
Vadim Lebedev
Group of Forest Biotechnology, Branch of Shemyakin and Ovchinnikov Institute of Bioorganic Chemistry Russian Academy of Sciences,
Pushchino, Moscow Region, Russia
Konstantin Shestibratov
Group of Forest Biotechnology, Branch of Shemyakin and Ovchinnikov Institute of Bioorganic Chemistry Russian Academy of Sciences,
Pushchino, Moscow Region, Russia







ABSTRACT
An efficient protocol was developed for Micropropagation of common ash (Fraxinus excelsior L.). The in vitro culture was established from shoots of 60 years old plus trees. Single, two and three node segments of decapitated shoots were cultured on MS medium supplemented with 1, 2, or 4 mg L–1 BAP and 0.1 mg L–1 IAA for induction of axillary shoots. The best multiplication rate (7.6) was achieved using single node explants on medium with 1 and 2 mg L–1 BAP. Shoots on all the explant types were significantly taller on the medium with 2 mg L–1 BAP. Efficiency of axillary shoot formation was the highest in distal nodes and reduced toward proximal nodes of explants. Effects of type and concentration of sugar, light intensity and dark treatment on rooting and subsequent acclimatization of microshoots were studied. The significantly greater percentage (72-93%) of shoots formed roots on 0.5 mg L–1 NAA, as compared to 55-73% on 0.5 mg L–1 IBA. The maximum rooting (100%) and highest number of roots per shoot (4.7) were obtained at a Photon Flux Density (PFD) of 25 μmol m–2 sec–1 on the medium with 30 g L–1 sucrose. Dark treatment during 7 days had significant positive effect on the root induction. For acclimatization in the greenhouse, rooted shoots were transplanted into different substrates containing peat, perlite and vermiculite at different ratios. Survival rate in 3 months was 96-100% and all the plants exhibited a normal phenotype. The developed technology may be used for mass propagation of common ash superior trees.
PDF Abstract XML References Citation
Received: October 14, 2015;
Accepted: December 15, 2015;
Published: February 15, 2016
How to cite this article
Vadim Lebedev and Konstantin Shestibratov, 2016. Large-Scale Micropropagation of Common Ash. Biotechnology, 15: 1-9.
DOI: 10.3923/biotech.2016.1.9
URL: https://scialert.net/abstract/?doi=biotech.2016.1.9
DOI: 10.3923/biotech.2016.1.9
URL: https://scialert.net/abstract/?doi=biotech.2016.1.9
INTRODUCTION
The genus Fraxinus comprises about 70 different species of ash trees native to Europe, Asia and North America. Fraxinus excelsior L. (common or European ash) is the most common species in European countries where it is a very important hardwood tree. Because of its strength, hardness, straight grain and elasticity, ash timber is used for the production of tool handles, sports equipment, furniture and flooring. Due to its ornamental qualities, common ash is also widely planted for landscaping. For the large-scale production, F. excelsior is propagated traditionally by seeds that causes considerable phenotypic variation and is not suitable for propagation of superior genotypes. Vegetative propagation of selected clones is very important for the plant improvement, but reliable methods for rooting common ash cuttings are missing1. Ornamental cultivars of ash have been propagated by grafting onto seedling rootstocks. However, this method is labor-intensive and limited by season and therefore it is not suitable for mass production. In vitro culture is an attractive alternative approach for accelerated propagation of selected genotypes of this species. Shoot organogenesis in common ash was mostly achieved by using explants from juvenile plants-seedlings grown in the greenhouse2 or in vitro3,4. The in vitro cultures of F. excelsior were also established from mature trees7,8. This species can also be micropropagated via somatic embryogenesis from zygotic embryos5,6 or adventitious shoot regeneration from epicotyls9. Tissue culture can be used not only for rapid propagation of superior trees, but also for use in genetic modification technology. To our knowledge, there are no reports on common ash genetic transformation, but transgenic plants of F. pennsylvanica10, F. profunda11 and F. americana12 were obtained using hypocotyls as initial explants. However, the best objects for both clonal micropropagation and genetic engineering are plus trees that can only be selected by phenotype at mature age. Therefore, the aim of this research was to develop a protocol for highly efficient large-scale production of F. excelsior plants from vegetative tissues of mature trees. In previous studies, we have optimized the mineral and hormonal composition of the media for multiplication and rooting13 and evaluated effects of natural and synthetic growth stimulators on rooting and acclimatization of common ash microplants14. Here, we evaluated the effect of explant size and BA concentration on the proliferation. We also determined suitable light intensity, carbohydrate type and concentration on rooting and subsequent acclimatization under non-sterile conditions.
