
H. Patel
Phytochemical, Pharmacological and Microbiological Laboratory, Department of Biosciences, Saurashtra University, Rajkot-360 005, Gujarat, India
Y. Vaghasiya
Department of Biochemistry, Saurashtra University, Rajkot-360 005, Gujarat, India
B.R.M. Vyas
Phytochemical, Pharmacological and Microbiological Laboratory, Department of Biosciences, Saurashtra University, Rajkot-360 005, Gujarat, India
S. Chanda
Phytochemical, Pharmacological and Microbiological Laboratory, Department of Biosciences, Saurashtra University, Rajkot-360 005, Gujarat, India
Bacteriology Journal
Year: 2012 | Volume: 2 | Issue: 2 | Page No.: 23-45







ABSTRACT
Staphylococcus aureus is a pathogen of major concern because of its ability to cause a diverse array of diseases ranging from minor infections to life threatening septicemia and its ability to adapt to adverse environmental conditions. Methicillin resistance among clinical isolates of S. aureus is still increasing. Knowledge of the risks and benefits associated with antibiotics makes the clinician better able to optimize the patient’s care. Numerous recent publications have documented the fact that despite growing problems with resistance to antimicrobial agents amongst important bacterial pathogens, the number of new antibiotics being brought to the market has shown a precipitous decline over the past several decades, with only a few antimicrobial agents are available for treatment of such infections and none of these possesses ideal characteristics. The rise in antibiotic-resistant microorganisms in recent years has led to an increasing search for new antibiotics.
PDF Abstract XML References Citation
Received: November 23, 2011;
Accepted: March 09, 2012;
Published: May 24, 2012
How to cite this article
H. Patel, Y. Vaghasiya, B.R.M. Vyas and S. Chanda, 2012. Antibiotic-resistant Staphylococcus aureus: A Challenge to Researchers and Clinicians. Bacteriology Journal, 2: 23-45.
URL: https://scialert.net/abstract/?doi=bj.2012.23.45
URL: https://scialert.net/abstract/?doi=bj.2012.23.45
INTRODUCTION
The emergence of resistance to antibiotics in Gram positive pathogens has become a major international problem (Patel et al., 2011). Antibiotic therapy is typically started before susceptibility information is available, but inappropriate initial therapy is associated with adverse clinical outcomes (Rodriguez-Bano et al., 2009). The pathogen S. aureus may use a variety of strategies to resist antibiotic therapy; these antibiotic-resistant strains are called Methicillin-Resistant Staphylococcus aureus (MRSA) (Khorvash et al., 2008). Infections caused by resistant pathogens are associated with higher morbidity and mortality than those caused by susceptible pathogens (Vaghasiya and Chanda, 2007). MRSA are relatively harmless and commonly found in the nose and on the skin of healthy people, where they can cause disease only when the body’s defense systems are compromised. MRSA infections are easily spread (Dadgar et al., 2006; Sheen, 2010) first detected in Europe in the 1960s. Today, MRSA is present in the hospitals of the majority countries and is usually resistant to a number of antibiotics. Clinical infections are mainly patients in hospital intensive care units, nursing homes and other chronic care facilities (Mertz et al., 2010), but MRSA strains are now being increasingly isolated from community-acquired infections as well. The increased incidence of community-associated MRSA infection has been associated with reports of increased morbidity and mortality specially, a longer duration of fever, a higher incidence of pulmonary complications along with bone and joint infections, prolonged hospitalization and the re-emergence of a severe staphylococcal sepsis syndrome (Gonzalez et al., 2005). S. aureus resistance is caused by expression of an altered penicillin-binding protein (PBP2a) and these PBP2a, as a transpeptidase, facilitates bacterial growth and cell wall synthesis at concentrations of β-lactams inhibitory to native penicillin-binding proteins (Klevens et al., 2007). PBP2a is chromosomally encoded within an externally acquired segment of DNA called mec DNA. This mec DNA is a large (approximately 30-50 kb) DNA fragments that does not occur in Methicillin-Sensitive S. aureus (MSSA) and is always located at a fixed site in the S. aureus chromosome, specially near the pur-nov-his gene cluster (Kuhl et al., 1978). mec DNA contains mecA, the structural gene for PBP2a; mecI and mecR1, regulatory elements controlling mecA transcription; and 20-45 kb of mec-associated DNA. The mec-associated DNA has been found to contain transposons and insertion elements providing a mechanism for the considerable variability found within the mec region. IS431 is a common insertion sequence in the staphylococcal plasmids and chromosome and is present within the mec DNA region. IS431 serves as a trap for resistance determinants with similar IS elements, accounting for the multiple drug resistance phenotype common in MRSA (Chambers, 1999). The mecA is present in all MRSA; there is considerable variation in the presence of the other genes (Archer et al., 1994). mecR1-mecI is present in 60 to 95% of mecA-positive S. aureus (Weller, 1999). Because mecI is such a strong repressor, it has been concluded that phenotypically resistant mecA positive S. aureus strains either do not possess mecI, or have mutations within mecI which prevent it from functioning (Kobayashi et al., 1998). Inactivation of mecI, by either mutation or deletion, is an important step in the production of PBP2a and expression of methicillin-resistance (Schentag et al., 1998).
The purpose of this review was to give a path to researchers and clinicians for awareness about multi-drug resistant S. aureus and its alternative treatment is in high demand.
STRUCTURE OF Staphylococcus aureus
The cell walls of gram-positive bacteria exhibit a wide diversity from simple to very complex structures (Fig. 1).
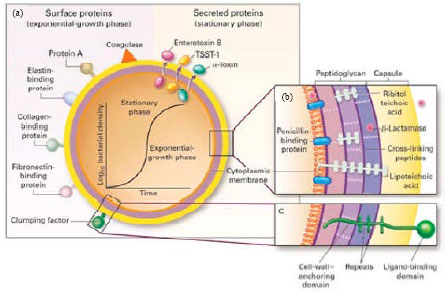 | |
| Fig. 1(a-b): | Structure of S. aureus (Lowy, 1998) |
Staphylococcal cell walls have a rather extraordinary type of structural design and belong to the most highly cross-linked type but the walls of other gram-positive bacteria exhibit a much lower degree of cross-linking and the muropeptide fraction of these walls does not contain long oligomeric chains. For example, the walls of bacilli exhibit about 50-55% cross-linking and the most important component of the oligopeptide fraction is trimeric and tetrameric muropeptides (Gally et al., 1991). The cell wall envelope functions as a physical barrier that protects bacteria from their environment and as a rigid exoskeletal element that prevents bacterial rupture in low osmolar environments such as host tissues (Sjoquist et al., 1972). The cell wall of the microorganisms plays an important role in the susceptibility to infections and pathogenicity (Van Heijenoort and Gutmann, 2000).
The cell wall of S. aureus is structurally similar to that of Group A streptococci: both are composed of murein (Tomasz, 2000), teichoic acids (Baddiley, 1989) and wall-associated surface proteins (Mazmanian et al., 2001). Murein consists of glycan strands that are cross-linked by peptide bridges supplying the structural integrity of the sacculus. It is a distinctive feature of staphylococci that the observed degree of murein cross-linking which was determined as a ratio of bridged peptides to the total amount of all peptide ends in general, is extremely high, of the order of 80-90% (Gally and Archibald, 1993). The carbohydrate antigen is a teichoic acid which in S. aureus is a polymer of N-acetylglucosamine and polyribitol phosphate. Antibodies of teichoic acid can be detected in normal human serum and elevated antibody titers are present in patients with deep-seated staphylococcal infections. Teichoic acid has no established role in virulence and antibodies of this carbohydrate are not protective. The protein component of the cell wall includes protein A which reacts with IgG of normal human serum (Lowy, 1998) and it can be released from the bacterial surface by treatment of staphylococci with lysostaphin, a glycylglycine endopeptidase that cleaves the pentaglycyl cross-bridge of the cell wall (Sjoquist et al., 1972). Lysozyme, an N-acetylmuramidase that cuts the glycan strands (Hash and Rothlauf, 1967, releases protein A molecules as a spectrum of fragments with varying masses due to the presence of linked peptidoglycan fragments of different sizes (Navarre et al., 1998). Although, bacterial peptidoglycan structure varies from one species to another, several structural or functional elements are conserved (Schleifer and Kandler, 1972). The glycan strands of all bacterial peptidoglycan consist of repeat disaccharide units, N-acetylglucosamine-(β1-4)-N-acetylmuramic acid (GlcNAc-MurNAc) (Ghuysen and Strominger, 1963). Glycan chains are cross-linked by short cell wall peptides and generate a three-dimensional molecular network that maintains the integrity of the bacterium (Tipper and Strominger, 1965). Finally, penicillin binding proteins catalyze the polymerization of lipid II subunits via trans-glycosylation and trans-peptidation reactions, thus generating the cross-linked peptidoglycan that constitutes the main component of the bacterial cell wall (Perry et al., 2002).
