
R. Arunachalam
Environmental Nanotechnology Division, Sri Paramakalyani Centre for Environmental Sciences, Manonmaniam Sundaranar University, Alwarkurichi-627 412, Tamil Nadu, India
E.G. Wesely
Centre for Biotechnology, Muthayammal College of Arts and Science, Rasipuram-637408, Tamil Nadu, India
J. George
Centre for Biotechnology, Muthayammal College of Arts and Science, Rasipuram-637408, Tamil Nadu, India
G. Annadurai
Environmental Nanotechnology Division, Sri Paramakalyani Centre for Environmental Sciences, Manonmaniam Sundaranar University, Alwarkurichi-627 412, Tamil Nadu, India
Current Research in Bacteriology
Year: 2010 | Volume: 3 | Issue: 1 | Page No.: 15-26







ABSTRACT
The present study was aimed at to identify the cellulase producing Actinomycetes from soil of Southwest ghats, Tamilnadu, India. The morphology, cultural, physiological and chemo taxonomical analysis and phylogenetic analysis of TBG-V20 were characterized. According to the morphological, isolate TBG-V20 was identified as a representative of the genus Streptomyces. The 16S rRNA region of this strain was amplified and sequenced. A Neighbor-Joining algorithm tree of 16S rRNA was constructed. In endocellulase activity, the Carboxy Methyl Cellulose (CMCase) activities of the strain TBG-V20 on eighth day an amount of 935 U L-1 of glucose, 225 μg mL-1 of protein and 870 mg/100 mL of growth (biomass) on tenth day were recorded. In exocellulase activity strain TBG-V20 on first day an amount of 500 μg mL-1 glucose was produced. According to the observation and phylogenetic analysis, the strain TBG-V20 was proved to belong to the species Streptomyces noboritoensis with cellulase production.
PDF Abstract XML References Citation
How to cite this article
R. Arunachalam, E.G. Wesely, J. George and G. Annadurai, 2010. Novel Approaches for Identification of Streptomyces noboritoensis TBG-V20 with Cellulase Production. Current Research in Bacteriology, 3: 15-26.
DOI: 10.3923/crb.2010.15.26
URL: https://scialert.net/abstract/?doi=crb.2010.15.26
DOI: 10.3923/crb.2010.15.26
URL: https://scialert.net/abstract/?doi=crb.2010.15.26
INTRODUCTION
Streptomycetes are the largest and well-studied group of actinomycetes. A wide variety of bacteria are known for their production of hydrolytic enzymes with streptomycetes being the best known enzyme producers (Vinogradova and Kushnir, 2003). Bacterial, fungal and actinomycete cellulase and xylanases have attracted considerable research interest because of their potential applications in recovery of fermentable sugars from hemicellulose, biobleaching of pulp and paper industry and to other industrial applications (Rawashdeh et al., 2005). Actinomycete’s, one of the known cellulase-producers, has attracted considerable research interest due to its potential applications in recovery of fermentable sugars from cellulose that can be of benefit for human consumption and to the ease of their growth (Jang and Chen, 2003) compared to anerobic cellulase producers such as Paenibacillus curdlanolyticus (Pason et al., 2006). The biotechnology applications of cellulases began in the early 1980s in animal feed followed by food applications (Harchand and Singh, 1997). Today, these enzymes account for approximately 20% of the world’s enzyme market (Jaradat et al., 2008). The Streptomyces are able to utilize a wide range of organic compounds as a carbon source, including complex biological materials such as cellulose and lignin and can also utilize an inorganic nitrogen source (Kutzner, 1986). Nowadays, enzyme cost is the major impediment to commercialization of enzymatic hydrolysis; the major challenge is to significantly increase the activity of cellulases (Jaradat et al., 2008; Walker and Wilson, 1991). Rawashdeh et al. (2005) isolated several Streptomyces isolates, upon further characterization; these isolates were able to produce cellulase, pectinase and relatively large amount of xylanase.
Determination of difference in the sequence of the 16S rRNA gene is well established as a standard method for the identification and phylogenetic classification of prokaryotic species, genera and families ( Michael and Sharon, 2007), but more recently it has also become important as a means to identify an unknown bacterium to the genus or species level. Thereby, in this investigation cellulase producing strain TBG-V20 was isolated from soil samples and characterized to belong to Streptomyces noboritoensis ( Shirling and Gottlieb, 1969) with cellulase activity by morphology, biochemical and phylogenetic analysis.
