
M.M. Afifi
Department of Botany and Microbiology, Faculty of Science, Al-Azhar University, Assuit 71524, Egypt
H.M. Atta
Department of Botany and Microbiology, Faculty of Science (Boys), Al-Azhar University, Cairo, Egypt
A.A. Elshanawany
Department of Botany and Microbiology, Faculty of Science, Al-Azhar University, Assuit 71524, Egypt
U.M. Abdoul-raouf
Department of Botany and Microbiology, Faculty of Science, Al-Azhar University, Assuit 71524, Egypt
A.M. El-Adly
Department of Botany and Microbiology, Faculty of Science, Al-Azhar University, Assuit 71524, Egypt
Bacteriology Journal
Year: 2012 | Volume: 2 | Issue: 3 | Page No.: 46-65







ABSTRACT
Hygromycin B is an aminoglycoside antibiotic that kills bacteria, fungi and higher eukaryotic cells by inhibiting protein synthesis. It is biosynthesized in this research by Streptomyces sp. AZ151, isolated from soil sample in Assuit governorate and chosen among 194 actinomycete isolates. Morphological, physiological and biochemical culture characteristics as well as 16S rRNA nucleotide analysis confirmed strain AZ151 as Streptomyces crystallinus. An optimum environmental and nutritional condition in culture medium of this strain showed strong antibacterial and antifungal activities. Using organic solvent extraction, silica gel column chromatography and TLC, a high active metabolite, Hygromycin-B, was separated and identified as a suggested empirical formula of C15H30N2O10 and confirmed a biological efficiency.
PDF Abstract XML References Citation
Received: November 02, 2011;
Accepted: April 18, 2012;
Published: May 24, 2012
How to cite this article
M.M. Afifi, H.M. Atta, A.A. Elshanawany, U.M. Abdoul-raouf and A.M. El-Adly, 2012. Biosynthesis of Hygromycin-B Antibiotic by Streptomyces crystallinus AZ151 Isolated from Assuit, Egypt. Bacteriology Journal, 2: 46-65.
URL: https://scialert.net/abstract/?doi=bj.2012.46.65
URL: https://scialert.net/abstract/?doi=bj.2012.46.65
INTRODUCTION
The genus Streptomyces is a particularly fruitful source of antifungals, antibiotics (antibacterials) and chemotherapeutic (anticancer) drugs (Raja and Prabakarana, 2011). The bacterial genus Streptomyces is a high G+C content, Gram-positive filamentous soil bacteria with a complex life cycle that includes morphological differentiation and spore formation (Del-Sol et al., 2007). Medical geography is the influence of environmental (natural and human) factors on biological system of human, plant and animals which is even recognized as an effective and basic factor in mortality (Esmaeili and Moshiri, 2011). Also, it has long been appreciated as a rich source for the production of various secondary metabolites including many pharmaceutically valuable compounds such as antibiotics, anti-cancer agents, immunosuppressants and enzyme inhibitors (Hindra and Elliot, 2010; Myles, 2003). Based on 16S rDNA sequence and phylogenetic tree analysis, Khucharoenphaisan et al. (2012) assigned Streptomyces strain FSPNRU 102 and reported that this a new isolate belong to the Streptomyces niveoruber. It is well documented that the biosynthesis of Streptomyces secondary metabolites is typically regulated via multiple regulatory pathways operating with several layers of complicated control systems (Chen et al., 2010; Lee et al., 2005). The rRNA sequences is a particularly powerful tool in streptomycete taxonomy. In addition, rRNA sequence has also been useful for answering questions concerning the horizontal transfer of genes within the genus Streptomyces (Huddleston et al., 1997).
The crude antibiotic was tested by using pre coated Thin-Layer Chromatography (TLC) plates and purified by column chromatography using silica gel (Augustine et al., 2005). The antibacterial activity of Hygromycin A (HA) arises from protein synthesis inhibition and is dependent upon a methylenedioxy bridged-aminocyclitol moiety. Selective gene deletions and chemical complementation in Streptomyces hygroscopicus NRRL 2388 showed that the hyg18 and hyg25 gene products, proposed to generate a myo-inositol intermediate, are dispensable for HA biosynthesis but contribute to antibiotic yields (Palaniappan et al., 2009). Antibacterial compound was purified from the filtrate by solvent extraction method. A comparative study on the total antibiotic sensitivity of the free cells and immobilized cells showed that the immobilized strains were found to be effective against the tested microorganisms. The immobilized cell of actinomycetes was found to be more efficient for the production of secondary metabolites with batch fermentation (Dhananjeyan et al., 2010). The morphological character showed the variety of aerial hyphae and spore forming in each strain. The dendrogram was constructed based on biodegradation activity of tested strains (Khucharoenphaisan et al., 2011).
This research has first confirmed the isolation Streptomyces strain, AZ151 out of 194 isolates from 44 soil samples at upper Egypt (Assuit governorate). Then it confirmed the identity of this strain, by morphological, physiological and biochemical characteristics as well as 16S rRNA, as Streptomyces crystallinus. Optimum environmental and nutritional requirements were carried out to obtain the highest yield of antibiotic. An antimicrobial compound was separated and purified from the culture broth of this strain through organic solvent extraction, column chromatography and TLC purification. It possesses strong inhibitory activity toward fungal and bacterial strains tested which give a chance to be applied in medical and other fields.
MATERIALS AND METHODS
Microorganism: The actinomycete AZ151 was isolated from soil sample collected from Assuit governorate. It was purified using the soil dilution plate technique described by Williams and Davies (1965).
Screening for antimicrobial activity: The anti-microbial activity was determined according to Kavanagh (1972).
Taxonomic studies of actinomycete isolate (AZ151)
Morphological characteristics: Morphological characteristics of aerial hyphae, spore mass, spore surface, color of aerial and substrate mycelia and soluble pigments production were conducted by growing the organism on starch-nitrate agar medium and Yeast extract-malt extract agar medium (Atta et al., 2011).