MATERIALS AND METHODS
In vitro culture of common ash (F. excelsior L.) was established from shoots of 60 years old plus trees. Apical and axillary buds were washed in running tap water for 30 min, surface sterilized with 0.1% ethylmercury chloride for 6 min and rinsed 3 times with sterile distilled water15. Shoots were subcultured every four weeks in 330 mL glass jars containing 50 mL of MS medium16 supplemented with 30 g L–1 sucrose, 7 g L–1 agar (american type QP, Panreac), 2 mg L–1 BAP and 0.1 mg L–1 IAA. To investigate the effect of explant size, single, two and three node decapitated horizontally oriented shoots were used. These explants were cultured on proliferation medium at a density of 18, 9, or 6 explants per jar, respectively (18 nodes per jar in total). The BAP was added to the medium contained 0.1 mg L–1 IAA at 1, 2 and 4 mg L–1 concentration. The number of shoots, their height and number of nodes per shoot was estimated in four weeks of culture. On the basis of these data, the percentage of responding axillary buds, internode lengths and multiplication rates (ratio of the number of initial nodes to the total number of nodes on produced shoots) were calculated.
For root induction, shoot tips (15-25 mm in length) with 2-3 pairs of leaves were excised from proliferating culture and transferred into 250 mL plastic containers containing 50 mL of WPM medium17 with half strength of macrosalts supplemented with 7 g L–1 agar and 0.5 mg L–1 NAA. The following experiments were carried out to determine the effects of carbohydrates, light intensity and dark treatment on rooting: 1) sucrose (5, 10, or 30 g L–1), glucose (10 g L–1) and maltose (10 g L–1) combined with 0.5 mg L–1 of either NAA or IBA; 2) PFD at 25, 50, or 100 μmol m–2 sec–1 combined with 5, 10, or 30 g L–1 sucrose; 3) 7 days in the darkness followed by 16 h photoperiod, or without dark treatment. The rooting percentage and the number of roots (of more than 2 mm in length) per shoot were determined in 2, 3 and 4 weeks of transfer onto the rooting medium.
The culture media were adjusted with 1 N KOH to pH = 5.6-5.8 and autoclaved at 121°C for 20 min. Growth regulators and vitamins were filter-sterilized (Millipore, 0.22 μm) and added to the media after autoclaving. All the cultures were grown in a 16 h photoperiod provided by cool-white fluorescent lamps giving PFD of 50 μmol m–2 sec–1 (proliferation) or 25 μmol m–2 sec–1 (rooting) and a constant temperature of 23±1°C.
The rooted plants were acclimatized in the greenhouse. Plantlets were washed in running tap water to remove traces of medium, transferred to a substrate containing peat and perlite (3:1) and kept for 3-4 weeks at 22-25°C under relative humidity of 85-90%. The plants after dark treatment experiment were transferred into five different substrates: peat: perlite (2:1, 3:1, or 4:1), peat:vermiculite (3:1) or peat: perlite: vermiculite (1:1:1). After 1, 2 and 3 months, the number of surviving plantlets and the mean shoot height (only after 2 months) were recorded.
Experiments were repeated twice with four replications for each treatment using completely randomized design. Each replication consisted of 6-18 (proliferation), 12-24 (rooting), or 12-16 (acclimatization) microplants. Data were analyzed by using analysis of variance (ANOVA) technique. Percentage data were transformed to arcsine values prior to analysis. Means were evaluated according to Duncan's multiple range test at p = 0.05.
