
Martha Purnami Wulanjati
Department of Pharmacy, Faculty of Medicine and Health Sciences, Atma Jaya Catholic University of Indonesia, Jl. Pluit Raya 2, 14440 Jakarta, Indonesia
LiveDNA: 62.23153
Nastiti Wijayanti
Department of Animal Physiology, Faculty of Biology, Universitas Gadjah Mada, Jl. Teknika Selatan, Sekip Utara, Bulaksumur, 55281 Yogyakarta, Indonesia
Aris Haryanto
Department of Biochemistry and Molecular Biology, Faculty of Veterinary Medicine, Universitas Gadjah Mada, Jl. Fauna 2, Karangmalang, 55281 Yogyakarta, Indonesia
LiveDNA: 62.12752







ABSTRACT
Background and Objective: Newcastle disease (ND) is an infectious disease caused by ND virus (NDV), which is the major problem in poultry industries. Although vaccination program has been executed in Indonesia, Newcastle disease is still infecting chicken. The lack of vaccine protection against the disease, presumably due to genetic differences between vaccine strain and virus strain in the field. This study was conducted to analyze phylogenetic of Indonesian NDV isolates based on fusion (F) protein-encoding gene, with aim to determine which genotype group of Indonesian NDV isolates, compared to vaccine strain that circulating in Indonesia. Materials and Methods: Samples were RNA genome of NDV, which were isolated from chicken in Kartosuro, Karanganyar; Galur, Kulon Progo and Sukomoro, Magetan areas. The F gene was amplified by one step RT-PCR method and then was sequenced. The genetic variation among samples, other Indonesian isolates, LaSota and B1 vaccine strain were analyzed by pairwise distance method. Phylogenetic tree based on F gene sequence was generated by Neighbor-Joining method (1000 bootstrap repetition) and Kimura-2 parameter model. Result: The genetic variation of F gene nucleotide and F protein amino acid between samples and vaccine strains were higher compared to the variation between samples and other Indonesian isolates. Phylogenetic analysis showed that the samples were included in genotype VII class II, while LaSota and B1 vaccine strain included in genotype II class II. Conclusion: There were differences of fusion protein antigen and genotype group between Indonesian strains and vaccine strains. This finding encouraged to develop ND vaccine based on Indonesian isolates.
PDF Abstract XML References Citation
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Martha Purnami Wulanjati, Nastiti Wijayanti and Aris Haryanto, 2018. Phylogenetic Analysis of Newcastle Disease Virus from Indonesian Isolates Based on DNA-Sequence of Fusion Protein-Encoding Gene. Biotechnology, 17: 69-74.
DOI: 10.3923/biotech.2018.69.74
URL: https://scialert.net/abstract/?doi=biotech.2018.69.74
DOI: 10.3923/biotech.2018.69.74
URL: https://scialert.net/abstract/?doi=biotech.2018.69.74
INTRODUCTION
Newcastle disease (ND) is the main problem in poultry industry in many countries. In developing countries, ND is an endemic disease that inhibit the progress of poultry industry1. Office International des Epizooties (OIE) categorizes ND in the list A of animal disease, a group of infectious diseases that spread fast and seriously, give impact in public health and affect socio-economic communities2. The outbreak of ND can have a devastating effect as it caused almost 100% mortality in susceptible chicken3. It is also reported that ND causes significant economic losses in many countries, including in Indonesia4.
The etiological agent of ND is Avian Paramyxovirus Serotype-1 (APMV-1) or known as Newcastle disease virus (NDV)5. The NDV is included in the genus Avulavirus, subfamily Paramyxovirinae, family Paramyxoviridae, order Mononegavirales6. The NDV genome is single stranded RNA (ssRNA) negative sense which encodes six proteins, namely nucleocapsid (N), phosphoprotein (P), matrix (M), fusion protein (F), hemagglutinin-neuraminidase (HN) and RNA polymerase (L)7. Infection of the NDV in the host cell is mediated by two viral surface glycoproteins, HN and F-protein8.