Physiological and biochemical characteristics: Lecithinase was detected using egg B yolk medium according to the method of Nitsch and Kutzner (1969), Lipase (Elwan et al., 1977), Protease (Chapman, 1952), Pectinase (Hankin et al., 1971), α-amylase (Ammar et al., 1998) and Catalase Test (Jones, 1949). Melanin pigment (Pridham et al., 1956). Esculin broth and xanthine have been done according to Gordon et al. (1974). Nitrate reduction was performed according to the method of Gordon (1966). Hydrogen sulphide production was carried out according to Cowan and Steel (1974). The utilization of different carbon and nitrogen sources was carried out according to Pridham and Gottlieb (1948). Determination of Diaminopimelic acid (DAP) and sugar pattern was carried out according to Becker et al. (1964) and Lechevalier and Lechevalier (1970).
Color characteristics: The ISCC-NBS Color Name Charts illustrated with centroid detection of the aerial, substrate mycelia and soluble pigments (Kenneth and Deane, 1955).
DNA isolation and manipulation: The locally isolated actinomycete strain was grown for 6 days on a starch agar slant at 30°C. Two milliliters of a spore suspension were inoculated into the starch- nitrate broth and incubated for 4 days on a shaker incubator at 200 rpm and 30°C to form a pellet of vegetative cells (pre-sporulation). The preparation of total genomic DNA was conducted as described by Sambrook et al. (1989).
Amplification and sequencing of the 16S rRNA gene: PCR amplification of the 16S rRNA gene of the local actinomycete strain was conducted using two primers, StrepF; 5.-ACGTGTGCAG CCCAAGACA-3. and Strep R; 5.ACAAGCCCTGGAAACGGGGT-3. (Edwards et al., 1989). The PCR mixture consisted of 30 pmol of each primer, 100 ng of chromosomal DNA, 200 μM dNTPs and 2.5 units of Taq polymerase, in 50 μL of polymerase buffer. Amplification was conducted for 30 cycles of 1 min at 94°C, 1 min of annealing at 53°C and 2 min of extension at 72°C. The PCR reaction mixture was then analyzed via agarose gel electrophoresis and the remaining mixture was purified using QIA quick PCR purification reagents (Qiagen, USA). The 16 S rRNA gene was sequenced on both strands via the dideoxy chain termination method (Sanger et al., 1977).
Sequence similarities and phylogenetic analysis: The BLAST program (www.ncbi.nlm.nih.gov/blast) was employed in order to assess the degree of DNA similarity. Multiple sequence alignment and molecular phylogeny were evaluated using BioEdit software (Hall, 1999).
Parameters controlling antimicrobial agent biosynthesis: These included; incubation period and temperature; agitation and aeration; pH values; carbon source and nitrogen sources; vitamins, MgSO4.7H2O and K2HPO4 concentrations; inoculum age and size, amino acids; and medium kinds. All these parameters have been determined by the standard methods.
Fermentation and purification of antibacterial agent
Fermentation: The Streptomyces crystallinus, AZ151 inoculum was introduced aseptically into each sterile flask containing the following ingredients (g L-1): Arabinose, 20; NaNO3, 2.0; K2HPO4, 0.8; MgSO4.7H2O, 0.7; vitamin H, ppm and KCl, 0.5. The pH was adjusted at 8.0 before sterilization. After 10 days of incubation at 35°C filtration was carried out through filter paper Whatman No. 1 and followed by centrifugation at 5000 rpm for 15 min. Only clear filtrates were tested for their antimicrobial activities.
Extraction: The clear filtrate was adjusted at different pH values (4-9) and extraction process was carried out using different solvents separately at the level of 1:1 (v/v). The organic phase was concentrated to dryness under vacuum using a rotary evaporator.
Precipitation: The precipitation process of the crude compound was carried out using petroleum ether (b.p. 60-80°C) followed by centrifugation at 5000 r.p.m for 15 min. The precipitate was tested for its antimicrobial activities.
Separation: Separation of the antimicrobial compound into its individual components was conducted by thin layer chromatography using chloroform and methanol (24:1, v/v) as a solvent system.
Purification: The purification of the antimicrobial compound was carried out using silica gel column (2.5x50) chromatography. Chloroform and Methanol 8:2 (v/v), was used as an eluting solvent. The column was left overnight until the silica gel (Prolabo) was completely settled. One milliliter crude extract to be fractionated was added on the silica gel column surface and the extract was adsorbed on top of silica gel. Fifty fractions were collected (each of 5 mL) and tested for their antibacterial activities.
Physico-chemical properties of antimicrobial agent
Elemental analysis: The elemental analysis of C, H, O, N and S was carried out at the microanalytical center, Cairo University, Egypt.
Spectroscopic analysis: The IR, UV and Mass spectrum were determined at the micro analytical center of Cairo University, Egypt.
Biological activity: The Minimum Inhibitory Concentration (MIC) has been determined by the cup method assay (Kavanagh, 1972).
Characterization of the antibacterial agent: The antibiotic produced by Streptomyces crystallinus, AZ151 was identified according to the recommended international references of Umezawa (1977) and Berdy (1974, 1980).
RESULTS
Isolation, purification and bioactivity of actinomycete isolates: Isolation and purification of actinomycete colonies (the broadest source of antibiotics) from 40 soil samples collected from various Egyptian localities e.g. (Assiut, Luxor and El-Minia governorates). The highest number of isolates (84) out of 194 (43.2%) were isolated on starch nitrate agar medium followed by 55 isolates (28.3%) on both starch casein agar and glycerol asparagine agar (data not shown). Screening test for 194 actinomycete isolates, against certain bacteria, fungi and yeast, confirmed that the highest percentage (38%) 74 active isolates was obtained against Staphylococcus aureus 90.5% (67) followed by Aternaria alternata, 43.2% (32) and Klebsiella pneumoniae, 41.8% (31) while the lowest percentage was obtained against Fusarium verticillioides 21.6% (16 isolates), Salmonella typhi 20.2% (15 isolates), Escherichia coli 9.4% (7 isolates), Aspergillus fumigatus 9.4% (7 isolates), Saccharomyces cerevisiae 8.1% (6 isolates) and Aspergillus flavus 6.7% (5 isolates) (data not shown).
Characterizations of the actinomycete isolate, AZ151
Morphological characteristics: Spore chains are spiral and rectiflexibiles, spore masses are medium red and reddish gray, spore surfaces are smooth and reverse color light yellow to light brown while, diffusible pigment production is moderate yellowish brown to deep brown (Plate 1).
Cell wall hydrolysate: The cell wall hydrolysate contains LL-diaminopimelic acid (LL-DAP) and sugar pattern not detected.