RESULTS
Response to the shoot formation varied with the type of explant (single, two, or three node segments) and also the concentration of BAP in the medium (Table 1). Increasing levels of BAP stimulated growth of axillary buds, but response of larger explants was decreased. The maximal frequency of responding buds (101.9%) was observed at 4 mg L–1 BAP with single-node explants. Thus, high concentration of BAP induced growth of not only the axillary buds of explants (maximal frequency is 100%) but also the adventitious ones. The shoots on explant of all types were significantly taller on the medium with 2 mg L–1 BAP (45.9-57.0 mm) than on medium with 1 and 4 mg L–1 BAP (33.9-40.3 and 31.7-36.7 mm, respectively). Number of nodes on induced shoots slightly varied from 3.6-4.3 nodes per shoot. Internode length positively correlated with total height of shoots. Thus, combination of 2 mg L–1 BAP and 0.1 mg L–1 IAA stimulated growth of shoots by the extension of internodes. The maximum multiplication coefficient of 7.6 nodes per initial node was obtained using single-node explants on medium with 2 or 4 mg L–1 BAP (Fig. 1a). The coefficient was slightly less in case of two node explants (up to 7.1) and significantly less for three-node explants (up to 5.6) (Fig. 1b). We observed necrosis of tips and top leaves in some shoots growing at 1 mg L–1 BAP regardless of the explant size.
The node position on two- and three-node explants significantly influenced the formation of axillary shoots in vitro (Fig. 1a, b). Axillary shoot formation was highest in distal nodes and reduced toward proximal ones. For example, there was no bud response from base node of three-node explants on medium with 1 mg L–1 BAP at all. However, increasing concentration of BAP stimulated shoot growth from lower axillary buds and percentage of distal nodes in shoot formation decreased from 90.9-64.6% and from 75.8-49.5% for two-node and three-node explants, respectively.
Two most widely used auxins, NAA and IBA, were applied for rooting of common ash shoots on medium with different sugar composition and concentration. The significantly greater percentage (72-93%) of shoots formed roots on 0.5 mg L–1 NAA, as compared to 55-73% in case of 0.5 mg L–1 IBA (Table 2). The influence of sugars on rooting frequency was also significant. The likelihood of rooting was decreased on medium with 5 g L–1 sucrose in comparison with other treatments. The average number of roots per shoot was also maximal on medium with NAA, supplemented with 30 g L–1 sucrose or 10 g L–1 glucose (2.3 roots per shoot).
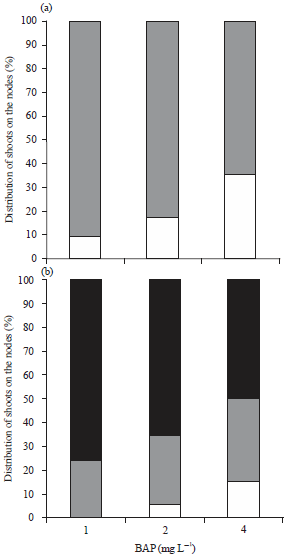 | |
| Fig. 1(a-b): | Effect of node position on shoot formation of common ash, (a) Two-node explants and (b) Three node explants. All media also contained (0.1 mg L–1) IAA. Proximal nodes white, medial nodes hatched and distal nodes grey |
| Table 1: | Effect of explant type on proliferation of common ash in vitro* |
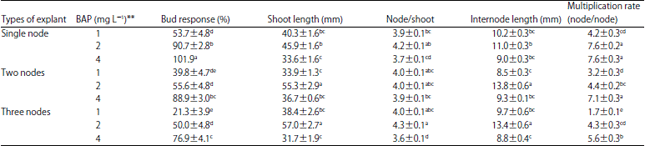 | |
| *values are Means±standard error, **All treatments contain 0.1 mg L–1 IAA, Values followed by different letters are significantly different at p = 0.05 | |
| Table 2: | Effects of auxins and carbohydrates on rooting of common ash shoots (in 4 weeks) |
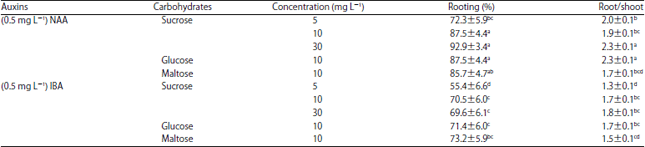 | |
| Values followed by different letters are significantly different at p = 0.05 | |
| Table 3: | Effects of PFD and sucrose concentration on rooting of common ash shoots (in 4 weeks) |
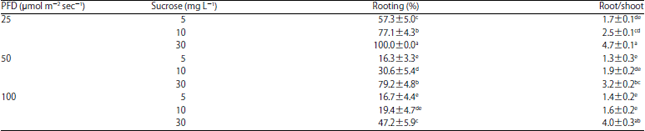 | |
| Values followed by different letters are significantly different at p = 0.05 | |
The shortest roots were obtained on the medium with 10 g L–1 maltose and slightly longer on 5 g L–1 sucrose (data not shown). The medium containing 30 g L–1 sucrose stimulated growth of vigorous shoots with large leaves (Fig. 2a-c).