The cleavage site of Fo protein is one of the factors determining the systemic spread and virulence of NDV9,10. The cleavage specificity is determined by the amino acid sequence on the cleavage site of F protein and varies in strain types. Low virulent NDV strains contain monobasic or dibasic amino acids on the cleavage site of F protein, which is not sensitive to intracellular proteases, thus depending on the extracellular proteases in the host respiratory and enteric tract. High virulent NDV strains have multiple basic amino acids on the cleavage site that it can be recognized by furin (intracellular protease)10. High virulent NDV strains contain lysine (K) and arginine (R) with motif 112R-R-Q-R/K-R116 on C terminus F2 and phenylalanine (F) at position 117 N terminus F19.
The clinical symptom of ND is difficult to distinguish with other viral diseases in poultry such as avian influenza which caused by Avian Influenza Virus (AIV) and avian encephalomyelitis caused by Avian Encephalomyelitis Virus (AEV), so that technique for differential diagnosis of ND is essential to detect mixed viral infection11,12. To prevent of ND infection, a vaccination program has been conducted13. However, the outbreak of ND still occurs despite of intensive vaccination4. In 2009 and 2010, the outbreak of ND occurred in poultry farms in Indonesia causing 70-80% mortality from total chickens14. The NDV vaccine designed based on phylogenetic proximity to the NDV that occurs during outbreaks can provide better control of ND15. The objective of this study was to determine genotyping of Indonesian NDV isolates based on F gene sequence. Protein F plays an important role in determining the virulence of NDV. It can induce host-protective immunity against NDV infection. Therefore, a study to determine the genotype of NDV based on the sequence of F protein encoding gene is important meaning to determine the antigenic and genotypic differences between Indonesian NDV isolates with vaccine strains (LaSota and B1) circulating in Indonesia to develop strategy for ND prevention in chicken.
MATERIALS AND METHODS
The study was carried out in October-December, 2016. The materials and methods used in the study is as follows:
Samples: Samples were RNA genome of NDV, which were isolated from broiler chicken in Kartosuro, Karanganyar (0627/04/2013), native chicken from Galur, Kulon Progo (0663/04/2013) and broiler chicken from Sukomoro, Magetan (0819/05/2013)16.
Amplification of full-length F gene: The full-length F gene was amplified using one step RT-PCR method (Transcriptor, Roche) with F gene specific primer, as follows:
| • | Forward primer (28 bp): 5’GTCAGATCTTGATGGGCTCCAA ACCTTC 3’ | |
| • | Reverse primer (28 bp): 5’ATGAATTCTCACGCTCTTGTGGT GGCTC 3’ | |
RT PCR cycle program was reverse transcription at 50oC for 30 min, initial denaturation at 94°C for 7 min, 40 cycles that consisted of denaturation at 94°C for 20 sec, annealing at 57°C for 30 sec and elongation at 68°C for 1 min 45 sec, final elongation at 68°C for 7 min. The RT-PCR product was analyzed by 1% agarose gel electrophoresis then sequenced with F gene specific primer.
Pairwise distance matrix and phylogenetic tree: The F gene sequences were aligned using CLUSTALW in MEGA 6.0 software and analyzed using BLAST. The genetic variation of F gene and F protein of samples against other 7 Indonesian isolates, B1 and LaSota vaccine strain was analyzed using pairwise distance method17.
Phylogenetic tree based on F gene sequence was constructed using Neighbor-Joining method with 1000 bootstrap repetition and using Kimura-2 model4,18. To build phylogenetic tree, F gene sequences from 18 NDV strains with various genotypes that were taken from GenBank used as follows: chicken/Banjarmasin/010/10 (HQ697254), chicken/Gianyar/013/10 (HQ697257), chicken/Kudus/018/10 (HQ697260), chicken/Kudus/017/10 (HQ697259), chicken/ Sragen/014/10 (HQ697258), chicken/Makassar/003/09 (HQ697256), chicken/Sukorejo/019/10 (HQ697255), chicken/Bali/020/10 (HQ697261), chicken/USA(CA)/ 1083(Fontana)/72 (AY288992), strain Italien (EU293914), strain Miyadera (M18456), chicken/Northern.Ireland/Ulster/67 (AY562991), Australia/v4 (JX524203), LaSota (DQ195265), B1 (AF309418), VG/GA (EU289028), Gamefowl/US(CA)212676/ 2002 (EF520718) and Northern Pintail/US(AK)/196/1998 (EF612277).