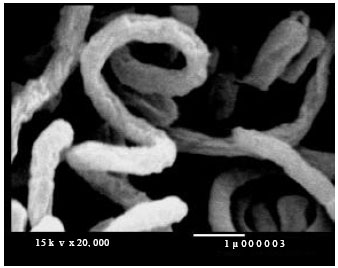 | |
| Plate 1: | Scanning electron micrograph of the actinomycete isolate AZ-A151 growing on starch nitrate agar medium showing spiral spore were, rectiflexibiles and had a smooth surface. Neither sclerotic granules, sporangia nor flagellated spores were observed (X20, 000) |
Physiological and biochemical characteristics: The actinomycete isolate AZ151 could hydrolyze starch and protein, whereas lipid, pectin and catalase are negative, Melanin pigment is positive, degradation of xanthin, esculin, production of H2S, decomposition of urea, utilization of citrate and KCN are positive but nitrate reduction is negative.
The isolate under study utilizes; D-mannose, D-glucose, D-galactose, mannitol, meso-inositol, raffinose and trehalose but do not utilize, D- xylose, sucrose, L-rhamnose, L-arabinose, lactose, maltose and ribose; whereas, doubtful result was obtained with D-fructose. Good growth on L-glycine, L-asparagine and L-lysine. No growth on L-valine, L-leucine L-histidine, L-phenyl alanine and L-methionine. Moreover, no growth in the presence of up to 5% NaCl. The growth is not inhibited in the presence of 0.1% (w/v) phenol and at 451°C but inhibited in the presence of 0.01% (w/v) sodium azide. The actinomycete isolate is resistant to Ampicillin (25 μg mL-1), Nalidixic acid (30 μg mL-1), Cefoperazone (75 μg mL-1) and Fusidic acid (10 μg mL-1), whereas not resistant to Polymyxin (30 μg mL-1), Gentamicin (10 μg mL-1) and Kanamycin (30 μg mL-1) (Table 1).
Color and culture characteristics: Data recorded on AZ151 declared that, the growth of this strain was disappeared in ISP-1, moderate in ISP-2, 4, 5, 6 and 7 and good growth on SNA and ISP-3. While, the aerial mycelium appeared red on all media used except SNA medium, it was reddish gray. Substrate mycelium light brown on all media used except ISP-5 has shown light yellow. Almost all media used didn't induce any diffusible pigments except SNA shown moderate yellowed brown and ISP-6, 7 showed deep brown pigments (data not shown).
Molecular phylogeny of the selected isolate: The nucleotide sequence of the 16S rRNA gene (1.5 kb) of the actinomycete isolates AZ151 and the phylogenetic tree (as displayed by the Tree View program) revealed 92% similarity with Streptomyces crystallinus (Fig. 2). Multiple sequence alignment was conducted the sequences of the 16S rDNA gene of Streptomyces crystallinus and the sequencing product was determined as 1141 bp (Fig. 1).
| Table 1: | The morphological, physiological and biochemical characteristics of the actinomycete isolate AZ151 |
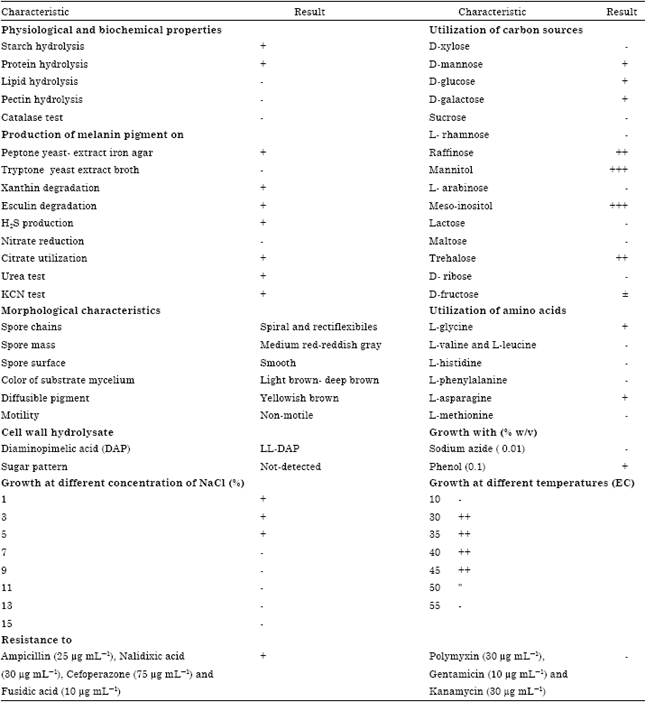 | |
| -: Negative and +: Positive. ±: doubtful results, ++: Moderate growth, +++: Good growth | |
Identification of actinomycete isolate, AZ151: This was performed basically according to the recommended international Key’s viz. (Buchanan and Gibbons, 1974; Williams, 1989) and Numerical taxonomy of Streptomyces species program (PIB WIN). On the basis of above collected data and in view of the comparative study of the recorded properties of AZ151 in relation to the most closest reference strain, viz., Streptomyces crystallinus, it could be concluded that it is identical on the basis of spore mass is medium red or reddish gray, spore chain is spiral and rectiflexibiles and non motile spores.
| Table 2: | A comparative study of the characteristic properties of AZ151 in relation to reference strain, Streptomyces crystallinus |
 | |
| ND: Melanin pigment not-detected | |
 | |
| Fig. 1: | The sequence alignment was conducted the sequences of the 16S rDNA gene of Streptomyces crystallinus |
Cell wall hydrolysate contains LL-diaminopimelic acid and sugar pattern not detected. Melanin pigments are produced. Utilization of D-mannose, D-glucose, D-galactose, mannitol, meso-inositol, raffinose and trehalose but do not utilize D-xylose, sucrose, rhamnose, L-arabinose, lactose, maltose and Ribose, whereas, doubtful with D-fructose. In view of all the previous characteristics of AZ151, it could be stated that it is suggestive of being belonging to Streptomyces crystallinus (Table 2).