The root formation was significantly affected by both light intensity and sucrose concentration. The rooting frequency under low light levels (25 μmol m–2 sec–1) was 2-4 times higher than under high light levels (100 μmol m–2 sec–1) (Table 3). Increasing sucrose concentration in the medium significantly improved rooting and raised root number of common ash micro shoots. The maximum rooting (100%) and highest number of roots per shoot (4.7) were obtained at PFD of 25 μmol m–2 sec–1 on the medium with 30 g L–1 sucrose. Unlike the root induction, the root number was only marginally affected by the light intensity. As in the previous experiment, we observed shortest roots (up to 10-15 mm) on medium with 5 g L–1 sucrose, whereas the mean root lengths on 10 and 30 g L–1 sucrose were about 20 and 30 mm, respectively (data not shown). Rooting medium containing 30 g L–1 sucrose also stimulated shoot growth.
Dark treatment during 7 days had significant positive effect on the root induction in 2 and 3 weeks on rooting medium, whereas there was no significant difference in 4 weeks (Table 4). However, 18.6% of dark-treated shoots demonstrated growth in 4 weeks in culture while only 3.9% of shoots rooted without dark treatment.
Plants rooted on medium with 10 and 30 g L–1 sucrose and 10 g L–1 glucose demonstrated significantly higher survival rates in 3 months in the greenhouse as compared to 5 g L–1 sucrose and 10 g L–1 maltose (Table 5). Rooting medium with 10 g L–1 sucrose induced the longest shoots (26.6 mm) in 2 months of acclimatization.
 | |
| Fig. 2(a-f): | Micropropagation of common ash, (a) Multiplication stage, (b, c) Rooting stage, (d) Acclimatization stage, (e) Growing of acclimatized plants and (f) Two years old plants |
| Table 4: | Effect of dark treatment on rooting of common ash shoots |
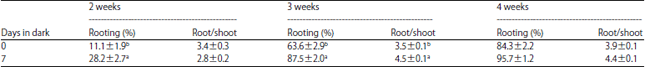 | |
| Values followed by different letters are significantly different at p = 0.05 | |
| Table 5: | Influence of carbohydrates in rooting media on acclimatization of common ash |
 | |
| Values followed by different letters are significantly different at p = 0.05 | |
Sucrose and PFD levels during rooting had no significant effect on the subsequent acclimatization the survival rates were very high (98-100%) in all the treatments (Table 6, Fig. 2d-e). Growth of shoots in the greenhouse significantly stimulated rooting on medium with 30 g L–1 sucrose regardless of intensity of light treatments. After 2 months, tallest plants (61.2 mm) were obtained with the shoots growing in vitro at 25 μmol m–2 sec–1.
As in the previous experiment, because of high survival rates (96-100% in all treatments) it was impossible to determine the effect of tested substrates and dark period during rooting in vitro (Table 7). We only observed large differences in the shoot heights in 2 months of acclimatization. In most treatments, dark-treated plants exhibited the higher increase in growth.
| Table 6: | Influence of PFD and sucrose concentrations during rooting on acclimatization of common ash |
 | |
| Values followed by different letters are significantly different at p = 0.05 | |
| Table 7: | Influence of dark treatment during rooting and substrate on acclimatization of common ash |
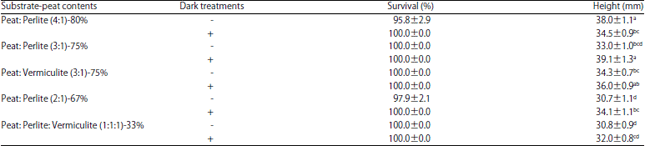 | |
| Values followed by different letters are significantly different at p = 0.05 | |
The substrates containing peat and perlite at 3:1 or 4:1 ratios produced the tallest plants (38-39 mm), whereas the smallest plants (less than 31 mm) were obtained on the substrates containing decreased content of peat (2:1; peat: perlite or 1:1:1; peat: perlite: vermiculite). Replacement of perlite with vermiculite had no effect. All the micropropagated plants of common ash exhibited normal phenotype (Fig. 2f).