VIRULENCE FACTORS
Staphylococcus aureus is a pathogen expressing multiple virulence factors that mediate host colonization, invasion of damaged skin and mucosa, dissemination through the body and evasion of host defense mechanisms (Ferry et al., 2005; Chanda et al., 2010). The pathogenicity and virulence of S. aureus infections is associated to various bacterial surface components (e.g., capsular polysaccharide and protein A) including those recognizing adhesive matrix molecules (e.g., clumping factor (clf), Fibronectin Binding Protein (FBN) and to extracellular proteins (e.g., coagulase, hemolysins, enterotoxins, Toxic-Shock Syndrome (TSS) toxin, exfoliatins toxin and Panton-Valentine leukocidin (Labandeira-Rey et al., 2007). Virulence factors can generally be separated into three based on their function: the adhesins, the toxins and the immune-modulators. The adhesins are surface-attached proteins that allow the bacteria to attach to a wide variety of human tissues. The toxins are secreted proteins that cause tissue damage and generate pus in abscesses which is believed to facilitate transmission between hosts. The immune-modulators are proteins that interfere with host immunity preventing defense against infections (Collins et al., 2010). Some microbial surface proteins mediate the adherence of S. aureus to host proteins, such as fibrinogen and fibronectin. These plasma proteins coat indwelling medical devices and the ability of the bacteria to adhere to the deposited proteins is believed to be an important factor in the pathogenesis of wound and foreign body infections (Foster and Hook, 1998). In S. aureus, the adhesin genes which include clf (Ni Eidhin et al., 1998) and fnb (Jonsson et al., 1991) that encode the fibrinogen- and the fibronectin-binding proteins, respectively. Fibronectin-Binding Protein (FnBP) A and Fibronectin-Binding Protein (FnBP) B, encoded by the fnbA and fnbB genes, respectively, play prominent roles in S. aureus colonization and attachment of host tissues or implanted biomaterials (Greene et al., 1995). FnBPs also promote endocytic uptake of S. aureus by epithelial and endothelial cell lines and fibroblasts (Fowler et al., 2000).
DIFFERENCES BETWEEN HA-MRSA AND CA-MRSA
Several differences have been noted in HA-MRSA and CA-MRSA infections. HA-MRSA isolates are typically resistant to multiple non-β-lactam antimicrobials. However, CA-MRSA isolates are usually susceptible to numerous non-β-lactam antibiotics including trimethoprim-sulfamethoxazole, tetracyclines and clindamycin (Weber, 2005).HA-MRSA infections represent a burden for both patients and health care systems, because of their association with high morbidity and mortality and increased hospitalization costs (Cosgrove et al., 2005). And these problems arose firstly in large tertiary care hospitals with patients in burns (Kassis et al., 2011), post-operative (Jeannon et al., 2010),prolonged hospitalization (Simo and French, 2006) and intensive care wards (Karas et al., 2009).Increased risk of MRSA infection was associated with use of multiple broad spectrum antibiotics (Bradley, 1992), indwelling devices (Rimland, 1985), ventilatory support (Craven et al., 1981), severity of underlying disease (Agostino et al., 2010) and length of hospital stay (Elliott et al., 2010). Nosocomially-acquired MRSA isolates tended to be multidrug resistant; community-acquired MRSA strains obtained from patients without identified risk tended to be resistant only to methicillin. This more restricted set of antibiotic resistances has also been observed in studies of community-acquired MRSA strains among intravenous drug abusers compared with nosocomially acquired MRSA isolates (Herold et al., 1998). Methicillin and β-lactam resistance in HA-MRSA and CA-MRSA is mediated by the altered penicillin binding protein 2a. The mec gene facilitates production of “penicillin-binding protein 2a” and is carried on a mobile genetic element, staphylococcal cassette chromosome (SCCmec). PBP2a make stronger the cell wall and increases resistance to β-lactam antibiotics by blocking the β-lactam binding site (Ito et al., 2001). The most common hospital-acquired MRSA organisms contain SCCmec I, II and III, genetic elements that encode resistance to several antibiotics in addition to β-lactams. Type I contains no additional resistance determinants, but types II and III contain resistance determinants in addition to mecA; these additional genetic elements account for the antimicrobial resistance to numerous antibiotics in addition to the β-lactam agents. The three SCCmec types contained in HA-MRSA have an identical chromosomal integration site and cassette chromosome recombinase genes which are responsible for horizontal transfer of SCCmec (Daum et al., 2002). Thus, HA-MRSA is resistant to loads of different antibiotics and has a discriminating advantage as they are spread among patients by hands of personnel and contaminated environmental surfaces. The presence of underlying diseases and multiple types of instrumentation and procedures predisposes patients to colonization and infection by the multiple drug resistant strains of HA-MRSA. Type IV is now found in CA-MRSA and predominates in individuals without hospital-associated risk factors. SCCmec type IV has greater sensitivity to non-β-lactam antimicrobial agents (Baba et al., 2002) but are considered highly virulent because of the high prevalence of Panton-Valentine Leukocidin (PVL) producing strains. PVL has been found predominantly in SCCmec type IV and VCA-MRSA and it is clinically responsible for the excessive amount of leukocyte obliteration leading to large amounts of pus (Farley, 2008). The particular pattern of virulence and disease presentation of CA-MRSA has been linked to PVL a phage-borne toxin that has been associated with necrotic skin disease and pneumonia (Campbell et al., 2008).
S. aureus INFECTIONS: FROM HARMLESS TO LIFE-THREATENING
S. aureus causes a wide variety of infections, most of which are localized to the skin and are nonfatal. The bacterium produces many superficial skin and soft infections such as hair, impetigo, follicles (Siddiqi et al., 2002; Hisata et al., 2011). It also causes boils which are deeper pus-filled abscesses of the skin and underlying tissue. Colonization of the anterior nares with S. aureus has been shown to be a risk factor for invasive infection. It is the second most important cause of hospital-acquired pneumonia. It can cause meningitis usually as a result of infection after brain surgery or as a consequence of a S. aureus infection in the blood. S. aureus also causes a painful infection of joint fluid known as septic or infective arthritis. Most serious of all are the deep-seated infections such as osteomyelitis and an infection of the heart valves called endocarditis and toxin-mediated diseases such as gastroenteritis, staphylococcal scalded-skin syndrome (Yoke-Kqueen et al., 2006; Miller and Kaplan, 2009; Anam et al., 2010). Osteomyelitis is a highly debilitating condition resulting in significant morbidity and health care costs and it is disreputably difficult to treat (Tuzuner-Oncul et al., 2009). S. aureus is the most common cause of these surgical wound infections. Deeper wound infections are much more serious and almost always need additional surgery to remove infected tissue. These bloodstream infections often occur in patients who have a surgical wound or are receiving intravenous (IV) medications or supplements, in people undergoing dialysis for kidney failure, in diabetic and in IV drug users (Lodise et al., 2003).
CLINICAL ASPECTS AND EPIDEMIOLOGY
Methicillin-resistant S. aureus is resistant to many antibiotics that become a major clinical problem worldwide. The epidemiology of staphylococcal infections is beginning to modify again; strains generally associated with CA-MRSA are now showing up in nosocomial infections (Maree et al., 2007) and some health care-related strains are being acquired in the community. S. aureus has been a leading cause of human infections throughout history. From 1997-1999, S. aureus was reported as the most abundant cause of skin and soft tissue, bloodstream and lower respiratory tract infections in the United States, Canada, Europe, Latin America and the Western Pacific Coast (Diekema et al., 2001). The incidence of S. aureus associated infections has increased dramatically since the emergence of methicillin resistant strains and high rates of mortality and morbidity are occurring world-wide (Oliveira et al., 2002). Combined with the increasing problem of multiple antibiotic resistances, these numbers underline the high social and economic burden of this fastidious pathogen.
ANTIBIOTIC MISUSE AND THE EMERGENCE OF RESISTANCE
The discovery and development of antibiotics are among the most powerful and successful achievements of modern science and that has helped dramatically in curing patients suffering from bacterial infections (Nair and Chanda, 2005). The appropriate use of antibiotics is one of the most essential weapons against disease. Hence, interventions need to target inappropriate patterns of use, specifically those that have contributed most significantly to the development of resistance (Lubelchek and Weinstein, 2008).
ANTIBIOTICS
The propensity for antibiotic use to support the emergence of resistant pathogens is called antibiotic pressure and there are numeral reports of resistance rising during increased antibiotic use and falling after a reduction in use (McGowan, 1983). Over the past 6 decades, bacterial populations have responded to the discriminating pressure of antimicrobial drugs by evolving resistance to all commercially obtainable antimicrobial agents (Levin, 2001). Decreased discovery rates of novel classes of antimicrobial agents have substantiated a concept that for some bacterial species, we might face clinical infections for which there are no treatment options (Projan and Bradford, 2007). Treatment options for both inpatient management of severe infection and outpatient management of mild soft tissue infection are limited because of increasing rate of antimicrobial resistance (Johnson and Decker, 2008) and this antimicrobial resistance has a significant negative impact on the outcome of therapy and increases the risk factor of cross-infection in hospitals. Resistance leads to inappropriate empirical therapy, delay in starting effective treatment and the use of less effective, more expensive and more toxic drugs (French, 2005). MRSA is at present one of the most commonly identified antibiotic-resistant pathogens in many parts of the world, including Europe, the Americas, the Middle East, North Africa and East Asia (Diekema et al., 2001). The emergence of MRSA organisms with reduced susceptibility to a number of antibiotics is a serious and ongoing concern. MRSA will continue to evolve, hence the absolute necessity to control it before it really does get out of hand. Therefore, action must be taken to reduce this problem. The ultimate goal is to offer suitable and efficient antimicrobial drugs to the patient.