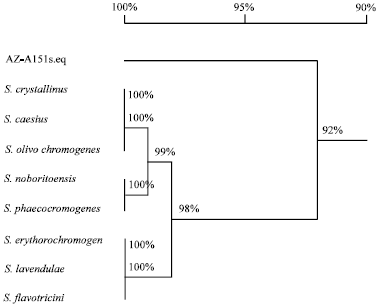 | |
| Fig. 2: | The phylogenetic position of the local Streptomyces sp. strain among neighboring species. The phylogenetic tree was based on the pairwise comparisons of 16S rDNA sequences |
Parameters controlling antimicrobial agent biosynthesis: The effect of different environmental and nutritional factors on the antimicrobial activity indicated that the maximum activities was obtained in starch nitrate broth medium at, incubation period (10 days), incubation temperature (35°C), agitation and aeration (160 rpm), pH value (8.0), carbon source (Starch), nitrogen source (NaNO3), water soluble vitamin (vitamin H), inoculum age (12 days), inoculum size.
Fermentation and isolation of antimicrobial agent: The Streptomyces crystallinus, AZ151 inoculum was introduced aseptically into each sterile flask containing the following ingredients (g L-1): Arabinose, 20; NaNO3, 2.0; K2HPO4, 0.8; MgSO4.7H2O, 0.7; vitamin H, ppm and KCl, 0.5. The pH was adjusted at 8.0 before sterilization. After 10 days of incubation at 35°C filtration was carried out through filter paper Whatman No. 1 and followed by centrifugation at 5000 rpm for 15 min. Only clear filtrate was tested for its antimicrobial activity.
The clear filtrate containing the active metabolite, was adjusted at pH 7.0 then extraction process was carried out using Ethyl acetate at the level of 1:1 (v/v). The organic phase was collected and evaporated under reduced pressure using rotary evaporator. The residual material was dissolved in least amount of DMSO and filtered. The filtrates were test for their antibacterial activities. The antimicrobial agent was precipitated by petroleum ether (b.p. 60-80 EC) and centrifuged at 4000 r.p.m for 15 min. The fraction was test for antimicrobial activities. 8-10% (v/v), amino acid (Asparagine), MgSO4.7H2O concentration (0.7 g L-1), K2HPO4 (0.8 g L-1), inoculum size (8-10 % (v/v) and at inoculum culture age (12 days) (Table 3).
Separation of antimicrobial agent into individual components was carried out by thin-layer chromatography using a solvent system composed of chloroform and methanol (24:1, v/v). Among three bands developed, only one band at Rf 0.55 showed antibacterial activity. The purification process through column chromatography packed with silica gel, indicated that maximum activity was at fractions Nos. 14-23 (Fig. 3).
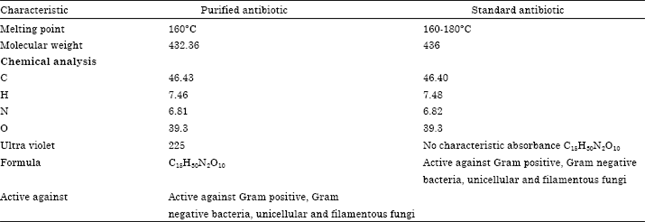
Physicochemical characteristics of the antimicrobial agent: The purified antimicrobial agent produced by Streptomyces crystallinus, AZ151 produces characteristic odour, its melting points is 160°C.
| Table 3: | The environmental conditions and nutritional requirements affecting the biosynthesis of antimicrobial agent by Streptomyces crystallinus, AZ151 |
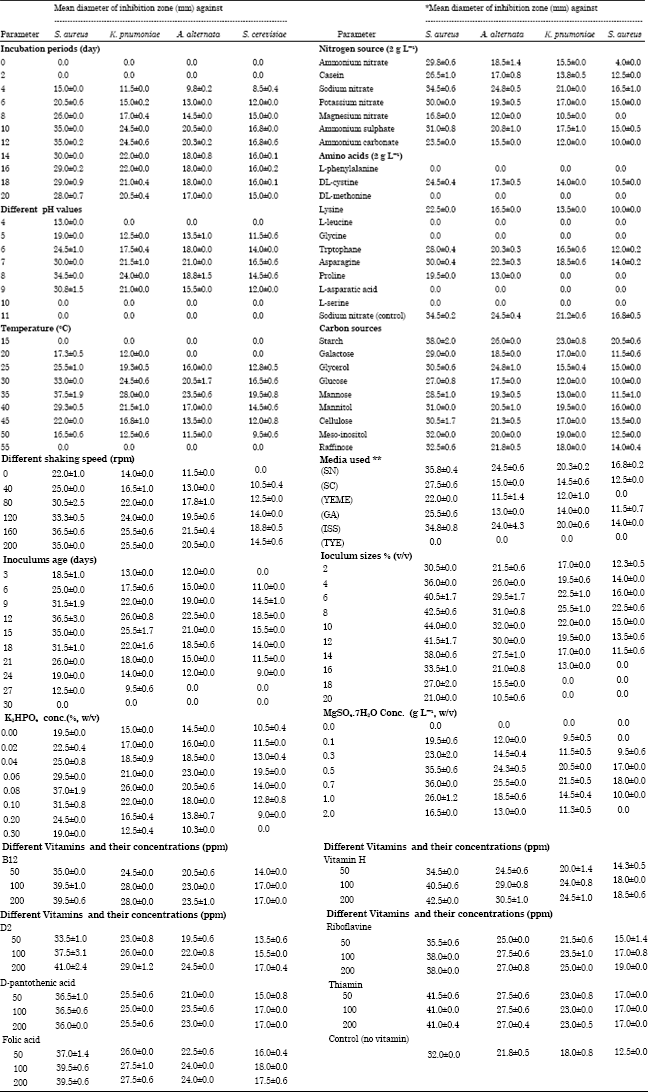 | |
| *Mean values of triplicate determinations were calculated. **SN: Starch nitrate medium, SC: Starch casein medium , YEME: Yeast extract malt extract medium, GA: Glycerol asparagine medium, ISS: Inorganic salt starch medium and TYE: Tryptone yeast extract medium | |
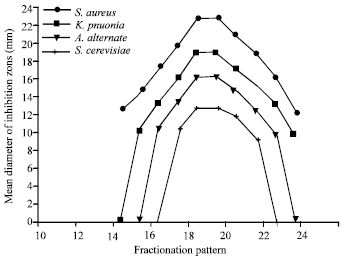 | |
| Fig. 3: | Antimicrobial activity of fractions for antimicrobial agent |
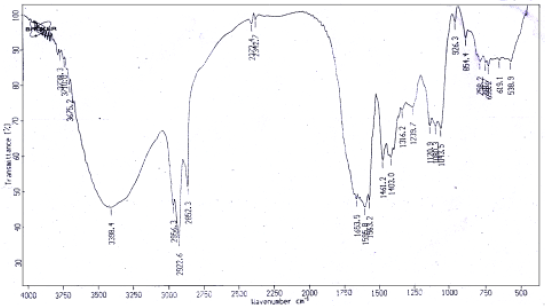 | |
| Fig. 4: | IR spectrum of antimicrobial agent |
The compound is freely soluble in chloroform, ethyl acetate, n-butanol, acetone, ethyl alcohol, methanol and 10% isopropyl alcohol but insoluble in petroleum ether, hexane and benzene.