MATERIALS AND METHODS
Materials
All the chemicals used in the analytical methods and media preparation were of analytical grade with maximum available purity supplied by Hi-media (Mumbai), SISCO (Chennai) and Sigma (USA). The 16S rRNA primers were from Genei (Bangalore) and PCR reagents and Taq Polymerase from Biogene, USA. The Actinomycetes presence soil samples were collected from Southwest ghats, Tirunelveli, Tamilnadu, India.
Isolation and Screening of Cellulase Producing Actinomycetes
In this study, was conducted during the period of November 2007-March 2008, for that, the soil samples were collected aseptically from Southwest ghats, Kerala, India at a depth of 6-12 inches. The soil samples were air dried in a hot air oven at 45°C for 1 h to reduce the proportion of bacteria other than Actinomycetes (Williams et al., 1972). Standard dilution plate technique was followed for the isolation of Actinomycetes (Kuster and Williams, 1964). Ten gram each of the soil samples were added to 90 mL distilled water in a 250 mL Erlenmeyer flask under sterile condition and kept in a rotary shaker (120 rpm) at room temperature for 30 min. Vacuum filtration was used for collecting supernatant and then the supernatant was serially diluted to obtain 10-4, 10-5 and 10-6 dilutions. Each dilution was plated on sabouraud’s agar. After inoculation of 7-9 days at 27±1°C the actinomycetes colonies were selected, counted and made into pure culture following single spore culture technique. The culture was maintained on sabouraud’s agar by periodical sub-culturing.
In screening, 24 colonies were used for screening; the inorganic salts cellulose agar was prepared. Twenty milliliter of the medium was poured in to the petriplates. The isolated Actinomycetes were inoculated into different petriplates and incubated at room temperature (27±1°C) for 7 days then the plates were taken and 0.1% congored was spread over the plates. After half hour incubation 0.5 M NaCl was spread. After the 0.5 M NaCl was discarded, the presence of a clear zone indicates the cellulolytic activity of the strain. The diameter of the zone is measured and based on this the activity of the organisms were categories as weak and promising ones.
Cellulase Assay
The inorganic salts cellulose broth supplemented with 1% Carboxy Methyl Cellulose (CMCase) and 1% yeast extract was prepared. One hundred milliliter of the broth was taken in 250 mL Erlenmeyer flasks and inoculated heavily using the individual stock cultures maintained on sabouraud’s slants. The flasks in duplicate were incubated at room temperature at 120 rpm. During incubation, every day 1 mL of the broth was collected up to 12th day for CMCase assay. In endocellulase determination the β (1-4) glucanase was determined by measuring the reducing sugar as glucose by dinitrosalicyclic acid (DNS) method (Miller, 1959). The reaction mixture was contained 1 mL of w/v CMC in citrate buffer pH 5.0 and 1 mL of the broth, which was collected on first day to seven days. The assay was started by incubating the reaction mixture at 50°C for 20 min. The reaction was stopped by the addition of 3 mL DNS reagent and boiling for 5 min. After cooling the absorbance was read at 540 nm in a colorimeter. Gluscose was used as the standard and the reducing sugar formed was calculated as the glucose equivalent. One unit of CMCase was expressed as 1 μmol of reducing sugar released per min per mL. In exocellulase activity, the Whatman No.1 filter papers were cut into 4 mm dia pieces. A reaction mixture was contains 1 mL of 1% citrate buffer pH 5.0 and 1 mL of the broth was prepared. Thirty milligram filter papers were added to it. Further assay was similar to the above. For determining the amount of cellulase (protein estimation) produced by strain TBG-V20 was performed by the method of Lowry et al. (1951).
Extraction of Protein
Extraction was carried out with buffers used for the enzyme assay. Five hundred milligram of the sample was weighed, ground well and grinded well with a mortar and pestle in 5-10 mL of the buffer. Protein estimation was carried out based on method described by Lowry et al. (1951). Standard graph was sketched and calculated the amount of protein in the sample. Expressed the amount of protein in IU mL-1 of culture broth was used for calculation. Estimating a biomass, 100 mL culture was filtered through a pre-weighed and dried Whatman filter paper. The filter paper was dried at 40°C for 24 h and the weight was taken in a weighing balance. The biomass was expressed in mg mL-1. Based on the performance of cellulose activity out of 24 one colony was taken for further study.