β-lactam group: Under increased discriminating pressure, S. aureus developed multiple mechanisms of resistance to modified penicillins, including methicillin (Fig. 2).
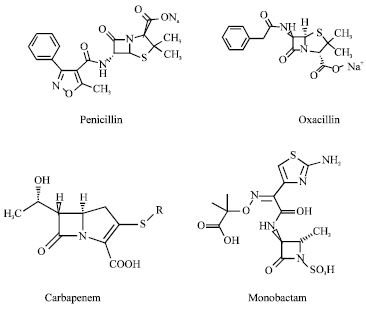 | |
| Fig. 2: | Structure of β-lactams |
Although, methicillin is resistant to hydrolysis by small quantities of staphylococcal β-lactamase, strains of S. aureus have been isolated that are competent of producing large amount of β-lactamase (Lakshmi et al., 2011). These hyperproducers of β-lactamase tend to resist methicillin through limited hydrolysis of the antibiotic, resulting in a phenotype that with deference to methicillin is intermediate between susceptible and resistant (McDougal and Thornsberry, 1986). A second mechanism of low-level resistance to methicillin by S. aureus strains involves the production of altered forms of native PBPs. S. aureus expresses at least four different PBPs, designated PBP1, 2, 3 and 4 that are the targets of β-lactam antibiotics (Hiramatsu, 1995; Jasmine et al., 2007). PBPs are essential proteins that are catalyze the trans-peptidation reaction and attached to the cytoplasmic membrane that cross-links the peptidoglycan of the bacterial cell wall; therefore the binding of β-lactam antibiotics to PBPs leads to a lethal event. Low level resistance to β-lactam antibiotics can be due to either a decrease in the binding affinities of PBPs for penicillins, or an increase in the production of PBPs, or both. Isolates containing the PBP2a-mediated resistance mechanism are clinically resistant to all available β-lactams, including penicillins, β-lactam/β-lactamase inhibitor combinations, cephalosporins, carbapenems and monobactams (Fasola and Peterson, 1992). PBP2a is encoded by the mecA gene which is not present in methicillin susceptible strains and it is believed to have been acquired from a distantly connected species, although the exact origin has not been found yet (Hiramatsu, 2001).
β-lactam-β-lactamase inhibitor combinations: The β-lactam-β-lactamase inhibitor combinations (amoxicillin-clavulanate, only available in oral formulation in the United States; ampicillin-sulbactam [IV]; ticarcillin-clavulanate [IV] and piperacillin-tazobactam [IV]) all have good activity against MSSA, but not MRSA and are active against anaerobes and gram-negative bacilli to varying degrees, making them appropriate choices for the treatment of polymicrobial infections including MSSA, such as complicated SSTI (Chambers, 2005).
Clindamycin: The lincosamide class of clindamycin an antibiotic commonly used in the treatment of skin infections. Clindamycin is active agent against gram-positive cocci, including MRSA. They are primarily used to treat less severe skin infections associated with MRSA that are identified in the community (Fig. 3). Lincosamides bind to the 23S ribosomal RNA in the 50S subunit of the ribosome. This binding inhibits the translocation of RNA during protein synthesis and blocks initiation of polypeptide formation. Lincosamides are bacteriostatic but can be bactericidal against highly susceptible bacteria (Le and Lieberman, 2006). As compared with the macrolides, clindamycin is more widely used to treat staphylococcal infections. It has specific use in the treatment of patients with hypersensitivity to penicillins in the community and is also used to treat serious infections because it blocks protein production. Many coagulase-negative staphylococci, particularly nosocomial isolates, are resistant to clindamycin antibiotic (Biedenbach et al., 2007).
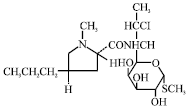 | |
| Fig. 3: | Structure of clindamycin |
The most common adverse reaction caused by clindamycin involves mainly the gastrointestinal system. Antibiotic-associated diarrhea is not uncommon in patients receiving clindamycin and in a subset of those patients is caused by the overgrowth of toxogenic C. difficile, resulting in antibiotic-associated colitis with pseudo-membrane formation. Clindamycin causes comparatively few significant drug interactions, but may potentiate neuromuscular blocking agents and may reduce the levels of cyclosporine (Thurnheer et al., 1999). Clindamycin may be administered orally, intramuscularly or parenterally (Zuckerman, 2004). The dosing of clindamycin for CA-MRSA in children is dependent on the severity of the infection. For mild infections like cellulitis, the dosing is 30 mg/kg/day by mouth, every 6-8 h. For severe infections such as osteomyelitis, the dosing is 40 mg/kg/day, administered intravenously and divided every 6-8 h (Martinez-Aguilar et al., 2003). Common adverse effects of clindamycin include nausea and abdominal pain. The standard oral preparation is clindamycin hydrochloride which may be given in doses ranging from 150-450 mg every 6 h. Another oral preparation of clindamycin is a palmitate ester suspension. Oral preparations are absorbed fast and have approximately 90% bioavailability. For intravenous delivery, 600-900 mg of clindamycin phosphate is infused every 6-8 h. This preparation may also be administered intramuscularly in doses as high as 600 mg. This drug penetrates into most tissues and fluids except cerebrospinal fluid (Enoch et al., 2009). SSTI caused by S. aureus are often treatable with clindamycin. Because of its activity against predominant clones of CA-MRSA in some geographic locales, clindamycin has been used for empiric and directed treatment of SSTI (Rahbar and Hajia, 2007), because its cell-wall active agent, to stop toxin production in patients with infections caused by toxin-producing strain of S. aureus (Stryjewski and Chambers, 2008). Detection of clindamycin resistance is very important in CA-MRSA because clindamycin is one of the antibiotics recommended to treat CA-MRSA infections, particularly in pediatric infections (Katopodis et al., 2010).
Fluoroquinolones: Fluoroquinolone group of antibiotics are widely used for the treatment of various infections and have efficient oral absorption, good tissue distribution and a broad range of activities against aerobic pathogens (Shrivastava et al., 2009). Fluoroquinolones are bactericidal agents that inhibit bacterial DNA gyrase (A and gyrB) and topoisomerase IV (C and parE) (Truong et al., 1997). Resistance is usually related with point mutations in the gyr or par loci (Fig. 4). Side effects that may be associated with this class include gastrointestinal distress, abnormalities in liver enzymes, cardiac conduction disturbances, hyperglycemia or hypoglycemia and drug rashes (Hooper and Wolfson, 1991). Clinically available fluoroquinolones include norfloxacin, ofloxacin, ciprofloxacin, levofloxacin, moxifloxacin and gemifloxacin. Ofloxacin is one of the most effective and quinolones which is used in current clinical practice than comparative drug such as ciprofloxacin resistance has been demonstrated among clinical S. aureus, particularly in MRSA (Blumberg et al., 1991; Shrivastava et al., 2009).
Trimethoprim/sulfamethoxazole: Trimethoprim/Sulfamethoxazole (TMP/SMX) is a combination of trimethoprim (Fig. 5), a diaminopyrimidine and sulfamethoxazole (Fig. 6), a sulfonamide. It is widely used as low-cost antibacterial agent for the treatment of community-acquired non-serious MRSA infections as well as other infections such as urinary tract infections. It is also used in combination with agents such as rifampin and the topical antibiotic mupirocin to eradicate MRSA colonization in patients. TMP/SMX is bactericidal agent against S. aureus and inhibits bacterial replication (Kaka et al., 2006).
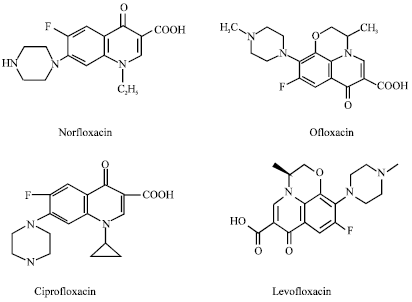 | |
| Fig. 4: | Structure of fluoroquinolones |
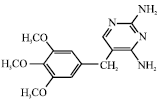 | |
| Fig. 5: | Structure of trimethoprim |
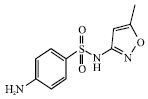 | |
| Fig. 6: | Structure of sulfamethoxazole |
Clinicians have used TMP/SMX in the past as an alternative to penicillins or vancomycin in the treatment of S. aureus infection and colonization. Although, TMP-SMX has demonstrated fair clinical efficacy against MRSA, it is not commonly used to treat serious MRSA infections. TMP/SMX is well tolerated, but sulfonamides can create a variety of untoward effects that are due partly to allergy and partly to direct toxicity.
Rifampin: Rifamycins inhibit only transcription and subsequent translation to proteins (Fig. 7). Their mechanism of antibacterial action involves the inhibition of a single target enzyme the β-subunit of RNA polymerase (Wehrli, 1983). It binds to the RNA polymerase and prevents synthesis of RNA. Rifampicin is used primarily as adjunctive (i.e., combination) therapy in difficult-to-treat staphylococcal infections, including MRSA infections. Because it is highly bio-available, rifampin can be administered orally. However, this antibiotic is rarely used as a single agent to treat CA-MRSA because rapid resistance can emerge among S. aureus on exposure to this drug.