Elemental analysis: The elemental analytical data of the antibacterial agent revealed the following data: C = 46.43; H = 7.46; N = 6.81; O = 39.43 and S = 0.0. This analysis indicates a suggested calculated empirical formula of C15H30N2O10.
Spectroscopic characteristics: The spectroscopic analysis of purified antimicrobial agent produced by Streptomyces crystallinus, AZ151 have been determined. The infrared (IR) spectrum showed characteristic band corresponding to 21 peaks (Fig. 4), the ultraviolet (UV) spectrum recorded a maximum absorption peak at 225 NM (Fig. 5) and the Mass spectrum indicate that the molecular weight was 432.36 (Fig. 6).
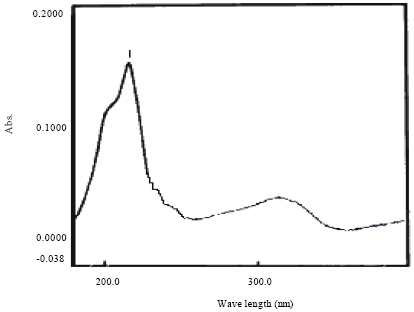 | |
| Fig. 5: | Ultraviolet absorbance of antimicrobial agent |
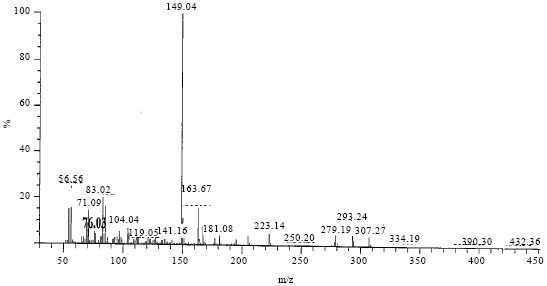 | |
| Fig. 6: | Mass spectrum of antimicrobial agent |
Identification of the antimicrobial agent: On the basis of the recommended keys for the identification of antibiotics and in view of the comparative study of the recorded properties of the antimicrobial agent, it could be stated that the antimicrobial agent is suggestive of being belonging to Hygromycin-B antibiotic (Table 4).
Biological activities of the antimicrobial agent: Data recorded in Table 5 indicated that the antimicrobial agent is fairly active against both, Gram positive and Gram negative bacteria and unicellular and filamentous fungi.
| Table 4: | A comparative study of characteristic properties of antimicrobial agent in relation to Reference antibiotic (Hygromycin-B) |
 | |
| Table 5: | Minimum inhibitory concentration of the antimicrobial agent |
 | |
The Minimum Inhibitory Concentration (MIC) of antibiotic produced by Streptomyces crystallinus, AZ151 was determined and results showed that MIC (μg mL-1) against Staphylococcus aureus (1.73), Klebsiella pneumonia (3.9), Escherichia coli and Salmonella typhi (7.8), for Aspergillus flavus and Alternaria alternata (31.25) and Saccharomyces cerevisiae (62.5) (Table 5).
DISCUSSION
The aminoglycosides such as Hygromycin B are a large and diverse class of antibiotics that characteristically contain two or more aminosugars linked by glycosidic bonds to an aminocyclitol component. The cyclitol is 2-deoxystreptamine-streptomycin which has a streptidine moiety (Umezawa and Hooper, 1982). Soil, in particular, is an intensively exploited ecological inhabitant, of which, produce many useful natural products, including clinically important antibiotics (Waksman, 1961; Waksman, 1975). Among soil inhabitants, actinomycetes and more specifically streptomycetes, are of practical importance because they produce most of the useful natural antibiotics for medical use. Nevertheless, selective isolation of soil actinomycetes is important for understanding their ecological properties and for finding novel strains which can produce useful bioactive secondary metabolites. Therefore, various media and techniques have been developed for selective isolation of actinomycetes (Hozzein et al., 2008). A total 6 actinomycetes were isolated from the soil sample through crowded plate technique were subjected to primary screening and identified as Intrasporangium sp., Dactyl sporangium sp., Micromonospora sp., Streptoverticillium sp. and two Streptomyces sp. (Raja et al., 2010).
In the course of searching for new actinomycetes producing antibiotics in our study, screening program has been conducted. The data presented in this study gave detail information on the isolation and identification of actinomycetes from various Egyptian localities e.g. (Assiut, Luxor and El-Minia governorates). The results indicated the highest percentage of isolation (43.2%), 84 isolates out of 194, were isolated on Starch Nitrate Agar (SNA) medium followed by (28.3%), 55 isolates on both starch casein agar and glycerol asparagine agar (data not shown).
The most active isolates, in terms of mean diameter of inhibition zone (mm), 74 (38%) was obtained against Staphylococcus aureus 90.5% (67) followed by Aternaria alternata 43.2% (32) and klebsiella pneumoniae, 41.8% (31) while the lowest percentage was obtained against Fusarium verticillioides 21.6% (16 isolates), Salmonella typhi 20.2% (15 isolates), Escherichia coli 9.4% (7 isolates), Aspergillus fumigatus 9.4% (7 isolates), Saccharomyces cerevisiae 8.1% (6 isolates) and Aspergillus flavus 6.7% (5 isolates).
Many other percentages were also found 100% (14) against Phytophthora megasperma, followed by Alternaria solani and Alternaria alternata 78.5% (11) while the lowest percentage was obtained against Fusarium solani and Saccharomyces cerevisiae 14.2% (2) and 7.1% (1), respectively (Aghighi et al., 2004). In addition, streptomycetes isolates appear to be highly active against Gram-positive bacteria (Hamdi et al., 1980; Hussein et al., 1981; Saadoun et al., 1998).