Morphology and Taxonomy
The determination of Aerial mass colour, reverse colony colour and the 7 to 14 days old cultures growth characters in different culture media which were reported by International Streptomyces Project (ISP) (Shirling and Gottlieb, 1966). Medium ISP-1 to ISP-7 was used according to description of Shirling and Gottlieb (1966). The colour of soluble pigments were noted and compared with Methuen hand book of colour which was noted from Kornerup and Wanscher (1967). The microscopic studies were prepared according to the method described by Chakrabarthi (1998). The pure cultures of the Actinomycetes strains were inoculated to the contract line of immersed cover slips at an angle of 45° on Sabouraud’s agar plates prepared aseptically. The plates with cover slips were incubated at 28°C for 4-8 days. After attaining suitable growth with spore chains the cover slips were removed and placed on micro slides with the growth on the upper surface. The slides were viewed under a phase contrast microscope (Nikon Optiphot-II). The details of the conidial chain/sporangium, spores, Arial and substrate mycelium and fragmentation of the mycelia were noted and the slides were photographed at suitable magnifications. In physiological and biochemical studies, melanin production was determined on ISP-6 media; after 4 days of growth the colour change of media were noted by the methodology described by Shirling and Gottlieb (1966).
Carbon Source Utilization
Different organic carbon sources were sterilized using di-ethy1 ether and incorporated to carbon utilization agar (Shirling and Gottlieb, 1966). These include D-Fructose, meso-Inositol D-mannitol, Raffinose, L-Rhamnose, Sucrose and D-Xylose at 1% w/v. Results were determined after 7, 14 and 21 days by comparing with a negative control (without carbon source) and a positive control containing D-Glucose. In Chemotaxonomical studies, the Actinomyces strains were inoculated in 100 mL sabaraud’s broth in Erlenmeyer flask and incubated for 5 days at room temperature in a rotary shaker (120 rmp). The cells were killed with formalin (con.1%) for 24 h at room temperature and harvested by centrifugation. The cells were washed once in double distilled water and once in 95% ethanol and then dried by overnight heating in a hot air oven at 45°C. The dried cells were analyzed for Diaminopimilic acid (DAP) and carbohydrates (Staneck and Roberts, 1974).
Extraction of Genomic DNA
Samples used for DNA extraction were collected from a highly sproulated growth on sabouraud’s agar were inoculated to 25 mL yeast extract malt extract (YEME) broth supplemented with 0.5% glucose and 5 mM MgCl2 in a 100 mL Erlenmeyer flask, incubated for 2-3 days at room temperature at 10 rpm. Genomic DNA was isolated as described by Murray and Thompson (1980). The integrity of the obtained genomic DNA was detected by electrophoresis in 1% agarose gel (Sigma Aldrich, India) stained with ethidium bromide.
16S rRNA Amplification
The 16S rRNA was amplified using the bacterial universal primer pairs. The reaction mixture for PCR amplification was prepared in a total volume of 50 μL with 10x PCR buffer, 10 mM of dNTPs, 1.0 μL of the taq polymerase and 200 pmol of forward and reverse primers. The amplifications were performed in a DNA thermal cycler 480 (Perkin Elorer, USA). The PCR reaction details were as follows: 5 min at 95°C for initial denaturation, 1 min at 94°C for denaturation, 2 min at 49°C for annealing, 1 min at 72°C for extension with total 24 cycles of amplification and 7 min at 72°C for the final extension. The 16S rRNA (EU857664) was purified using GenEluteTM Gel Extraction Kit (Sigma Aldrich, USA) and sequencing of 16S rRNA gene was done in an automated ABI-3100 Genetic Analyser (GeNei, India).
Phylogenetic Analysis
BLASTN (optimized for megablast) searches were manipulated with the sequences of Streptomyces noboritoensis TBG-V20. The corresponding sequences of representative species were used for phylogenetic analyses. Neighbor-Joining (NJ) algorithm was used to construct a graphical phylogenetic tree using software MEGA 4.1 (Tamura et al., 2007). The program was starts with a set of aligned sequences using Clustal W and searches for phylogenetic trees that are optimal according to NJ algorithm.