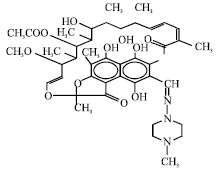 | |
| Fig. 7: | Structure of rifampin |
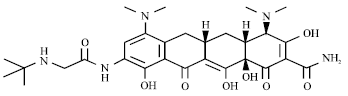 | |
| Fig. 8: | Structure of tigecycline |
It is used for synergy with other antibiotics like fusidic acid (Jensen, 1968). Rifampin has been shown to be highly effective in eradicating staphylococcal infections related with foreign bodies (Widmer et al., 1990). Resistance to rifampin occurs by mutations in the rpoB gene and single mutations in S. aureus selected in vitro and in vivo have been associated with both low and high levels of resistance depending on the nature of the amino acid change (Wichelhaus et al., 1999). Some clinical isolates of both S. aureus and S. pneumoniae have been found to carry multiple mutations in the cluster regions (Padayachee and Klugman, 1999). Common adverse effects of this antibiotic include vomiting, anorexia and abnormal pain. Rifampin can cause discoloration of body fluids, such as urine, sweat, saliva and tears. A rare but possible serious adverse effect associated with rifampicin is hepatotoxicity.
Tigecycline: Tigecycline is the first clinically available drug in a new class of antibiotics called the glycylcyclines which are structurally parallel to the tetracyclines in that they contain a central four-ring carbocyclic skeleton (Fig. 8). Tigecycline antibiotic has a substitution at the D-9 position which is believed to confer broad spectrum activity (Entenza and Moreillon, 2009). Tigecycline targets the bacterial ribosome and is a bacteriostatic agent. Tigecycline is effective against highly resistant gram-positive bacteria, including methicillin-resistant S. aureus and penicillin-resistant S. pneumoniae, as well as having activity against a variety of gram-negative bacteria, including those possessing extended-spectrum β-lactamase plasmids (Livermore, 2005).
Vancomycin: More than 95% of patients with S. aureus infections worldwide do not respond to first-line antibiotics such as penicillin or ampicillin (Rubin et al., 1999). Many multi-resistant MRSA strains are currently only susceptible to a single class of clinically available bactericidal antibiotic, the glycopeptides such as vancomycin (Fig. 9). However, if resistance to these agents emerges, some staphylococcal infections could be untreatable (Sieradzki et al., 1999).
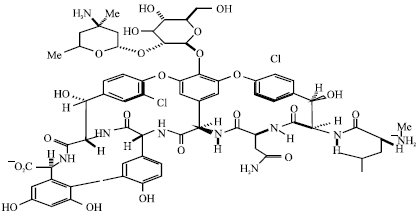 | |
| Fig. 9: | Structure of vancomycin |
Vancomycin was first discovered in 1956 in some soil samples from Southeast Asia, where it was produced by an actinomycete, S. orientalis (Rolston et al., 1990). It is not absorbed from the gastrointestinal tract in clinically relevant concentrations and so it must be used as a parenteral agent for systemic infections (Tsuji et al., 2007). It binds to the C-terminal end of late peptidoglycan precursors, preventing the effective formation of a bacterial cell wall (Courvalin, 2006). Its actions against bacteria are carried out on the outer surface of the bacterial cell membrane, as it cannot penetrate into the cytoplasm. Thus, vancomycin depends upon the bacterial translocation of the cell wall precursors onto the outer surface of the microbial membrane. Because vancomycin is effective against cell wall synthesis, it is active only against gram-positive organisms. Gram-positive organisms stain as they do because of the presence of the peptidoglycan cell wall. Gram-negative organisms do not have such a cell wall and are thus unaffected by vancomycin (Reed, 2007). Vancomycin remains the standard treatment for serious MRSA infections and serious MSSA infections in patients with β-lactam allergies (Khorvash et al., 2008). Its label notes the effectiveness of vancomycin in the treatment of staphylococcal endocarditis and other indications including bone infections, septicemia, skin and skin structure infections and lower respiratory tract infections. The most commonly seen adverse effect of this antibiotic is “Red Man Syndrome.” Patients with this syndrome can present with flushing, red neck, pruritus and rash involving the majority of the body (Savignon-Marinho et al., 2011). “Red Man Syndrome” is caused by an infusion-related release of histamine. Additional possible adverse effects related with vancomycin include nephrotoxicity and ototoxicity. Nephrotoxicity is not that common unless vancomycin is co-administered with other nephrotoxic agents such as gentamicin. Ototoxicity may occur only when the serum concentration of the antibiotic is extremely high. Even though vancomycin is a potent antibiotic against CA-MRSA, more and more resistance has surfaced. In 1996, the first case of Vancomycin-Intermediate S. aureus (VISA) was reported (Ward et al., 2001). After more than 40 years of clinical use, vancomycin-Resistant S. aureus (VRSA) was first identified in Detroit, Michigan in 2002, mediated by the vanA gene complex acquired from vancomycin-resistant enterococci (Centers for Disease Control and Prevention, 2002) its mechanism relies on inhibiting cell wall synthesis. VISA exhibits thickened peptidoglycan cell wall structures, differing from VRSA (Sakoulas and Moellering, 2008).
Linezolid: Linezolid is a synthetic oxazolidinone class of antimicrobial agent that binds to the ribosome (Fig. 10) and inhibits microbial protein synthesis (Champney and Miller, 2002).
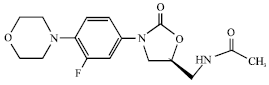 | |
| Fig. 10: | Structure of linezolid |
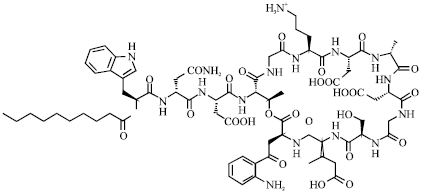 | |
| Fig. 11: | Structure of daptomycin |
The antibiotic reversibly blocks the formation of protein synthesis initiation complexes by binding to the 23S ribosomal RNA of the 50S ribosomal subunit, near the interface formed with the 30S ribosomal subunit (Hutchinson, 2003). In vitro studies have confirmed that linezolid has good activity against most medically important Gram-positive bacteria, including activity against MRSA and S. aureus with intermediate resistance to glycopeptides (Mutnick et al., 2002) and achieves better tissue penetration than vancomycin, but is bacteriostatic rather than bactericidal. Linezolid is approved in Europe and the USA for the treatment of complicated Skin and Skin-Structure Infections (SSSIs) as well as hospital-acquired and community-acquired pneumonia (Beibei et al., 2010). Linezolid is an extremely useful but expensive anti-staphylococcal agent. It remains active against most S. aureus isolates because of its excellent bio-availability, the option of oral treatment is very appealing and can diminish hospital length of stay (Falagas et al., 2008). Headache, nausea and thrombocytopenia are the main side effects, the latter usually occurring about two weeks into therapy. Disadvantages include expense, hematologic side effects and potential for resistance among S. aureus strains (Peeters and Sarria, 2005).
Daptomycin: Daptomycin is a cyclic lipopeptide group in clinical use and approved for the treatment of complicated skin and skin structure infections and right-sided endocarditis. This agent was developed in the early 1980s, but was initially abandoned because of concerns about skeletal muscle toxicity (Fig. 11). Daptomycin is active in vitro against staphylococci and other gram-positive bacteria (Eisenstein, 2004). It offers enhanced activity against resistant hospital pathogens such as MRSA. Its mechanism of action suggests that daptomycin causes a calcium-dependent rupture of the bacterial cell membrane, resulting in a net efflux of potassium that inhibits DNA, RNA and protein synthesis. Daptomycin has rapid bactericidal activity without cell lysis, a feature that could reduce the release of bacterial molecules and lessen the inflammatory response (Fenton et al., 2004). It is effective at all growth phases, including the stationary phase.
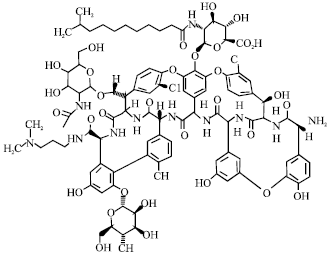 | |
| Fig. 12: | Structure of dalbavancin |
This property may be particularly useful in the treatment of indolent, deep-seated infections, such as osteomyelitis and endocarditis, in which bacteria expend a substantial amount of time in the stationary phase (Mascio et al., 2007). Daptomycin is obtainable only for intravenous administration and is highly protein bound (92%), with a half-life of approximately 8 h, allowing once-daily dosing. It has excellent efficacy against bacteria resistant to methicillin, vancomycin and linezolid. It is comparable to vancomycin for S. aureus bacteremia, including that related with right-sided endocarditis (Fowler et al., 2006). Resistance to daptomycin is uncommon but can be induced by serial passage in increasing concentrations of the antimicrobial. Clinically, daptomycin has occurred in patients who have received prolonged treatment (Skiest, 2006).