Among our strains (194), an actinomycete isolates (AZ151) showed high potencies against all microorganisms tested. Its morphological, physiological and biochemical characteristics as well as, its color and culture characteristics have been investigated. A similar study found that the highest percentage of active isolates was red and gray series and the lowest in the green and white ones (Saadoun et al., 1998). However, most antimicrobial producing species of streptomycetes were found in the gray and yellow series of no chromomeric type and no antibiotic produced by white and green series chromomeric type (Aria et al., 1976). Antimicrobial sensitivity testing of these strains was done by Kirby-Bauer disc diffusion method (Khan et al., 2011). Minimum Inhibitory Concentration (MIC) of the extracts was determined by the micro broth dilution against 14 clinical and standard strains of methicillin resistant and sensitive of Staphylococcus aureus (Dadgar et al., 2006). Over 110 soil actinomycetes isolates were screened among which one isolate showed high level of activity in Agar disk and Well diffusion methods against E. carotovora subsp. carotovora and identified as a new strain of Streptomyces plicatus (strain 101) (Zamanian et al., 2005).
The species belonging to the genus Streptomyces constitute 50% of the total population of soil actinomycetes and 75-80% of the commercially and medicinally useful antibiotics have been derived from this genus (Mellouli et al., 2003). Over 110 soil actinomycetes isolates were screened among which one isolate showed high level of activity in Agar disk and Well diffusion methods against E. carotovora subsp. carotovora and identified as a new strain of Streptomyces plicatus (strain 101) (Zamanian et al., 2005).
The extract from culture filtrate of endophytic Streptomyces sp. ST8 by ethyl acetate has activity against S. mutans ATCC25175 and 104B. The extract at such concentrations (0.05-5 mg mL-1) showed the inhibition of bacterial adherence on glass surfaces and saliva-coated hydroxyapatite. The crude extract also decreased the activity of glucosyltransferase and glucan-binding lectin from both strains (Taechowisan et al., 2008).
Besides, selective isolation of soil actinomycetes is important for understanding their ecological properties and for finding novel strains which can produce useful bioactive secondary metabolites. Therefore, various media and techniques have been developed for selective isolation of actinomycetes (Hozzein et al., 2008).
In this study, we found the nucleotide sequence of the 16S rRNA gene (1.5 kb) of the actinomycete isolate, AZ151 evidenced a 92% similarity with Streptomyces crystallinus 16S rRNA genes. In addition, multiple sequence alignment was conducted the sequences of the 16S rDNA gene of Streptomyces crystallinus, (CHR and SNG). The sequencing product was determined as 1141 bp2). On the basis of theses data and in view of the comparative study of the recorded properties of AZ151 in relation to the most closest reference strain, viz. Streptomyces crystallinus, it could be stated that it is suggestive of being belonging to Streptomyces crystallinus which can produce a broad-spectrum antibiotic.
In addition, 16S rRNA sequence data have proved invaluable in Streptomycetes systematics, in which they have been used to identify several newly isolated Streptomyces (Mehling et al., 1995). This finding is in agreement with that noticed of antibiotic phenazine derivatives and their formation pathways in a new Streptomyces strain P510, where culture characteristics and 16S rRNA nucleotide analysis confirmed strain P510 as Streptomyces griseoluteus (Wang et al., 2011).
Several methods have been developed to identify Streptomyces species. These include, culturing methods using the selective plating technique (Kuster and Williams, 1964), construction of genetic marker systems (Wipat et al., 1991), a combination of chemical markers, the presence of LL diaminopimelic acid and the absence of characteristic sugars in the cell wall (Lechevalier and Lechevalier, 1970; Atta et al., 2011). Also, sensitivity to antibiotics and phages, serological reactions and ecological properties has also been used for the classification of Streptomyces spp. (Shirling and Gottlieb, 1966; Lechevalier and Lechevalier, 1980).
The highest antimicrobial activity was achieved at optimum environmental and nutritional conditions in Streptomyces crystallinus AZ151 culture. Several studies have shown the optimization of nutritional and environmental conditions for antibiotic production (Kumar and Satyanarayana, 2007; Latifian et al., 2007; Gupta et al., 2008; Kagliwal et al., 2009; Atta et al., 2011). San et al. (2010) worked on two venoms of Calloselasma rhodostoma and Ophiophagus hannah and determined (MIC) with Staphylococcus aureus ATCC25923, ATCC29213 and ATCC43300. The MIC values obtained for Calloselasma rhodostoma were 125 μg mL-1 when tested against S. aureus ATCC25923 and ATCC43300 while it was 250 μg mL-1 when tested against S. aureus ATCC29213. MIC values obtained for Ophiophagus hannah were 250 μg mL-1 when tested against all three strains.
In fact, the active metabolites were extracted by ethyl acetate (Criswell et al., 2006; Sekiguchi et al., 2007; Augustine et al., 2005). The extract from culture filtrate of endophytic Streptomyces sp. ST8 by ethyl acetate has activity against S. mutans ATCC25175 and 104 B. The extract at such concentrations (0.05-5 mg mL-1) showed the inhibition of bacterial adherence on glass surfaces and saliva-coated hydroxyapatite. The crude extract also decreased the activity of glucosyltransferase and glucan-binding lectin from both strains (Taechowisan et al., 2008).
The organic phase was collected and evaporated under reduced pressure using rotary evaporator. Moreover, the purification process indicated that maximum activity was recorded between fraction Nos. 14-23. Many workers used a column chromatography packed with silica gel and the same situation was observed (Hitchens and Kell, 2003; Criswell et al., 2006; Sekiguchi et al., 2007).
CONCLUSION
In this study we determined the physico-chemical characteristics of the purified antimicrobial agents produced by Streptomyces crystallinus AZ151 and indicated that, it produces characteristic odour, its melting points is 160°C, infrared (IR) spectrum showed characteristic band corresponding to 21 peaks, a maximum absorption UV peak recorded at 225 NM and Mass spectrum confirmed that the molecular weight is at 432.36. Consequently, the elemental analytical data are find to be:
C = 46.43; H = 7.46; N = 6.81 and O = 39.3, indicate a suggested empirical formula of C15H30N2O10 and being belonging to Hygromycin-B antibiotic. Physico-chemical characteristics of many antibiotics were determined (Koshiyama et al., 1969; Singh and Gurusiddaiah, 1984; Omura et al., 1987; Uyeda et al., 2001; Yanai and Murakami, 2004; Atta et al., 2011).