RESULTS
F gene amplification and sequencing: The amplification with F gene specific primers in the three NDV RNA genomes showed a single band with size of 1681 bp (Fig. 1). The sequencing product of F gene from Karanganyar and Magetan were 1613 bp, while F gene from Kulon Progo was 1607 bp.
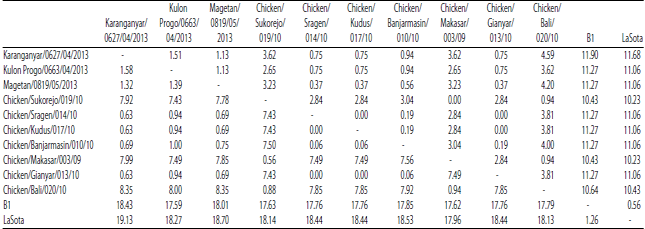
Pairwise distance analysis of F gene nucleotide sequence and F protein amino acid sequence: The result of pairwise distance analysis of F gene and F protein amino acid sequence among 7 Indonesian NDV isolates, LaSota and B1 vaccine strain was presented in Table 1.
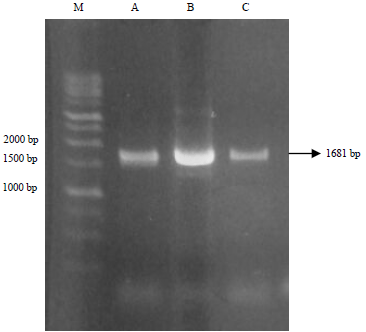 | |
| Fig. 1: | Visualization of 1.5% agarose gel electrophoresis of F gene amplification product |
| M: DNA ladder 1 kb (Geneaid). A: Amplicon of Karanganyar (0627/04/2013), B: Amplicon of Kulon Progo (0663/04/2013) and C: Amplicon of Magetan (0819/05/2013) | |
| Table 1: | Matrix of pairwise distance values (%) of F gene nucleotide (below diagonal matrix) and F protein amino acid |
 | |
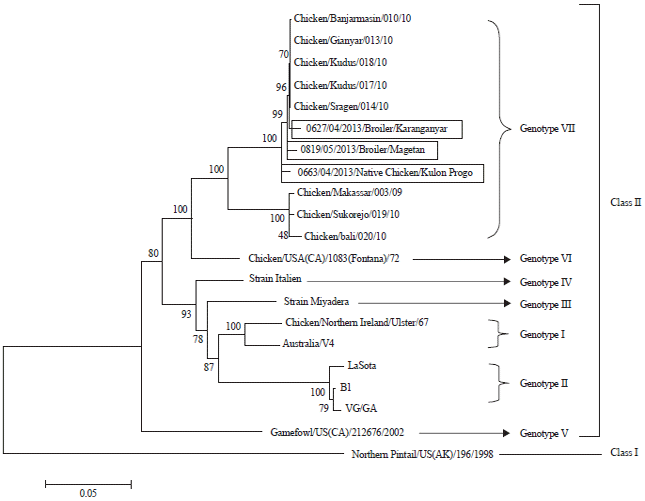 | |
| Fig. 2: | Phylogenetic tree based on F gene sequence of NDV isolates. Phylogenetic tree was generated by Neighbor-Joining method (1000 bootstrap repetition) and Kimura-2 parameter model |
The distance values of F gene nucleotide were below the diagonal matrix while the distance values of F protein amino acid were above the diagonal matrix. Based on pairwise distance matrix data, the three samples showed a higher variation of F gene nucleotide to LaSota and B1 vaccine strains compared with Indonesian isolates. The F gene nucleotide variation of the three samples against the vaccine strains B1 and LaSota were 17.59-18.43 and 18.27-19.13%, respectively, while the F gene nucleotide variation of the three samples against 7 Indonesian isolates ranged from 0.63-8.35%. This pattern was the same as the variation of F protein amino acid. Distance value of F protein amino acid sequence of the three samples against 7 Indonesian isolates ranged from 0.37-4.59%. These values were lower than the distance values of F protein amino acid of the samples to LaSota and B1 vaccine strain which was ranged between 11.27-11.9 and 11.06-11.68%.