GenBank Accession Number
The GenBank accession number of the sequence reported in this study is EU857664.
RESULTS AND DISCUSSION
Morphology
In this study, 24 strains were obtained and one colony was found to produce cellulase which could be used for further studies and it was termed as TBG-V20. Cellulase activity of strain TBG-V20 on inorganic salt cellulose agar was found to be 26 mm diameter zone after 7 days of incubation at 28±1°C.
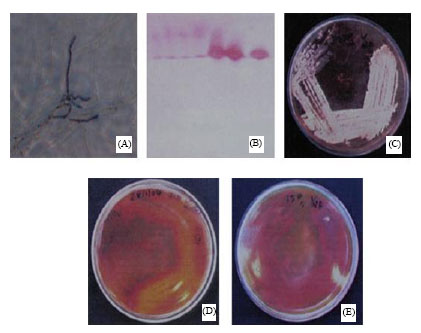 | |
| Fig. 1: | Cellulase producing strain Streptomyces noboritoensis TBG-V20. (A) Top morphology view of Aerial mycelium with conidial chains (>50 spores), (B) Presesnce of LL-Diaminopimilic acid as major cell wall peptidoglycan, (C) Predominantly white spore mass colour and (D) Reverse colony colour is yellow brown; (E) Production of melanin (Aerial mass colour) in ISP media |
The isolate TBG-V20 gives morphology similar to that of a Streptomyces isolate with a distinct substrate and aerial mycelium with conidial chains observed on microscopic examination. The conidial chains with more than 50 spores were observed and these were oblong in shape (Fig. 1A). The spore chains of Streptomyces may be of different type, straight to flexuous (Rectus-flexibilis), open loops (Relinaculam-apertum), pen of closed spirals (spira) and verticillate (Pridham et al., 1958). The microsopic examinations of the present isolate TBG-V20 shows that the conidial chains are Rectus-flexibilis. The cell wall analysis shows the presence of LL-Diaminopimilic acid (LL-DAP) as major cell wall peptidoglycan content (Fig. 1B). There were no diagnostic sugars detected in cell wall. So the isolate TBG-V20 was identified as a representative of genus Streptomyces (Waksman and Henrici, 1943).
Table 1 shows the cultural characteristic of the isolate after an incubation period of one week at 28±1°C. The spore mass color of the Streptomyces members may vary from white, gray, red, yellow, green and blue to violet. Colour of the aerial mycelium is one of the prominent identification characters of Streptomyces isolates at species level (Pridham and Tresner, 1974). On further examination of the morphological characters the isolate is having a spore mass colour of predominantly white (Fig. 1C) and occasionally grey. The International Streptomyces Project (ISP) (Shirling and Gottlieb, 1966) has recommended recording of aerial mycelial colour in different media for use as a taxonomic character, which was also followed in the present study. ISP-6 and ISP-7 were showed the reverse colony color is yellow brown and is not producing any diffusible pigments (Fig. 1D). The isolate was showing good growth on ISP media and other suggested media (Table 1). The above-mentioned characters suggest that the isolate TBG-V20 can be identified as Streptomyces groups of white series (Nonomura, 1974).
| Table 1: | Cultural characteristics of isolate TBG-V20 |
 | |
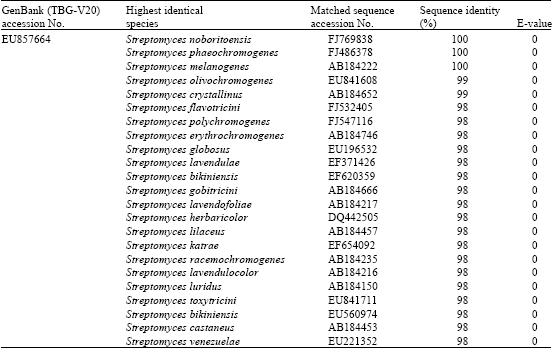
| Table 2: | Results of similarity searches between 16S rRNA genes isolated in the present investigation and GenBank accessions using BLASTN Algorithm (optimized for megablast) |
 | |
The isolate was utilizing D-fructose, Meso-ionsitol, Mannitol, Raffinose, Rhamnose and D-xylose as the carbon source. In addition, isolate TBG-V20 was not utilizing sucrose as the carbon source. The utilization of carbon compounds is an important aid for species determination among actinomycetes (Pridham and Gottlieb, 1948). The isolate TBG-V20 utilized almost all carbohydrates, but not sucrose respectively. Likewise, the pigment production stands out as a characteristic tool in identifying the various microorganisms. However, TBG-V20 does not produce any of these pigments, these minor physiological characters are not delaminating the identification of this strain as Streptomyces noboritoensis. The keys of Kuster (1972), Nonomura (1974), Szabo et al. (1975) and Shirling and Gottlieb (1966) with based on ISP description giving emphasis to aerial mass colour, melanin pigment production, spore chain morphology and carbon source utilization pattern, the isolate TBG-V20 was identified as Streptomyces noboritoensis.