DISCUSSION
The explant size is an important factor in plant tissue culture. Generally, the larger explants more responsive in vitro probably because they contain more nutrients and plant growth regulators. In order to determine the optimal size of the common ash explants for best multiplication rate, we compared single, two and three node shoot segments in vitro with different BAP concentrations (1, 2, or 4 mg L–1) in combination with 0.1 mg L–1 IAA. The node density per glass jar was identical in all treatments to simplify further normalization of data to medium volume or cultural vessel. These parameters are more essential and practical for economic evaluation of large-scale micropropagation than the multiplication rate per explant. MS medium was used as it had an advantage over DKW18 and WPM media in our previous experiments13. Our results showed that increase in the explant size leads to a significant decrease in the percentage of responding axillary buds. BAP stimulated proliferation and on medium containing 4 mg L–1 BAP single node explants produced shoots not only from axillary buds but also from dormant buds. The mean proliferation coefficient for single node explants was 1.3 and 1.7 times higher than for two node and three node explants, respectively. This observation confirms that the larger size of explant is not always an advantage. El Boullani et al.19 evaluated proliferation of globe artichoke among the four shoot size categories (<1 cm, 1-1.5 cm, 1.5-2 cm and >2 cm) and the highest rate was obtained for the ones of 1-1.5 cm in size. In our study, the shoot height was maximal on medium with 2 mg L–1 BAP and significantly decreased at 1 and 4 mg L–1 BAP regardless of the explant size. Differences between treatments in number of nodes per shoot were small and the shoot length generally correlated with the internode length. Thus, the concentration of BAP had no effect on the node number on axillary shoots and the new growth and combination of 2 mg L–1 BAP and 0.1 mg L–1 IAA was optimal for growth of common ash shoots by the extension of internodes of explants of all sizes.
We observed that first axillary shoots grow from the distal buds of two and three node explants. For example, the proximal buds of three-node explants at minimal concentration of BAP (1 mg L–1) did not produce shoots at all. These buds became more responsible to shoot formation only at increased BAP concentration. It is known that the position of explant may have a strong influence on organogenesis (topophysic effect). Our data contrast with previously reported results for in vitro propagation of eucalypt Corymbia hybrid where the first four or five nodes of seedlings were equally capable of shoot proliferation in organogenic culture20. It is possible that in addition to the endogenous hormonal balance of plant, this effect is also associated with the specificity of transport of exogenous cytokinins and auxins in explants in vitro.
The highest multiplication rate of common ash, 7.6 nodes per initial node, was obtained from single node explants cultivated on media with 2 or 4 mg L–1 BAP. Similarly, in the work of Hamad and Taha21 reported that separated shoots of pineapple had higher shoot formation per liter of solidified medium and a lower cost per shoot than cluster of shoots at an equal density of shoots per jar. Since high concentrations of cytokinins can induce somaclonal variations, hyperhydricity, etc., 2 mg L–1 BAP concentration is the most optimal for mass propagation of common ash using single-node explants.
Adventitious root formation is a difficult step in micropropagation of many woody species, which may limit the success of this technology. This process is influenced by different factors, primarily, such as kind and concentration of auxins, type and concentration of carbohydrates and light. We evaluated effects of all these factors on the rooting of common ash microshoots and their subsequent acclimatization to autotrophic conditions. Rhizogenesis of microcuttings were differently affected by auxin type and carbohydrates. As in our previous work13, using NAA was more preferable than IBA with respect to rooting frequency and the root number per shoot. Although both IBA and NAA induced similar rooting percentages of F. pennsylvanica shoots, but roots on medium with NAA were thickened and short (less than 5 mm in length)22. Our results demonstrate the advantage of NAA as auxin for common ash rooting in vitro. The rooting frequency on media with 5 g L–1 sucrose was significantly less than in other treatments. We observed high root formation rate on media supplemented with maltose, although the root length was shortest. Our results are in contrary with studies of fruit trees (pear, apple), where shoots failed to root when maltose was used23,24. Lack of rooting on medium with maltose may be attributed to the absence or inactivation of enzymes metabolizing this carbon source. Probably, these enzymes are present in common ash plants.