Dalbavancin: In Fig. 12, dalbavancin (BI397) is a second-generation lipoglycopeptide group antibiotic with unique pharmacokinetic properties that allow dosing once weekly that will be evaluated by the FDA in the near future for the treatment of resistant gram-positive infections (Van Bambeke et al., 2004). Second-generation dalbavancin is a semi-synthetic derivative of the teicoplanin-related glycopeptide A40926 modified with an amide appendage at the C-terminus and an alteration of the hydrophobic acylglucosamine substituent and like teicoplanin is active against VanB enterococci as well as the staphylococci and other important species (Lopez et al., 2005). Dalbavancin has excellent activity against methicillin-resistant staphylococcus aureus but not against vancomycin-resistant enterococci (Jones et al., 2006). The dosage of dalbavancin is 1000 mg, intravenously initially and 500 mg 7 days later. Dalbavancin is more potent than oritavancin. Infected patients who received weekly dalbavancin had an overall success rate that was significantly higher than that of those who received other antibiotic like vancomycin (Raad et al., 2005). Clinical trials in cSSTIs suggest that dalbavancin is as effective agent as compare to linezolid. So it is a promising new antimicrobial agent for the treatment of cSSTIs (Seltzer et al., 2003).
Telavancin: Telavancin (TD-6424) is another in the line of second generation semisynthetic lipoglycopeptide group antibiotic that has a double mechanism of action (Fig. 13). First, it inhibits peptidoglycan chain formation, blocking both trans-glycosylation and trans-peptidation. Second, telavancin alters membrane potential and increases cellular permeability (Leonard and Rybak, 2008). Telavancin has a high proportion of protein binding (93%), a high volume of tissue distribution and a half-life of 7-9 h (Barrett, 2005).
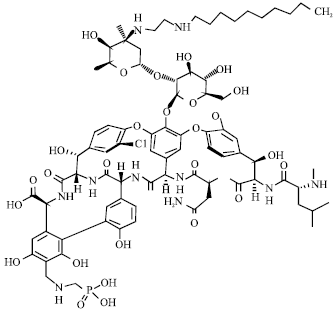 | |
| Fig. 13: | Structure of telavancin |
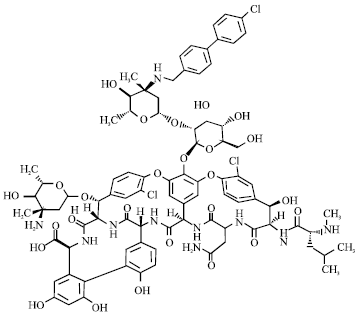 | |
| Fig. 14: | Structure of oritavancin |
An interesting and potentially important finding was that telavancin was active in an in vitro biofilm model where vancomycin and a number of other antibiotics were much less effective (Gander et al., 2005). Telavancin is rapidly bactericidal against staphylococci including VISA and glycopeptide tolerant strains and is bactericidal for enterococci in contrast to vancomycin (Rubinstein et al., 2011). The drug will be available only in parenteral form, probably for once daily administration.
Oritavancin: Oritavancin (LY333328) is another second-generation glycopeptides group antibiotic (Mercier and Hrebickova, 2005). It inhibits peptidoglycan biosynthesis at the same site as vancomycin (transglycosylation), but also forms dimers with higher affinity and at lower concentrations than vancomycin (Fig. 14). The drug may also act as an inhibitor of the transglycosylase enzyme.
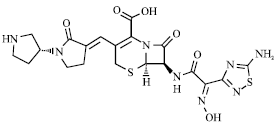 | |
| Fig. 15: | Structure of ceftobiprole |
The activity of oritavancin against vancomycin-resistant organisms is owing to its ability to dimerize, allowing more favorable interactions with the bacterial peptidoglycan chain. In addition, it is inhibits cell wall formation by blocking the transglycosylation step in peptidoglycan biosynthesis (Allen and Nicas, 2003). Oritavancin has a spectrum of in vitro activity parallel to that of vancomycin, but is active against A, B and VanC Enterococcus and is completely active against MRSA, including some VRSA strains (Poulakou and Giamarellou, 2008). Oritavancin is available for intravenously and is excreted slowly in urine and faeces. It accumulates in tissues and macrophages and has a extremely long terminal half-life that permits once-daily dosing (Fetterly et al., 2005).
Ceftobiprole: Methicillin-resistant S. aureus was cross-resistance to all other β-lactam drugs but a number of other new β-lactam agents are undergoing clinical trials and will soon be added to the therapeutic armamentarium for MRSA infections (Fig. 15). This drug looks extremely promising, especially as empiric therapy against both hospital and community acquired pathogens (Bogdanovich et al., 2005). Ceftobiprole is a fourth-generation cephalosporin administered intravenously, with activity against MRSA (Appelbaum, 2006). It is characterized by a strong affinity for penicillin-binding protein PBP2a and PBP2x which are responsible for resistance in Staphylococcus spp. and S. pneumoniae. It is active against gram-negative bacteria, including a high proportion of P. aeruginosa strains and gram-positive bacteria, including MRSA (Zhanel et al., 2008). The terminal elimination half life is 3 h and the predominant mechanism responsible for elimination is glomerular filtration. It does not significantly induce or inhibit relevant cytochrome P450 enzymes and is neither a substrate for nor an inhibitor of P-glycoprotein (Murthy and Schmitt-Hoffmann, 2008).
CONCLUSION
The processes contributing to the reversal of antimicrobial resistance are complex and will differ with antimicrobial consumption patterns, lifestyle, bacterial species and environment. It is clear that antimicrobial resistance is opposite to the good things in life: it is easy to get, but hard to lose. On the positive side, the elimination of a given antimicrobial drug can, when resistance frequencies are very high, relatively quickly reduce the frequency of resistance. Thus, human exposure rates to antimicrobial-resistant bacteria and their transferable genetic elements will be appreciably reduced. In conclusion, numerous questions must ultimately be addressed in order to make significant advances toward reducing disease caused by antibiotic resistant strains of S. aureus. Clinicians should be guided for the optimum incorporation of new antibiotics in place of glycopeptides for the treatment of serious MRSA infections. The search of novel agents with good activity against both susceptible and resistant pathogens is essential.
REFERENCES
- Allen, N.E. and T.I. Nicas, 2003. Mechanism of action of oritavancin and related glycopeptide antibiotics. FEMS Microbiol. Rev., 26: 511-532.
PubMed - Anam, K., A.G. Suganda, E.Y. Sukandar and L.B.S. Kardono, 2010. Antibacterial effect of component of Terminalia muelleri Benth. against Staphylococcus aureus. Int. J. Pharmacol., 6: 407-412.
CrossRef - Archer, G.L., D.M. Niemeyer, J.A. Thanassi and M.J. Pucci, 1994. Dissemination among staphylococci of DNA sequences associated with methicillin resistance. Antimicrob. Agents Chemother., 38: 447-454.
Direct Link - Baba, T., F. Takeuchi, M. Kuroda, H. Yuzawa and K.I. Aoki et al., 2002. Genome and virulence determinants of high virulence community-acquired MRSA. Lancet, 359: 1819-1827.
CrossRefPubMedDirect Link - Barrett, J.F., 2005. Recent developments in glycopeptides antibacterials. Curr. Opin. Investig. Drugs, 6: 781-792.
PubMed - Beibei, L., C. Yun, C. Mengli, B. Nan, Y. Xuhong and W. Rui, 2010. Linezolid versus vancomycin for the treatment of gram-positive bacterial infections: meta-analysis of randomised controlled trials. Int. J. Antimicrob. Agents, 35: 3-12.
PubMed - Blumberg, H.M., D. Rimland, D.J. Carroll, P. Terry and I.K. Wachsmuth, 1991. Rapid development of ciprofloxacin resistance in methicillin-susceptible and-resistant Staphylococcus aureus. J. Infect. Dis., 163: 1279-1285.
CrossRefDirect Link - Bogdanovich, T., L.M. Ednie, S. Shapiro and P.C. Appelbaum, 2005. Antistaphylococcal activity of ceftobiprole, a new broad-spectrum cephalosporin. Antimicrob. Agents Chemother., 49: 4210-4219.
CrossRef - Campbell, S.J., H.S. Deshmukh, C.L. Nelson, I.G. Bae and M.E. Stryjewski et al., 2008. Genotypic characteristics of Staphylococcus aureus isolates from a multinational trial of complicated skin and skin structure infections. J. Clin. Microbiol., 46: 678-684.
CrossRef - CDC., 2002. Staphylococcus aureus resistant to vancomycin-United States, 2002. MMWR Morbidity Mortality Weekly Rep., 51: 565-567.
PubMedDirect Link - Chambers, H.F., 1999. Penicillin-binding protein-mediated resistance in pneumococci and staphylococci. J. Infect. Dis., 179: S353-S359.
PubMed - Champney, W.S. and M. Miller, 2002. Linezolid is a specific inhibitor of 50S ribosomal subunit formation in Staphylococcus aureus cells. Curr. Microbiol., 44: 350-356.
PubMed - Chanda, S., V. BRM, Y. Vaghasiya, H. Patel, 2010. Global Resistance Trends and the Potential Impact of Methicillin Resistant Staphylococcus aureus (MRSA) and its Solutions. In: Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology, Mendez-valas, A. (Ed.). Formatex, Badazoj, Spain, pp: 529-536.
- Collins, J., J. Rudkin, M. Recker, C. Pozzi, J.P. O'Gara and R.C. Massey, 2010. Offsetting virulence and antibiotic resistance costs by MRSA. ISME J., 4: 577-584.