Phylogenetic tree: Phylogenetic tree based on F gene nucleotide sequence could be depicted in Fig. 2. Phylogenetic analysis showed that isolates from Karanganyar, Kulon Progo and Magetan belonged to genotype VII class II, which also consisted of other Indonesian NDV isolates. This was dissimilar to the LaSota and B1 vaccine strain that belonged to genotype II class II.
DISCUSSION
Newcastle disease (ND) is still infecting poultry in Indonesia, although ND vaccination has been implemented. In Indonesia, LaSota and B1 vaccine are choice vaccines that commonly used for prevention of ND in chickens. LaSota and B1 vaccine agents were included in the low virulent viruses1. The three samples in this study, namely Karanganyar (0627/04/2013), Kulon Progo (0663/04/2013) and Magetan (0819/05/2013) were high virulent viruses16. The samples were characterized by determination of amino acid sequence on the cleavage site of Fo protein on amino acid No. 112-117. The virulence of NDV can be determined not only by the presence of multiple basic amino acids in cleavage region but also by molecular pathotyping using RT-PCR and restriction fragment length polymorphism (RFLP) methods using HinfI restriction enzyme19.
In this study, the pairwise distance and phylogenetic were analyzed based on fusion (F) protein sequence of NDV. Fusion protein has an important role in virulence of the virus20. Fusion protein could induce protective immunity in chicken against NDV21,22. The study of Kim et al.23 showed that F protein from NDV is a major part that provided protective immunity in development of genotype-matched vaccine.
The result of pairwise distance analysis both on nucleotide sequence of F gene and amino acid sequence of F protein of samples against LaSota and B1 vaccine strains showed higher values compared to the values between samples and other Indonesian isolates (Table 1). The results of this genetic variation analysis are in accordance with the previous study of Xiao et al.14. They conducted a homologous analysis of 8 high virulent ND viruses isolated during outbreak in Indonesia in 2009 and 2010. The homologous analysis showed that amino acid sequence identity of F protein among the 8 NDV was higher than the sequence identity value of 8 NDV against B1 and LaSota vaccine strains.
Two different systems were used to classify ND viruses. The first system, developed by Aldous et al.24, grouped the NDV into 6 lineages and 13 sublineages based on phylogenetic analysis of the F gene cleavage activation region and peptide signals. The second system, based on the genome length and nucleotide sequences, classified the NDV into two major groups namely class I and class II. Class I was a group of avirulent viruses that have a genome length of 15198 nucleotides. Class I consisted of 9 genotypes25,26. Class II consisted of 10 genotypes. The initial genotype groups were I, II, III, IV and IX that containing 15186 nucleotides in genome, while the final genotype groups were V, VI, VII, VIII and X containing 15192 nucleotides in genome25. The phylogenetic analysis of this study showed that sample isolates and the other Indonesian isolates belonged to genotype VII class II, while LaSota and B1 vaccine were belonged to genotype II class II (Fig. 2). The result of the phylogenetic analysis was in line with the study of Dharmayanti et al.4 and Xiao et al.14. Their studies showed that Indonesian NDV isolates were included in genotype VII class II. It convinced that majority of NDV circulating in Indonesia belonged in genotype group VII class II.