REFERENCES
- Aghighi, S., G.H.S. Bonjar, R. Rawashdeh, S. Batayneh and I. Saadoun, 2004. First report of antifungal spectra of activity of Iranian actinomycetes strains against Alternaria solani, Alternaria alternate, Fusarium solani, Phytophthora megasperma, Verticillium dahliae and Saccharomyces cerevisiae. Asian J. Plant Sci., 3: 463-471.
CrossRefDirect Link - Atta, H.M., B.M. Haroun and M.A. Khalifa, 2011. Physico-chemical characteristics of vernamycin-A antibiotic biosynthesis by Streptomyces SP-AZ-SH-29. J. Saudi Chem. Soc., 15: 247-255.
CrossRefDirect Link - Augustine, S.K., S.P. Bhavsar and B.P. Kapadnis, 2005. A non-polyene antifungal antibiotic from Streptomyces albidoflavus PU 23. J. Biosci., 30: 201-211.
Direct Link - Becker, B., M.P. Lechevalier, R.E. Gordon and H.A. Lechevalier, 1964. Rapid differentiation between Nocardia and Streptomyces by paper chromatography of whole-cell hydrolysates. Applied Microbiol., 12: 421-423.
PubMed - Berdy, J., 1974. Recent developments of antibiotic research and classification of antibiotics according to chemical structure. Adv. Applied Microbiol., 18: 309-406.
PubMed - Chapman, G.H., 1952. A simple method for making multiple tests of a micro-organism. J. Bacteriol., 63: 147-149.
PubMed - Criswell, D., V.L. Tobiason, J.S. Lodmell and D.S. Samuels, 2006. Mutations conferring aminoglycoside and spectinomycin resistance in Borrelia burgdorferi. Antimicrob. Agents Chemother., 50: 445-452.
PubMed - Dadgar, T., M. Asmar, A. Saifi, M. Mazandarani and H. Bayat et al., 2006. Antibacterial activity of certain iranian medicinal plants against methicillin-resistant and sensitive Staphylococcus aureus. Asian J. Plant Sci., 5: 861-866.
CrossRefDirect Link - Dhananjeyan, V., N. Selvan and K. Dhanapal, 2010. Isolation, characterization, screening and antibiotic sensitivity of actinomycetes from locally (Near MCAS) collected soil samples. J. Biol. Sci., 10: 514-519.
CrossRefDirect Link - Del-Sol, R., I. Armstrong, C. Wright and P. Dyson, 2007. Characterization of changes to the cell surface during the life cycle of Streptomyces coelicolor: Atomic force microscopy of living cells. J. Bacteriol., 189: 2219-2225.
Direct Link - Edwards, U., T. Rogall, H. Blocker, M. Emde and E.C. Bottger, 1989. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucl. Acids Res., 17: 7843-7853.
CrossRefPubMedDirect Link - Esmaeili, P. and R. Moshiri, 2011. Geographical factors in medicine and human settlements. J. Applied Sci., 11: 212-218.
CrossRef - Gordon, R.E., 1966. Some criteria for the recognition of Nocardia madurae (vincent) blanchard. J.Gen. Microbiol., 45: 355-364.
CrossRefPubMedDirect Link - Gordon, R.E., D.A. Barnett, J.E. Handerhan and C.H.N. Pang, 1974. Nocardia coeliaca, Nocardia autotrophica and the Nocardin strain. Int. J. Syst. Evolut. Microbol., 24: 54-63.
CrossRef - Gupta, S., M. Kapoor, K.K. Sharma, L.M. Nair and R.C. Kuhad, 2008. Production and recovery of an alkaline exo-polygalacturonase from Bacillus subtilis RCK under solid-state fermentation using statistical approach. Biores. Technol., 99: 937-945.
Direct Link - Hall, T.A., 1999. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acid Symp. Ser., 41: 95-98.
Direct Link - Hamdi, Y.A., A.M. Al-Taj and A. Dewwdar, 1980. Genera and species of actinomycetes isolated from Iraqi soils. Egypt. J. Microbiol., 15: 6-21.
Direct Link - Hankin, L., M. Zucker and D.C. Sands, 1971. Improved solid medium for the detection and enumeration of pectolytic bacteria. Applied Microbiol., 22: 205-209.
Direct Link - Hindra, P.P. and M.A. Elliot, 2010. Regulation of a novel gene cluster involved in secondary metabolite production in Streptomyces coelicolor. J. Bacteriol., 192: 4973-4982.
CrossRef - Hitchens, G.D. and D.B. Kell, 2003. On the effects of thiocyanate and venturicidin on respiration-driven proton translocation in Paracoccus denitrificans. Biochim. Biophys. Acta, 766: 222-232.
PubMed - Hozzein, W.N., M.I.A. Ali and W. Rabie, 2008. A new preferential medium for enumeration and isolation of desert actinomycetes. World J. Microbiol. Biotechnol., 24: 1547-1552.
CrossRefDirect Link - Huddleston, A.S., N. Cresswell, M.C. Neves, J.E. Beringer, S. Baumberg, D.I. Thomas and E.M. Wellington, 1997. Molecular detection of streptomycin-producing streptomycetes in Brazilian soils. Applied Environ. Microbiol., 63: 1288-1297.
Direct Link - Jones, K.L., 1949. Fresh isolates of actinomycetes in which the presence of sporogenous aerial mycelia is a fluctuating characteristic. J. Bacteriol., 57: 141-145.
Direct Link - Kagliwal, L.D., S.A. Survase and R.S. Singhal, 2009. A novel medium for the production of cephamycin C by Nocardia lactamdurans using solid-state fermentation. Bioresour. Technol., 100: 2600-2606.
CrossRefPubMedDirect Link - Khan, F., I. Shukla, M. Rizvi, T. Mansoor and S.C. Sharma, 2011. Detection of biofilm formation in Staphylococcus aureus. Does it have a role in Treatment of MRSA Infections? Trends Med. Res., 6: 116-123.
CrossRefDirect Link - Khucharoenphaisan, K., U. Puangpetch, K. Puttaraksa and K. Sinma, 2011. Grouping of actinomycetes isolated from termites using biochemical character. J. Biol. Sci., 11: 314-319.
CrossRefDirect Link - Khucharoenphaisan, K., N. Sripairoj and K. Sinma, 2012. Isolation and identification of actinomycetes from termite's gut against human pathogen. Asian J. Anim. Vet. Adv., 7: 68-73.