CrossRefDirect Link - Cosgrove, S.E., Y. Qi, K.S. Kaye, S. Harbarth, A.W. Karchmer and Y. Carmeli, 2005. The impact of methicillin resistance in Staphylococcus aureus bacteremia on patient outcomes: Mortality, length of stay and hospital charges. Infect. Control Hosp. Epidemiol., 26: 166-174.
PubMed - Courvalin, P., 2006. Vancomycin resistance in gram-positive cocci. Clin. Infect. Dis., 42: S25-S34.
PubMed - Craven, D.E., C. Reed, N. Kollisch, A. DeMaria, D. Lichtenberg, K. Shen and W.R. McCabe, 1981. A large outbreak of infections caused by a strain of Staphylococcus aureus resistant of oxacillin and aminoglycosides. Am. J. Med., 71: 53-58.
PubMed - Dadgar, T., M. Asmar, A. Saifi, M. Mazandarani and H. Bayat et al., 2006. Antibacterial activity of certain iranian medicinal plants against methicillin-resistant and sensitive Staphylococcus aureus. Asian J. Plant Sci., 5: 861-866.
CrossRefDirect Link - Agostino, C.D., L. Scorzolini, A.P. Massetti, M. Carnevalini and G.D. Ettorre et al., 2010. A seven-year prospective study on spondylodiscitis: epidemiological and microbiological features. Infection, 38: 102-107.
PubMed - Daum, R.S., T. Ito, K. Hiramatsu, F. Hussain, K. Mongkolrattanothai, M. Jamklang and S. Boyle-Vavra, 2002. A novel methicillin-resistance cassette in community-acquired methicillin-resistant Staphylococcus aureus isolates of diverse genetic backgrounds. J. Infect. Dis., 186: 1344-1347.
PubMed - Diekema, D.J., M.A. Pfaller, F.J. Schmitz, J. Smayevsky, J. Bell, R.N. Jones and M. Beach, 2001. Survey of infections due to Staphylococcus species: Frequency of occurrence and antimicrobial susceptibility of isolates collected in the United States, Canada, Latin America, Europe and the Western pacific region for the SENTRY antimicrobial surveillance program, 1997-1999. Clin. Infect. Dis., 32: S114-S132.
CrossRefPubMedDirect Link - Eisenstein, B.I., 2004. Lipopeptides, focusing on daptomycin, for the treatment of Gram-positive infections. Expert Opin. Investig. Drugs, 13: 1159-1169.
PubMed - Elliott, R.A., H.L. Weatherly, N.S. Hawkins, G. Cranny and D. Chambers et al., 2010. An economic model for the prevention of MRSA infections after surgery: non-glycopeptide or glycopeptide antibiotic prophylaxis?. Eur. J. Health Econ., 11: 57-66.
PubMed - Enoch, D.A., J.A. Karas and S.H. Aliyu, 2009. Oral antimicrobial options for the treatment of skin and soft-tissue infections caused by methicillin-resistant Staphylococcus aureus (MRSA) in the UK. Int. J. Antimicrob. Agents, 33: 497-502.
PubMed - Entenza, J.M. and P. Moreillon, 2009. Tigecycline in combination with other antimicrobials: a review of In vitro, animal and case report studies. Int. J. Antimicrob. Agents, 34: 8e1-8e9.
PubMed - Falagas, M.E., I.I. Siempos and K.Z. Vardakas, 2008. Linezolid versus glycopeptide or beta-lactam for treatment of Gram-positive bacterial infections: Meta-analysis of randomised controlled trials. Lancet Infect. Dis., 8: 53-66.
PubMedDirect Link - Farley, J.E., 2008. Epidemiology, clinical manifestations, and treatment options for skin and soft tissue infection caused by community-acquired methicillin-resistant Staphylococcus aureus. J. Am. Acad. Nurse Pract., 20: 85-92.
PubMed - Ferry, T., T. Perpoint, F. Vandenesch and J. Etienne, 2005. Virulence determinants in Staphylococcus aureus and their involvement in clinical syndromes. Curr. Infect. Dis. Rep., 7: 420-428.
PubMed - Fetterly, G.J., C.M. Ong, S.M. Bhavnani, J.S. Loutit and S.B. Porter et al., 2005. Pharmacokinetics of oritavancin in plasma and skin blister fluid following administration of a 200-milligram dose for 3 days or a single 800-milligram dose. Antimicrob. Agents Chemother., 49: 148-152.
PubMed - Foster, T.J. and M. Hook, 1998. Surface protein adhesins of Staphylococcus aureus. Trends Microbiol., 6: 484-488.
PubMed - Fowler, T., E.R. Wann, D. Joh, S. Johansson, T.J. Foster and M. Hook, 2000. Cellular invasion by Staphylococcus aureus involves a fibronectin bridge between the bacterial fibronectin-binding MSCRAMMs and host cell β1 integrins. Eur. J. Cell Biol., 79: 672-679.
PubMed - Fowler, Jr. V.G., H.W. Boucher, G.R. Corey, E. Abrutyn and A.W. Karchmer et al., 2006. Daptomycin versus standard therapy for bacteremia and endocarditis caused by Staphylococcus aureus. Neng. J. Med., 355: 653-665.
Direct Link - French, G.L., 2005. Clinical impact and relevance of antibiotic resistance. Adv. Drug Del. Rev., 57: 1514-1527.
CrossRefDirect Link - Gally, D. and A.R. Archibald, 1993. Cell wall assembly in Staphylococcus aureus: Proposed absence of secondary crosslinking reactions. J. Gen. Microbiol., 139: 1907-1913.
PubMed - Gally, D.L., I.C. Hancock, C.R. Harwood and A.R. Archibald, 1991. Cell wall assembly in Bacillus megaterium: Incorporation of new peptidoglycan by a monomer addition process. J. Bacteriol., 173: 2548-2555.
Direct Link - Gander, S., A. Kinnaird and R. Finch, 2005. Telavancin: in vitro activity against staphylococci in a biofilm model. J. Antimicrob. Chemother., 56: 337-343.
CrossRef - Ghuysen, J.M. and J.L. Strominger, 1963. Structure of the cell wall of Staphylococcus aureus, strain copenhagen II separation and structure of disaccharides. Biochemistry, 2: 1119-1125.
CrossRef - Gonzalez, B.E., G. Martinez-Aguilar, K.G. Hulten, W.A. Hammerman and J. Coss-Bu et al., 2005. Severe Staphylococcal sepsis in adolescents in the era of community-acquired methicillin-resistant Staphylococcus aureus. Pediatrics, 115: 642-648.
PubMed - Greene, C., D. McDevitt, P. Francois, P.E. Vaudaux, D.P. Lew and T.J. Foster, 1995. Adhesion properties of mutants of Staphylococcus aureus defective in fibronectin-binding proteins and studies on the expression of fnb genes. Mol. Microbiol., 17: 1143-1152.
PubMed - Hash, J.H. and M.V. Rothlauf, 1967. The N,O-diacetylmuramidase of Chalaropsis species. I. Purification and crystallization. J. Biol. Chem., 242: 5586-5590.
PubMed - Herold, B.C., L.C. Immergluck, M.C. Maranan, D.S. Lauderdale and R.E. Gaskin et al., 1998. Community-acquired methicillin-resistant Staphylococcus aureus in children with no identified predisposing risk. J. Am. Med. Assco., 279: 593-598.
PubMedDirect Link - Hiramatsu, K., 2001. Vancomycin-resistant Staphylococcus aureus: A new model of antibiotic resistance. Lancet Infect. Dis., 1: 147-155.
CrossRefPubMedDirect Link - Hisata, K., T. Ito, N. Matsunaga, M. Komatsu and J. Jin et al., 2011. Dissemination of multiple MRSA clones among community-associated methicillin-resistant Staphylococcus aureus infections from Japanese children with impetigo. J. Infect. Chemother., 17: 609-621.
PubMed - Hooper, D.C. and J.S. Wolfson, 1991. Fluoroquinolone antimicrobial agents. N. Engl. J. Med., 324: 384-394.
CrossRefPubMedDirect Link - Hutchinson, D.K., 2003. Oxazolidinone antibacterial agents: A critical review. Curr. Top. Med. Chem., 3: 1021-1042.
PubMed - Ito, T., Y. Katayama, K. Asada, N. Mori, K. Tsutsumimoto, C. Tiensasitorn and K. Hiramatsu, 2001. Structural comparison of three types of staphylococcal cassette chromosome mec integrated in the chromosome in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother., 45: 1323-1336.
CrossRefDirect Link - Jasmine, R., P. Daisy and B.N. Selvakumar, 2007. Evaluating the antibacterial activity of Elephantopus scaber extracts on clinical isolates of β-lactamase producing methicillin resistant Staphylococcus aureus from UTI patients. Int. J. Pharmacol., 3: 165-169.
CrossRefDirect Link - Johnson, M.D. and C.F. Decker, 2008. Antimicrobial agents in treatment of MRSA infections. Dis. Mon., 54: 793-800.
PubMed - Jones, R.N., M.G. Stilwell, H.S. Sader, T.R. Fritsche and B.P. Goldstein, 2006. Spectrum and potency of dalbavancin tested against 3322 Gram-positive cocci isolated in the United States Surveillance Program (2004). Diagn. Microbiol. Infect. Dis., 54: 149-153.