Phylogenetic Analysis
A 557 bp sequence was amplified from the genome DNA with the bacterial universal primers, its sequence was submitted to GenBank (EU857664). As is shown in Table 2, strain TBG-V20 16S rRNA sequence was used for identity search, which was made using BLASTN algorithm (optimized for megablast). The 16S rRNA gene showed high similarity with 16S rRNA genes deposited in the GenBank (Table 2). TBG-V20 strain 16S rRNA had 100% identity (E value 0.0) with 16S rRNA gene of S. noboritoensis (FJ769838) followed by 100% idenity (E value 0.0) with 16S rRNA gene of S. phaeochromogenes (FJ486378) and 100% identity (E value 0.0) with 16S rRNA gene of S. melanogenes (AB184222). Moreover, TBG-V20 had 99% identity (E value 0.0) with 16S rRNA gene of S. olivochromogenes (EU841608) and S. crystallinus (AB184652) and also TBG-V20 had 98% identity (E value 0.0) with S. flavotricini (FJ532405), S. polychromogenes (FJ547116), S. erythrochromogenes (AB184746), S. globosus (EU196532), S. lavendulae (EF371426), S. bikiniensis (EF620359), S. gobitricini (AB184666), S. lavendofoliae (AB184217), S. herbaricolor (DQ442505), S. lilaceus (AB184457), S. katrae (EF654092), S. racemochromogene (AB184235), S. lavendulocolor (AB184216), S. luridus (AB184150), S. toxytricini (EU841711), S. bikiniensis (EU560974), S. castaneus (AB184453) and S. venezuelae (EU221352). In this study, 16S rRNA gene of different Streptomyces species (different strains of a species) was obtained by BLASTN search, however 23 strains of Streptomyces species were selected on the basis of high identity (%) with good E value for phylogenetic analysis. As shown in Fig. 3, two strains belonging to Streptomycetaceae were relatively closely related to Streptomyces; strain TBG-V20 had a clade supported with S. noboritoensis (FJ769838).
The cumulative results from a limited number of studies to date suggest that 16S rRNA gene sequencing provides genus identification in most cases (>90%) but less so with regard to species (65-83%), with regard to species from 1-14% of the isolates remaining unidentified after testing (Drancourt et al., 2000; Mignard and Flandrosis, 2006; Woo et al., 2003). Michael and Sharon, 2007 reported that, the minimum 500-525 bp essential for phylogenetic analysis and also for species identification minimum >99% similarity and ideal >99.5% similarity should be desirable. <0.5% similarity and other properties such as phenotype should be considered to final species identification. E value is related to the probability that the observed degree of similarity could have arisen by change: E is the number of sequences that would be expected to match as well or better than the one being considered, if the same database were probed with random sequences.