High genetic variation and different genotype groups based on nucleotide sequences of F gene indicated that there were antigenic differences between NDV isolates from Karanganyar, Kulon Progo and Magetan with the strains of B1 and LaSota commercial vaccines. This antigenic difference may affect the protection of the vaccine against the virus14,15,17. The antigenic differences between ND vaccine strain and NDV strain in the field generated poor and inadequate post-vaccination protection4,14. The previous study of Miller et al.15 stated that vaccines with a high homology to challenge NDV reduced viral shedding significantly compared to vaccines with a low homology to the challenge NDV. Roohani et al.17 showed that in the vaccine efficacy test, the genotype VII NDV recombinant vaccine yielded better protection against the morbidity and mortality of genotype VII NDV isolate compared to LaSota and B1 vaccines. The genotype VII NDV recombinant vaccine also significantly decreased viral shedding compared to LaSota vaccine.
The result of this study that showed antigenic and genotypic distinction between NDV sample and commercial vaccine (LaSota and B1) encouraged to develop ND vaccine based on local NDV isolate. The F proteins from NDV samples that have high homology with F protein from other Indonesian isolate are potential to be vaccine agents in the production of ND recombinant vaccines in Indonesia. For further study, it is suggested to conduct F protein expression and then immunization and challenge test in chicken to investigate the potency of F protein as a vaccine candidate.
CONCLUSION
It is concluded that variation of F gene nucleotide and F protein amino acid sequence between Indonesian NDV isolates (Karanganyar (0627/04/2013), Kulon Progo (0663/04/2013) and Magetan (0819/05/2013) and vaccine strains (LaSota and B1) were higher compared to the variation against other Indonesian isolates. Phylogenetic analysis classified Indonesian isolates into genotype group VII class II, which is different with LaSota and B1 vaccine strain that belonged to genotype group II class II.
SIGNIFICANCE STATEMENT
The current study discovers antigenic and genotypic differences between Indonesian NDV isolates and vaccine strain (LaSota and B1) circulating in Indonesia. This study suggested researchers to formulate ND vaccine based on local virus isolates. The study may be helpful to develop strategy for ND prevention in chicken.
ACKNOWLEDGMENT
This study was funded by research grant "Hibah Berbasis Kompetensi" from Ministry of Research, Technology and Higher Education of the Republic of Indonesia, fiscal year 2015-2017, with grant number 198/LPPM/2015.
REFERENCES
- Alexander, D.J., E.W. Aldous and C.M. Fuller, 2012. The long view: A selective review of 40 years of Newcastle disease research. Avian Pathol., 41: 329-335.
CrossRefPubMedDirect Link - Dharmayanti, N.I., R. Hartawan, D.A. Hewajuli and R. Indriani, 2014. Phylogenetic analysis of genotype VII of Newcastle disease virus in Indonesia. Afr. J. Microbiol. Res., 8: 1368-1374.
Direct Link - Alexander, D.J., 2000. Newcastle disease and other avian paramyxoviruses. Rev. Sci. Tech., 19: 443-462.
CrossRefPubMedDirect Link - De Leeuw, O. and B. Peeters, 1999. Complete nucleotide sequence of Newcastle disease virus: Evidence for the existence of a new genus within the subfamily Paramyxovirinae. J. Gen. Virol., 80: 131-136.
CrossRefDirect Link - Chang, A. and R.E. Dutch, 2012. Paramyxovirus fusion and entry: Multiple paths to a common end. Viruses, 4: 613-636.
CrossRefDirect Link - Ganar, K., M. Das, S. Sinha and S. Kumar, 2014. Newcastle disease virus: Current status and our understanding. Virus Res., 184: 71-81.
CrossRefDirect Link - Miller, P.J. and M.K. Torchetti, 2014. Newcastle disease virus detection and differentiation from avian influenza. Methods Mol. Biol., 1161: 235-239.
CrossRefDirect Link - Haryanto, A., R. Ermawati, V. Wati, S.H. Irianingsih and N. Wijayanti, 2016. Analysis of viral protein-2 encoding gene of avian encephalomyelitis virus from field specimens in Central Java region, Indonesia. Vet. World, 9: 25-31.
