. and A.B. Bhatt, 2011. Diversity and function of plant growth promoting Rhizobacteria associated with wheat Rhizosphere in North Himalayan region. Int. J. Environ. Sci., 1: 1135-1143.
Direct Link - Karuppiah, P. and S. Rajaram, 2011. Exploring the potential of chromium reducing Bacillus sp. and there plant growth promoting activities. J. Microbiol. Res., 1: 17-23.
CrossRefDirect Link - Kiraly, Z., Z. Klement, F. Solymosy and J. Voros, 1970. Methods in Plant Pathology with Special Reference to Breeding for Disease Resistance. 2nd Edn., Academia Kiado, Budapest, Hungary, pp: 183-192.
Direct Link - Kumar, K., N. Amaresan, S. Bhagat, K. Madhuri and R.C. Srivastava, 2011. Isolation and characterization of rhizobacteria associated with coastal agricultural ecosystem of rhizosphere soils of cultivated vegetable crops. World J. Microbiol. Biotechnol., 27: 1625-1632.
CrossRefDirect Link - Maniatis, T., E.F. Fritsch and J. Sambrock, 1982. Molecular Cloning: A Laboratory Manual. 3rd Edn., Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, USA.
Direct Link - Marei, E.M., 2013. Isolation and characterization of Bacillus subtilis phage from soil cultivated with liquorices root. Int. J. Microbiol. Res., 4: 43-49.
Direct Link - Mohammadi, K. and Y. Sohrabi, 2012. Bacterial biofertilizers for sustainable crop production: A review. J. Agric. Biol. Sci., 7: 307-316.
Direct Link - Principe, A., F. Alvarez, M.G. Castro, L. Zachi, S.E. Fischer, G.B. Mori and E. Jofre, 2007. Biocontrol and PGPR features in native strains isolated from saline soils of Argentina. Curr. Microbiol., 55: 314-322.
CrossRefDirect Link - Van Elsas, J.D., L.S. Van Overbeek, A.M. Feldmann, A.M. Dullemans and O. De Leeuw, 1991. Survival of genetically engineered Pseudomonas fluorescens in soil in competition with the parent strain. FEMS Microbiol. Lett., 85: 53-64.
CrossRefDirect Link - Zahran, H.H., M.S. Ahmad and E.A. Afkar, 1995. Isolation and characterization of nitrogen-fixing moderate halophilic bacteria from saline soils of Egypt. J. Basic Microbiol., 35: 269-275.
CrossRefDirect Link - Zayed, G., 1998. Can immobilization of Bacillus megaterium cells in alginate beads protect them against bacteriophages? Plant Soil, 197: 1-7.
CrossRefDirect Link