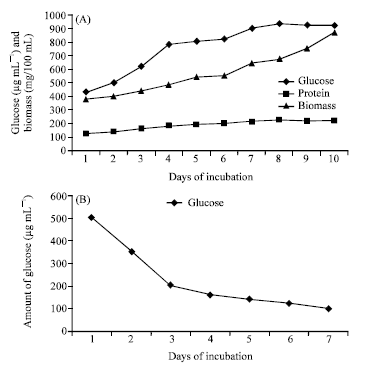 | |
| Fig. 2: | Cellulase activities of strain TBG-V20. (A) CMCase activities of the Streptomyces isolate TBG-V20; (B) Exocellulase activities of Streptomyces isolate TBG-V20 |
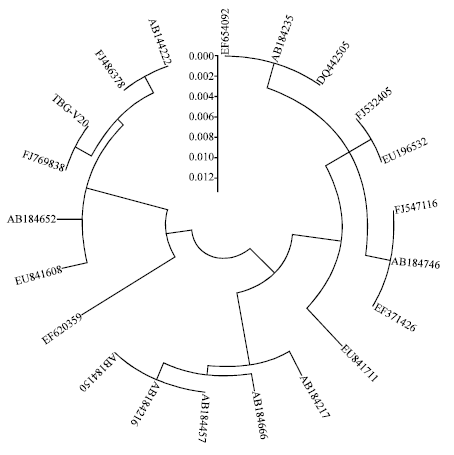 | |
| Fig. 3: | Phylogenetic tree is based on the nucleotide sequence of 16S rRNA genes. The Neighbor-Joining algorithm tree was constructed by MEGA 4.1 |
Values of E below about 0.05 would be considered significant; at least they might be worth considering (Arthur, 2005). According to Michael and Sharon, 2007 and Arthur (2005) reports, in this study, isolate TBG-V20 (557 bp) had (100%) ideal >99.5% similarity and E value (0.0) <0.05 with S. noboritoensis, S. phaeochromogenes and S. melanogenes. On the basis of BLASTN search and phenotypic results TBG-V20 was identified as a strain of S. noboritoensis.
Comparison of 16S rRNA sequences has become the ‘gold standard’ for the elucidation of phylogenetic relationships among microorganisms. As the number of sequences available for analysis continues to grow, the structure of phylogenetic trees derived from these sequences becomes both more intricate and more accurate (O’Connor et al., 1991). Pernodet et al. (1989), Chang et al. (1997) and Fukushima et al. (2002) reported that 16S rRNA, gyrB and 23S rRNA of various Streptomyces, Salmonella, Shigella and Eshcerichia coli species were partially sequenced and used for defining all members of the genus, groups of species or individual species. As shown in Fig. 3, two strains belonging to Streptomycetaceae were relatively closely related to Streptomyces; strain TBG-V20 had a branch supported with S. noboritoensis (FJ769838). The results supported that the phylogenic position of strain TBG-V20 in the genus Streptomyces might belong to the family Streptomycetaceae (Kataoka et al., 1997). The results of a phylogenetic analysis based on the 16S rRNA sequences can be suggested strain TBG-V20 should be positioned in the genus Streptomyces. In addition, the special characteristics mentioned in the study suggest that novel strain TBG-V20 belong to the species S. noboritoensis with high cellulose activity.
Cellulase Activity of the Isolate
Actinomyces have been always a source of thousand of bioactive compounds. Enzymes are one of the important products of this unusual group of bacteria. In endocellulase activity, the CMCase activites of the isolate TBG-V20 on different days of incubation is shown in Fig. 2A. The isolate was showing good CMCase activity on the eighth day. On the eighth day an amount of 935 U L-1 of glucose and 225 μg mL-1 of protein were recorded at pH 5 and 50°C temperature. The growth (biomass) was 870 on the tenth day. In exocellulase activity of the isolate TBG-V20 on different days of incubation are shown in Fig. 2B. The isolate was showing maximum exocellulase activity on first day of incubation. On the first day an amount of 500 U L-1 glucose was produced. Jaradat et al., 2008 have observed that Streptomyces strain J2 is showed the highest crude enzyme activity (432 U L-1) after 3 days of incubation at pH 7 and 60°C of temperature. This result is considerably similar to what was reported by Theberge et al. (1992) who showed that the optimum pH for endoglucanase from a strain of Streptomyces lividans was 5.5. However, the results appeared to contradict previous results reported by Solingen et al. (2001) of an alkaline novel Streptomyces species isolated from east African soda lakes that have an optimal pH of 8, highlighting the effect of alkaline environment on the adaptation of these Streptomyces. Furthermore, the maximum CMCase activity of our isolate TBG-V20 was recorded at 50°C with no significant difference between 50 and 60°C. These results are in agreement with results reported by McCarthy (1987), who reported an optimal temperature for cellulase activity in the range of 40-55°C for several Streptomyces species including S. lividans, S. flavogrisus and S. nitrosporus. Jang and Chen (2003) described a CMCase produced by a Streptomyces T3-1 with optimum temperature 50°C, whereas Schrempf and Walter (1995) described a CMCase production by a S. reticuli at an optimum temperature 55°C.