Direct Link - Jonsson, K., C. Signas, H.P. Muller and M. Lindberg, 1991. Two different genes encode fibronectin binding proteins in Staphylococcus aureus. The complete nucleotide sequence and characterization of the second gene. Eur. J. Biochem., 202: 1041-1048.
PubMed - Kaka, A.S., A.M. Rueda, S.A. Shelburne, K. Hulten, R.J. Hamill and D.M. Musher, 2006. Bactericidal activity of orally available agents against methicillin-resistant Staphylococcus aureus. J. Antimicrob. Chemother., 58: 680-683.
PubMed - Karas, J.A., D.A. Enoch, H.J. Eagle and M.M. Emery, 2009. Random meticillin-resistant Staphylococcus aureus carrier surveillance at a district hospital and the impact of interventions to reduce endemic carriage. J. Hosp. Infect., 71: 327-333.
PubMed - Kassis, C., R. Hachem, I.I. Raad, C.A. Perego and T. Dvorak et al., 2011. Outbreak of community-acquired methicillin-resistant Staphylococcus aureus skin infections among health care workers in a cancer center. Am. J. Infect. Control, 39: 112-117.
PubMed - Katopodis, G.D., I.N. Grivea, A.J. Tsantsaridou, S. Pournaras, E. Petinaki and G.A. Syrogiannopoulos. 2010. Fusidic acid and clindamycin resistance in community-associated, methicillin-resistant Staphylococcus aureus infections in children of Central Greece. BMC Infect. Dis., Vol. 10.
CrossRef - Khorvash, F., K. Mostafavizadeh, S. Mobasherizadeh, M. Behjati and A.E. Naeini et al., 2008. Antimicrobial susceptibility pattern of microorganisms involved in the pathogenesis of surgical site infection (SSI); A 1 year of surveillance. Pak. J. Biol. Sci., 11: 1940-1944.
CrossRefPubMedDirect Link - Klevens, R.M., M.A. Morrison and J. Nadle, 2007. Invasive methicillin-resistant Staphylococcus aureus infections in the United States. J. Am. Med. Assoc., 298: 1763-1771.
PubMed - Kobayashi, N., S. Urasawa, N. Uehara and N. Watanabe, 1998. Analysis of diversity of mutations in the mecI gene and mecA promoter/operator region of methicillin-resistant Staphylococcus aureus and Staphylococcus epidermidis. Antimicrob. Agents Chemother., 42: 717-720.
PubMed - Kuhl, S.A., P.A. Pattee and J.N. Baldwin, 1978. Chromosomal map location of the methicillin resistance determinant in Staphylococcus aureus. J. Bacteriol., 135: 460-465.
Direct Link - Labandeira-Rey, M., F. Couzon, S. Boisset, E.L. Brown and M. Bes et al., 2007. Staphylococcus aureus Panton-Valentine leukocidin causes necrotizing pneumonia. Science, 315: 1130-1133.
CrossRefDirect Link - Lakshmi, P.T.V., S. Radhika and A. Annamalai, 2011. Molecular docking analysis of phyto-ligands with multi drug resistant β-lactamases of Staphylococcus aureus. Trends Bioinform., 4: 23-34.
CrossRefDirect Link - Le, J. and J.M. Lieberman, 2006. Management of community-associated methicillin-resistant Staphylococcus aureus infections in children. Pharmacotherapy, 26: 1758-1770.
PubMed - Leonard, S.N. and M.J. Rybak, 2008. Telavancin: An antimicrobial with a multifunctional mechanism of action for the treatment of serious gram-positive infections. Pharmacotherapy, 28: 458-468.
PubMed - Levin, B.R., 2001. Minimizing potential resistance: A population dynamics view. Clin. Infect. Dis., 33: S161-S169.
PubMed - Livermore, D.M., 2005. Tigecycline: what is it, and where should it be used?. J. Antimicrob. Chemother., 56: 611-614.
CrossRef - Lodise, T.P., P.S. McKinnon and M. Rybak, 2003. Prediction model to identify patients with Staphylococcus aureus bacteremia at risk for methicillin resistance. Infect. Control Hosp. Epidemiol., 24: 655-661.
PubMed - Lopez, S., C. Hackbarth, G. Romano, J. Trias, D. Jabes and B.P. Goldstein, 2005. In vitro antistaphylococcal activity of dalbavancin, a novel glycopeptide. J. Antimicrob. Chemother., 66: 21-24.
PubMed - Lowy, F.D., 1998. Staphylococcus aureus infections. N. Engl. J. Med., 339: 520-532.
CrossRefPubMedDirect Link - Maree, C.L., R.S. Daum, S. Boyle-Vavra, K. Matayoshi and L.G. Miller, 2007. Community-associated methicillin-resistant Staphylococcus aureus isolates causing healthcare-associated infections. Emerg. Infect. Dis., 13: 236-242.
Direct Link - Martinez-Aguilar, G., W.A. Hammerman, E.O. Mason and S.L. Kaplan, 2003. Clindamycin treatment of invasive infections caused by community-acquired, methicillin-resistant and methicillin-susceptible Staphylococcus aureus in children. Pediatr. Infect. Dis. J., 22: 593-598.
PubMed - Mascio, C.T., J.D. Alder and J.A. Silverman, 2007. Bactericidal action of daptomycin against stationary-phase and nondividing Staphylococcus aureus cells. Antimicrob. Agents Chemother., 51: 4255-4260.
PubMed - Mazmanian, S.K., H. Ton-That and O. Schneewind, 2001. Sortase-catalysed anchoring of surface proteins to the cell wall of Staphylococcus aureus. Mol. Microbiol., 40: 1049-1057.
PubMed - McDougal, L.K. and C. Thornsberry, 1986. The role of β-lactamase in staphylococcal resistance to penicillinase-resistant penicillins and cephalosporins. J. Clin. Microbiol., 23: 832-839.
Direct Link - McGowan, J.E., 1983. Antimicrobial resistance in hospital organisms and its relation to antibiotic use. Rev. Infect. Dis., 5: 1033-1048.
PubMed - Mercier, R.C. and L. Hrebickova, 2005. Oritavancin: A new avenue for resistant Gram-positive bacteria. Expert Rev. Anti Infect. Ther., 3: 325-332.
PubMed - Mertz, D., R. Frei, N. Periat, C. Scheidegger, M. Battegay, W. Seiler and A.F. Widmer, 2010. Eradication of an epidemic Methicillin-Resistant Staphylococcus aureus (MRSA) from a geriatric university hospital: Evidence from a 10-year follow-up. Eur. J. Clin. Microbiol. Infect. Dis., 29: 987-993.
PubMed - Miller, L.G. and S.L. Kaplan, 2009. Staphylococcus aureus: a community pathogen. Infect. Dis. Clin. North Am., 23: 35-52.
PubMed - Murthy, B. and A. Schmitt-Hoffmann, 2008. Pharmacokinetics and pharmacodynamics of ceftobiprole, an anti-MRSA cephalosporin with broad-spectrum activity. Clin. Pharmacokinet., 47: 21-33.
PubMed - Mutnick, A.H., D.J. Biedenbach, J.D. Turnidge and R.N. Jones, 2002. Spectrum and potency evaluation of a new oxazolidinone, linezolid: Report from the SENTRY Antimicrobial Surveillance Program, 1998-2000. Diagn. Microbiol. Infect. Dis., 43: 65-73.
PubMed - Nair, R. and S. Chanda, 2005. Anticandidal activity of Punica granatum exhibited in different solvents. Pharmaceut. Biol., 43: 21-25.
CrossRefDirect Link - Navarre, W.W., H. Ton-That, K.F. Faull and O. Schneewind, 1998. Anchor structure of staphylococcal surface proteins. II. Cooh-terminal structure of muramidase and amidase-solubilized surface protein. J. Biol. Chem., 273: 29135-29142.
PubMed - Ni Eidhin, D., S. Perkins, P. Francois, P. Vaudaux, M. Hook and T.J. Foster, 1998. Clumping factor B (ClfB), a new surface-located fibrinogen-binding adhesin of Staphylococcus aureus. Mol. Microbiol., 30: 245-257.
PubMed - Oliveira, D.C., A. Tomasz and H. de Lencastre, 2002. Secrets of success of a human pathogen: molecular evolution of pandemic clones of meticillin-resistant Staphylococcus aureus. Lancet Infect. Dis., 2: 180-189.
PubMed - Padayachee, T. and K.P. Klugman, 1999. Molecular basis of rifampin resistance in Streptococcus pneumoniae. Antimicrob. Agents Chemother., 43: 2361-2365.
PubMed - Patel, H., A. Shah, M. Mistry and S. Chanda, 2011. In vitro antimicrobial susceptibility study in clinical isolates of streptococci and enterococci. Afr. J. Micro. Res., 5: 1374-1378.
Direct Link - Peeters, M.J. and J.C. Sarria, 2005. Clinical characteristics of linezolid-resistant Staphylococcus aureus infections. Am. J. Med. Sci., 330: 102-104.