CrossRefDirect Link - Koshiyama, H., H. Tsukiura, K. Fujisawa, M. Konishi and M. Hatori, 1969. Studies on cirramycin A1. I. Isolation and characterization of cirramycin A1. J. Antibiot., 22: 61-64.
PubMedDirect Link - Kuster, E. and S.T. Williams, 1964. Selection of media for isolation of Streptomycetes. Nature, 202: 928-929.
CrossRefDirect Link - Latifian, M., Z. Hamidi-Esfahani and M. Barzegar, 2007. Evaluation of culture conditions for cellulase production by two Trichoderma reesei mutants under solid-state fermentation conditions. Bioresour. Technol., 98: 3634-3637.
Direct Link - Lechevalier, M.P. and H. Lechevalier, 1970. Chemical composition as a criterion in the classification of aerobic actinomycetes. Int. J. Syst. Evol. Microbiol., 20: 435-443.
CrossRefDirect Link - Lee, E.J., N. Karoonuthaisiri, H.S. Kim, J.H. Park, C.J. Cha, C.M. Kao and J.H. Roe, 2005. A master regulator σB governs osmotic and oxidative response as well as differentiation via a network of sigma factors in Streptomyces coelicolor. Mol. Microbiol., 57: 1252-1264.
CrossRef - Mehling, A., U.F. Wehmeier and W. Piepersberg, 1995. Nucleotide sequences of Streptomyces 16S ribosomal DNA: Towards a specific identification system for Streptomycetes using PCR. Microbiology, 141: 2139-2147.
CrossRef - Mellouli, L., R.B. Ameur-Mehdi, S. Sioud, M. Salem and S. Bejar, 2003. Isolation, purification and partial characterization of antibacterial activities produced by a newly isolated Streptomyces sp. US24 strain. Res. Microbiol., 154: 345-352.
CrossRefDirect Link - Myles, D.C., 2003. Novel biologically active natural and unnatural products. Curr. Opin. Biotechnol., 14: 627-633.
PubMed - Omura, S., A. Nakagawa, T. Fujimoto, K. Saito, K. Otoguro and J.C. Walsh, 1987. Hygromycin A, an antitreponemal substance. I. Screening method and therapeutic effect for Treponema hyodysenteriae-caused infection in CF-1 mice. J. Antibiot., 40: 1619-1626.
PubMedDirect Link - Pridham, T.G. and D. Gottlieb, 1948. The utilization of carbon compounds by some Actinomycetales as an aid for species determination. J. Bacteriol., 56: 107-114.
Direct Link - Raja, A., P. Prabakaran and P. Gajalakshmi, 2010. Isolation and screening of antibiotic producing psychrophilic actinomycetes and its nature from rothang hill soil against viridans Streptococcus sp. Res. J. Microbiol., 5: 44-49.
CrossRefDirect Link - Raja, A. and P. Prabakarana, 2011. Actinomycetes and drug-An overview. Am. J. Drug Discovery Dev., 1: 75-84.
CrossRefDirect Link - Saadoun, I., M.J. Mohammad, F. Al-Momani and M. Meqdam, 1998. Diversity of soil streptomycetes in Northern Jordan. Actinomycetes, 9: 52-60.
Direct Link - San, T.M., J. Vejayan, K. Shanmugan and H. Ibrahim, 2010. Screening antimicrobial activity of venoms from snakes commonly found in Malaysia. J. Applied Sci., 10: 2328-2332.
CrossRefDirect Link - Sanger, F., S. Nicklen and A.R. Coulson, 1977. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. U.S.A., 74: 5463-5467.
CrossRefPubMedDirect Link - Sekiguchi, J.I., T. Miyoshi-Akiyama, E. Augustynowicz-Kopec, Z. Zwolska and F. Kirikae et al., 2007. Detection of multidrug resistance in Mycobacterium tuberculosis. J. Clin. Microbiol., 45: 179-192.
CrossRef - Shirling, E.B. and D. Gottlieb, 1966. Methods for characterization of Streptomyces species. Int. J. Syst. Evol. Microbiol., 16: 313-340.
CrossRefDirect Link - Singh, S.K. and S. Gurusiddaiah, 1984. Production, purification and characterization of chandramycin, a polypeptide antibiotic from Streptomyces lydicus. Antimicrob. Agents Chemother., 26: 394-400.
PubMedDirect Link - Taechowisan, T., A. Sitthipanya, A. Wanbanjob and P. Tantiwachwuttikul, 2008. Inhibitory effects of endophytic Streptomyces sp. ST8 on the growth, adherence and glucosyltransferase of Streptococcus mutans. J. Boil. Sci., 8: 43-51.
CrossRefDirect Link - Umezawa, H., 1977. Recent advances in bioactive microbial secondary metabolites. Jpn. J. Antibiot., 30: 138-163.
PubMed - Uyeda, M., M. Mizukami, K. Yokomizo and K. Suzuki, 2001. Pentalenolactone I and hygromycin A, immunosuppressants produced by Streptomyces filipinensis and Streptomyces hygroscopicus. Biosci. Biotechnol. Biochem., 65: 1252-1254.
PubMedDirect Link - Wang, Y., Q. Luo, X. Zhang and W. Wang, 2011. Isolation and purification of a modified phenazine, griseoluteic acid, produced by Streptomyce griseoluteus P510. Res. Microbiol., 162: 311-319.
PubMedDirect Link - Williams, S.T. and F.L. Davies, 1965. Use of antibiotics for selective isolation and enumeration of actinomycetes in soil. J. Gene. Microbiol., 38: 251-261.
CrossRefDirect Link - Wipat, A., E.M. Wellington and V.A. Saunders, 1991. Streptomyces marker plasmids for monitoring survival and spread of Streptomycetes in soil. Applied Environ. Microbiol., 57: 3322-3330.
PubMed - Yanai, K. and T. Murakami, 2004. The kanamycin biosynthetic gene cluster from Streptomyces kanamyceticus. J. Antibiot., 57: 351-354.
PubMed - Zamanian, S., G.H. Shahidi Bonjar and I. Saadoun, 2005. First report of antibacterial properties of a new strain of Streptomyces plicatus (strain 101) against Erwinia carotovora from Iran. Biotechnology, 4: 114-120.
CrossRefDirect Link - Nitsch, B. and H.J. Kutzner, 1969. Egg-yolk agar as a diagnostic medium for streptomycetes. Experientia, 25: 220-221.
CrossRefDirect Link