Schlochetermeier et al. (1992) and Wachinger et al. (1989) reported that Actinomycetes are playing an important role in degradation of cellulose and Streptomyces reticuli is a soil bacterium, which hydrolyzes crystalline cellulose due to the action of exocglucanase. Kamini et al. (1999) also reported Streptomyces species with cellulose activity. The isolate TBG-V20 is showing high cellulolytic activity in the preliminary screening. In this study the present strain TBG-V20 was showing good endocellulolytic activity in eighth day and exocellulolytic activity in first day which were differentiate our isolate from other scientist described strains. Isolate TBG-V20 had produced According to the above evidences, the present strain TBG-V20 was identified as a high cellulose producing Streptomyces species.
CONCLUSION
According to the results, the strain Streptomyces noboritoensis TBG-V20 has potentiality for industrial production for the reason that of the high yield of cellulase. It has been reported that the biosynthesis of cellulase is induced during growth on cellulose or other cellulose derivatives. In all cases, it has been found that it is essential to keep the required nutrients at low level to insure maximum accumulation of fermentation products. Overall, the study indicated that cellulase production from TBG-V20 isolate has potentiality for industrially important in this world for the production of the high yield of cellulase.
ACKNOWLEDGMENTS
Authors thanks to Manonmaniam Sundaranar University, Alwarkurichi, India and Muthayammal College of Arts and Science, Rasipuram, India for provided necessary facilities.
REFERENCES
- Chang, H.R., L.H. Loo, K. Jeyaseelan, L. Earnest and E. Stackebrandt, 1997. Phylogenetic relationships of salmonella typhi and salmonella typhimurium bases on 16S rRNA sequence analysis. Int. J. Syst. Bacteriol., 47: 1253-1254.
Direct Link - Drancourt, M., C. Bollet, A. Carlioz, R. Martelin, J.P. Gayral and D. Raoult, 2000. 16S ribosomal DNA sequence analysis of a large collection of environmental and clinical unidentifiable bacterial isolates. J. Clin. Microbiol., 38: 3623-3630.
Direct Link - Fukushima, M., K. Kakinuma and R. Kawaguchi, 2002. Phylogenetic analysis of Salmonella, Shigella and Escherichia coli strains on the basis of the gyrB gene sequence. J. Clini. Microbiol., 40: 2779-2785.
CrossRefPubMedDirect Link - Harchand, R.K. and S. Singh, 1997. Characterization of cellulase complex of Streptomyces albaduncus. J. Basic Microbiol., 37: 93-103.
Direct Link - Jang, H.D. and K.S. Chen, 2003. Production and characterization of thermostable cellulases from Streptomyces transformant T3-1. World J. Microbiol. Biotechnol., 19: 263-268.
CrossRef - Jaradat, Z., A. Dawagreh, Q. Ababneh and I. Saadoun, 2008. Influence of culture conditions on cellulase production by Streptomyces sp. (Strain J2). Jordan J. Biol. Sci., 1: 141-146.
Direct Link - Kamini, N.R., C. Hemachander, J. Geraldine, S. Mala and R. Puvanakrishnan, 1999. Microbial enzyme technology as an alternative to conventional chemical in leather industry. Curr. Sci., 77: 80-86.
Direct Link - Kataoka, M., K. Ueda, T. Kudo, T. Seki and T. Yoshida, 1997. Application of the variable region in 16S rRNA to create an index for rapid species identification in the genus Streptomyces. FEMS Microbiol. Lett., 151: 249-255.
CrossRef - Kuster, E., 1972. Simple working key for the classification and identification of named taxa included in the international Streptomyces project. J. Syst. Bacteriol., 22: 139-148.
Direct Link - Kuster, E. and S.T. Williams, 1964. Selection of media for isolation of streptomycetes. Nature, 202: 928-929.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - McCarthy, A.J., 1987. Lignocellulose degrading actinomycetes. FEMS Microbiol. Lett., 46: 145-163.