PubMed - Perry, A.M., H. Ton-That, S.K. Mazmanian and O. Schneewind, 2002. Anchoring of surface proteins to the cell wall of Staphylococcus aureus. III. Lipid II is an In vivo peptidoglycan substrate for sortase-catalyzed surface protein anchoring. J. Biol. Chem., 277: 16241-26248.
PubMed - Poulakou, G. and H. Giamarellou, 2008. Oritavancin: a new promising agent in the treatment of infections due to Gram-positive pathogens. Expert Opin. Investig. Drugs, 17: 225-243.
PubMed - Projan, S.J. and P.A. Bradford, 2007. Late stage antibacterial drugs in the clinical pipeline. Curr. Opin. Microbiol., 10: 441-446.
PubMed - Raad, I., R. Darouiche, J. Vazquez, A. Lentnek and R. Hachem et al., 2005. Efficacy and safety of weekly dalbavancin therapy for catheter-related bloodstream infection caused by gram-positive pathogens. Clin. Infect. Dis., 40: 374-380.
PubMedDirect Link - Rahbar, M. and M. Hajia, 2007. Inducible clindamycin resistance in Staphylococcus aureus: A cross-sectional report. Pak. J. Biol. Sci., 10: 189-192.
PubMedDirect Link - Rimland, D., 1985. Nosocomial infections with methicillin and tobramycin resistant Staphylococcus aureus implication of physiotherapy in hospital-wide dissemination. Am. J. Med. Sci., 290: 91-97.
PubMed - Rodriguez-Bano, J., A.B. Millan, M.A. Dominguez, C. Borraz and M.P. Gonzalez et al., 2009. Impact of inappropriate empirical therapy for sepsis due to health care-associated methicillin-resistant Staphylococcus aureus. J. Infect., 58: 131-137.
PubMed - Rolston, K.V., H. Nguyen and M. Messer, 1990. In vitro activity of LY264826, a new glycopeptide antibiotic, against gram-positive bacteria isolated from patients with cancer. Antimicrob. Agents Chemother., 34: 2137-2141.
Direct Link - Rubin, R. J., C.A. Harrington, A. Poon, K. Dietrich, J.A. Greene and A. Moiduddin, 1999. The economic impact of Staphylococcus aureus infection in New York City hospitals. Emerging Infect. Dis., 5: 9-17.
PubMedDirect Link - Rubinstein, E., T. Lalani, G.R. Corey, Z.A. Kanafani and E.C. Nannini et al., 2011. Telavancin versus vancomycin for hospital-acquired pneumonia due to gram-positive pathogens. Clin. Infect. Dis., 52: 31-40.
PubMed - Sakoulas, G. and R.C. Moellering, 2008. Increasing antibiotic resistance among methicillin-resistant Staphylococcus aureus strains. Clin. Infect. Dis., 46: S360-S367.
PubMed - Savignon-Marinho, D., G. Huff, B.L. Ferreira, H. Castro and C.R. Rodrigues et al., 2011. The study of vancomycin use and its adverse reactions associated to patients of a brazilian university hospital. BMC Res. Notes, Vol. 4.
CrossRef - Schentag, J.J., J.M. Hyatt, J.R. Carr, J.A. Paladino, M.C. Birmingham, G.S. Zimmer and T.J. Cumbo, 1998. Genesis of Methicillin-Resistant Staphylococcus aureus (MRSA), how treatment of MRSA infections has selected for vancomycin-resistant Enterococcus faecium and the importance of antibiotic management and infection control. Clin. Infect. Dis., 26: 1204-1214.
Direct Link - Schleifer, K.H. and O. Kandler, 1972. Peptidoglycan types of bacterial cell walls and their taxonomic implications. Bacteriol. Rev., 36: 407-477.
Direct Link - Seltzer, E., M.B. Dorr, B.P. Goldstein, M.Perry, J.A. Dowell and T. Henkel, 2003. Once-weekly dalbavancin versus standard-of-care antimicrobial regimens for treatment of skin and soft-tissue infections. Clin. Infect. Dis., 37: 1298-1303.
PubMedDirect Link - Shrivastava, S.M., S. Kumar and M. Chaudhary, 2009. Comparative evaluation of fixed dose combination of ofloxacin and ornidazole against some aerobic bacteria. Trends Med. Res., 4: 30-34.
CrossRefDirect Link - Siddiqi, F., Madahiah-bint-e-Masood, Noor-us-Saba, A. Samad, M. Qayyum and A.A. Qazilbash, 2002. Antibiogram sensitivity pattern of methicillin-resistant Staphylococcus aureus isolates from pus samples. Pak. J. Biol. Sci., 5: 491-493.
CrossRefDirect Link - Simo, R. and G. French, 2006. The use of prophylactic antibiotics in head and neck oncological surgery. Curr. Opin. Otolaryngol. Head Neck Surg., 14: 55-61.
PubMed - Sjoquist, J., J. Movitz, I.B. Johansson and H. Hjelm, 1972. Localization of protein A in the bacteria. Eur. J. Biochem., 30: 190-194.
PubMed - Skiest, D.J., 2006. Treatment failure resulting from resistance of Staphylococcus aureus to daptomycin. J. Clin. Microbiol., 44: 655-656.
CrossRef - Stryjewski, M.E. and H.F. Chambers, 2008. Skin and soft-tissue infections caused by community-acquired methicillin-resistant Staphylococcus aureus. Clin. Infect. Dis., 46: S368-3S77.
CrossRef - Thurnheer, R., I. Laube and R. Speich, 1999. Possible interaction between clindamycin and cyclosporin. Br. Med. J., Vol. 319.
CrossRef - Tipper, D.J. and J.L. Strominger, 1965. Mechanism of action of penicillins: a proposal based on their structural similarity to acyl-D-alanyl-D-alanine. Proc. Natl. Acad. Sci., 54: 1133-1141.
Direct Link - Truong, Q.C., J.C. Nguyen Van, D. Shlaes, L. Gutmann and N.J. Moreau, 1997. A novel, double mutation in DNA gyrase A of Escherichia coli conferring resistance to quinolone antibiotics. Antimicrob. Agents Chemother., 41: 85-90.
Direct Link - Tsuji, B.T., M.J. Rybak, K.L. Lau and G. Sakoulas. 2007. Evaluation of Accessory Gene Regulator (agr) Group and Function in the Proclivity towards Vancomycin Intermediate Resistance in Staphylococcus aureus. Antimicrob. Agents Chemother., 51: 1089-1091.
CrossRefDirect Link - Tuzuner-Oncul, A.M., C. Ungor, U. Dede and R.S. Kisnisci, 2009. Methicillin-resistant Staphylococcus aureus (MRSA) osteomyelitis of the mandible. Oral Surg. Oral Med. Oral Pathol.Oral Radiol. Endod., 107: e1-e4.
PubMed - Vaghasiya, Y. and S.V. Chanda, 2007. Screening of methanol and acetone extracts of fourteen Indian medicinal plants for antimicrobial activity. Turk. J. Biol., 31: 243-248.
Direct Link - Van Bambeke, F., Y. Van Laethem, P. Courvalin and P.M. Tulkens, 2004. Glycopeptide antibiotics: From conventional molecules to new derivatives. Drugs, 64: 913-936.
PubMed - Van Heijenoort, J. and L. Gutmann, 2000. Correlation between the structure of the bacterial peptidoglycan monomer unit, the specificity of transpeptidation, and susceptibility to beta-lactams. Proc. Natl. Acad. Sci., 97: 5028-5030.
Direct Link - Ward, P.B., P.D. Johnson, E.A. Grabsch, B.C. Mayall and M.L. Grayson, 2001. Treatment failure due to Methicillin-Resistant Staphylococcus aureus (MRSA) with reduced susceptibility to vancomycin. Med. J. Aust., 175: 480-483.
PubMed - Wehrli, W., 1983. Rifampin: Mechanisms of action and resistance. Rev. Infect. Dis., 5: S407-S411.
CrossRefPubMedDirect Link - Weller, T.M., 1999. The distribution of mecA, mecR1 and mecI and sequence analysis of mecI and the mec promoter region in staphylococci expressing resistance to methicillin. J. Antimicrob. Chemother., 43: 15-22.
PubMed - Wichelhaus, T.A., V. Schafer, V. Brade and B. Boddinghaus, 1999. Molecular characterization of rpoB mutations conferring cross-resistance to rifamycins on methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother., 43: 2813-2816.
Direct Link - Widmer, A.F., R. Frei, Z. Rajacic and W. Zimmerli, 1990. Correlation between In vivo and In vitro efficacy of antimicrobial agents against foreign body infections. J. Infect. Dis., 162: 96-102.
PubMed - Yoke-Kqueen, C., J. Laurence and S. Radu, 2006. Characterization of Staphylococcus aureus isolated from the skin surface of athletes and training environment by random amplified polymorphic DNA and antibiotic resistance profiling. Biotechnology, 5: 489-494.
CrossRefDirect Link - Zhanel, G.G., A. Lam, F. Schweizer, K. Thomson and A. Walkty et al., 2008. Ceftobiprole: A review of a broad-spectrum and anti-MRSA cephalosporin. Am. J. Clin. Dermatol., 9: 245-254.
PubMed - Zuckerman, J.M., 2004. Macrolides and ketolides: Azithromycin, clarithromycin, telithromycin. Infect. Dis. Clin. North Am., 18: 621-649.
PubMed