CrossRefDirect Link - Xiao, S., A. Paldurai, B. Nayak, A. Samuel and E.E. Bharoto et al., 2012. Complete genome sequences of Newcastle disease virus strains circulating in chicken populations of Indonesia. J. Virol., 86: 5969-5970.
CrossRefDirect Link - Miller, P.J., D.J. King, C.L. Afonso and D.L. Suarez, 2007. Antigenic differences among Newcastle disease virus strains of different genotypes used in vaccine formulation affect viral shedding after a virulent challenge. Vaccine, 25: 7238-7246.
CrossRefDirect Link - Haryanto, A., M. Purwaningrum, S, Verawati, S.H. Irianingsih and N. Wijayanti, 2015. Pathotyping of local isolates Newcastle disease virus from field specimens by RT-PCR and restriction endonuclease analysis. Procedia Chem., 14: 85-90.
CrossRefDirect Link - Roohani, K., S.W. Tan, S.K. Yeap, A. Ideris, M.H. Bejo and A.R. Omar, 2015. Characterisation of genotype VII Newcastle disease virus (NDV) isolated from NDV vaccinated chickens and the efficacy of LaSota and recombinant genotype VII vaccines against challenge with velogenic NDV. J. Vet. Sci., 16: 447-457.
CrossRefDirect Link - Kang, Y., Y. Li, R. Yuan, X. Li and M. Sun et al., 2014. Phylogenetic relationships and pathogenicity variation of two Newcastle disease viruses isolated from domestic ducks in Southern China. Virol. J., Vol. 11, No. 1.
CrossRefDirect Link - Haryanto, A., V. Wati, S. Pakpahan and N. Wijayanti, 2016. Molecular pathotyping of Newcastle disease virus from naturally infected chickens by RT-PCR and RFLP methods. Asian J. Anim. Sci., 10: 39-48.
CrossRefDirect Link - Dortmans, J.C., F.M. Koch, P.J. Rottier and B.P. Peeters, 2011. Virulence of Newcastle disease virus: What is known so far? Vet. Res., Vol. 42.
CrossRef - Arora, P., B.D. Lakhchaura and S.K. Garg, 2010. Evaluation of immunogenic potential of 75kDa and 56kDa proteins of Newcastle Disease Virus (NDV). Indian J. Exp. Biol., 48: 889-895.
Direct Link - Kumar, S., B. Nayak, P.L. Collins and S.K. Samal, 2011. Evaluation of the Newcastle disease virus F and HN proteins in protective immunity by using a recombinant avian paramyxovirus type 3 vector in chickens. J. Virol., 85: 6521-6534.
CrossRefDirect Link - Kim, S.H., N. Wanasen, A. Paldurai, S. Xiao, P.L. Collins and S.K. Samal, 2013. Newcastle disease virus fusion protein is the major contributor to protective immunity of genotype-matched vaccine. PLoS One, Vol. 8, No. 8.
CrossRefDirect Link - Aldous, E.W., J.K. Mynn, J. Banks and D.J. Alexander, 2003. A molecular epidemiological study of avian paramyxovirus type 1 (Newcastle disease virus) isolates by phylogenetic analysis of a partial nucleotide sequence of the fusion protein gene. Avian Pathol., 32: 239-257.
CrossRefDirect Link - Czegledi, A., D. Ujvari, E. Somogyi, E. Wehmann, O. Werner and B. Lomniczi, 2006. Third genome size category of avian paramyxovirus serotype 1 (Newcastle disease virus) and evolutionary implications. Virus Res., 120: 36-48.
CrossRefDirect Link - Kim, L.M., D.J. King, P.E. Curry, D.L. Suarez and D.E. Swayne et al., 2007. Phylogenetic diversity among low-virulence Newcastle disease viruses from waterfowl and shorebirds and comparison of genotype distributions to those of poultry-origin isolates. J. Virol., 81: 12641-12653.
CrossRefPubMedDirect Link