CrossRef - Janda, J.M. and S.L. Abbott, 2007. 16S rRNA gene sequencing for bacterial identification in the diagnostic laboratory: Pluses, perils, and pitfalls. J. Clin. Microbiol., 45: 2761-2764.
CrossRefDirect Link - Mignard, S. and J.P. Flandrois, 2006. 16S rRNA sequencing in routine bacterial identification: A 30-month experiment. J. Microbiol. Methods, 67: 574-581.
CrossRefDirect Link - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Murray, M.G. and W.F. Thompson, 1980. Rapid isolation of high molecular weight plant DNA. Nucl. Acids Res., 8: 4321-4326.
CrossRefPubMedDirect Link - O`Connor, S.P., M. Dorsch, A.G. Steigerwalt, D.J. Brenner and E. Stackebrandt, 1991. 16S rRNA sequences of bartonella bacilliformis and cat scratch disease bacillus reveal phylogenetic relationships with the Alpha-2 subgroup of the class Proteobacteria. J. Clin. Microbiol., 29: 2144-2150.
Direct Link - Pason, P., K.L. Kyu and K. Ratanakhanokchai, 2006. Paenibacillus curdlanolyticus strain B-6 xylanolytic-cellulolytic enzyme system that degrades insoluble polysaccharides. Applied Environ. Microbiol., 72: 2483-2490.
Direct Link - Pernodet, J.L., M.T. Alegre, F. Boccard and M. Guerineau, 1989. Organization and nucleotide sequence of a ribosomal RNA gene cluster from Streptomyces ambofaciens. Gene, 79: 33-46.
PubMed - Pridham, T.G. and D. Gottlieb, 1948. The utilization of carbon compounds by some Actinomycetales as an aid for species determination. J. Bacteriol., 56: 107-114.
Direct Link - Pridham, T.G., C.W. Hesseltine and R.G. Benedict, 1958. A guide for the classification of streptomycetes according to selected groups; placement of strains in morphological sections. Applied Microbiol., 6: 52-79.
PubMedDirect Link - Shirling, E.B. and D. Gottlieb, 1966. Methods for characterization of Streptomyces species. Int. J. Syst. Evol. Microbiol., 16: 313-340.
CrossRefDirect Link - Shirling, E. and D. Gottlieb, 1969. Cooperative description of type cultures of Streptomyces. IV. Species description from the second, third and fourth studied. Int. J. Syst. Evol. Microbiol., 19: 391-512.
CrossRef - Staneck, J.L. and G.N. Roberts, 1974. Simplified approach to identification of aerobic actinomycetes by thin-layer chromatography. Applied Microbiol., 28: 226-231.
Direct Link - Tamura, K., J. Dudley, M. Nei and S. Kumar, 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol., 24: 1596-1599.
CrossRefPubMedDirect Link - Theberge, M., P. Lacaze, F. Shareck, R. Morosoli and D. Kluepfel, 1992. Purification and characterization of an endoglucanase from Streptomyces lavidans 66 and DNA sequence of the gene. Applied Microbiol., 58: 815-820.
PubMed - Waksman, S.A. and A.T. Henrici, 1943. The nomenclature and classification of the actinomycetes. J. Bacteriol., 46: 337-341.
PubMed - Woo, P.C.Y., K.H.L. Ng, S.K.P. Lau, K.T. Yip and A.M.Y. Fung et al., 2003. Usefulness of the microseq 500 16S ribosomal DNA-based bacterial identification system for identification of clinically significant bacterial isolates with ambiguous biochemical profiles. J. Clin. Microbiol., 41: 1996-2001.
CrossRefDirect Link - Wachinger, G., K. Bronnenmeier, W.L. Staudenbauer and H. Schrempf, 1989. Identification of mycelium-associated cellulase from Streptomyces reticuli. Applied Environ. Microbiol., 55: 2653-2657.
PubMedDirect Link - Schrempf, H. and S. Walter, 1995. The cellulolytic system of Streptomyces retyiculi. Int. J. Biol. Macromolecules, 15: 353-355.
CrossRef - Williams, S.T., M. Shameemullah, E.T. Watson and C.I. Mayfield, 1972. Studies on the ecology of actinomycetes in soil-VI. The influence of moisture tension on growth and survival. Soil Biol. Biochem., 4: 215-225.
CrossRefDirect Link