It is known, that high production costs of woody plant species is caused by a number of factors, most of which are related to the heterotrophic or photomixotrophic nature of plant growth in tissue culture25. In order to enhance photosynthesis in plants in vitro (i.e., photoautotrophic growth) we reduced concentration of sucrose in the medium and enhanced light intensity during rooting of common ash. At the same time, we evaluated the effect of increased sucrose level as an energy source. The root formation of microshoots was considerably affected by light level and sucrose concentration. The rooting frequency had positive correlation with sucrose concentration and negative correlation with light intensity. It was significantly lower at the highest light levels than at the other levels. The best overall rooting (100% in 4 weeks) was achieved on medium with 30 g L–1 sucrose under PFD of 25 μmol m–2 sec–1. In agreement with our results, Mascarello et al.26 reported that rooting percentage of Myrtus communis L. increased significantly in the course of the decrease in light level from 100 to 50 and 25 μmol m–2 sec–1.
In our work, the medium with 30 g L–1 sucrose provided the highest rooting rate of common ash shoots. Use of this concentration also resulted in the highest root number per shoot. These data do not support the well-known opinion that the reduction of sugar content on medium may improve rooting. Our results are consistent with studies conducted both on fruit and forest trees, where beneficial effect of high concentration of carbohydrates (30 g L–1) on rooting was reported24,5. We did not use sucrose concentrations above because higher concentrations reduce economic efficiency of mass propagation.
The root formation of many woody species can be improved by dark incubation during the first phase of rooting. This method was also used for Fraxinus species with different results. In a study of Du and Pijut27, darkness was a key factor for rooting of F. pennsylvanica microplants since shoots did not form roots without the 10-day dark treatment. Pierik and Sprenkels5 showed that a 4-day dark period was required to rapidly obtain 100% rooting of common ash. On the other hand, Navarrete et al.28 reported that incubation of F. americana microshoots for 8 days in darkness was slightly more effective than illuminated culture for rooting. We observed enhanced root formation of dark-treated shoots in 2 and 3 weeks of rooting, but then the differences were not significant. However, one-week dark period stimulated growth of common ash shoots. Probably, the effect of dark period on rooting of woody plants is species-dependent.
The survival rate of plants during acclimatization is the most important parameter, which determines the efficiency of mass Micropropagation. Our results demonstrate that the type and concentration of carbon source in rooting media are important factors affecting plant survival rate, as well as the shoot growth of common ash ex vitro. Survival rate for shoots rooted on media containing 10 g L–1 glucose, 10 or 30 g L–1 sucrose was significantly higher (64-78%) than for the ones rooted in the presence of 5 g L–1 sucrose (39%) and 10 g L–1 maltose (26%). Thus, although unlike some other woody species24,23 F. excelsior formed roots on medium with maltose, but rooted shoots had low viability. Low survival rate of plants of two treatments in the greenhouse correlated with smallest root number and shortest roots of the shoots in vitro. Such correlation was not observed for plants that demonstrated high survival rate. Probably, the number of roots per shoot and the root length, after they reach certain values, become less important for survival.
In our other acclimatization experiments, determination of influence of such factors as light intensity, sucrose concentration, dark treatment during in vitro culture and substrate composition, was impossible due to very high survival rate under greenhouse conditions (96-100%). These differences between the two groups of experiments can be explained by the season effect. All plants were acclimatized in the greenhouse with a controlled climate, but the former group was planted during autumn-winter months, while the latter group was planted during the spring-summer season. The sucrose concentration in the media had significant effect on shoot growth ex vitro. The highest plants were obtained after rooting on media with 30 g L–1 sucrose regardless of the light intensity. We observed enhanced growth and large leaves of shoots rooted in the presence of 30 g L–1 sucrose under in vitro conditions. Our data are in agreement with a study of Premkumar et al.29, where the vigor of the acclimatized avocado plants treated with 30 and 50 g L–1 sucrose was clearly higher as compared to plants treated with 5 g L–1 sucrose. The plants from the treatments with higher sucrose concentrations displayed a higher concentration of non-structural carbohydrates and relatively higher content of Rubisco protein, that could be beneficial for the subsequent growth of the acclimatized plantlets29.
It is known that such substrates as perlite, vermiculite, peat moss and the mixture of peat moss with perlite provided highest survival rates and a tendency to increased plant growth during acclimatization30. For all substrate compositions tested, except for a mixture of peat and perlite (4:1), the dark-treated during in vitro rooting common ash plants were higher in 2 months after acclimatization. The maximum growth of plants was observed at substrates with maximum content of peat (mixtures of peat and perlite in 4:1 and 3:1 ratios). Our results are consistent with the data of Markovic et al.31, which showed that the percentage of acclimatized Dianthus deltoides L. plants planted in the 1:1 peat and sand substrate mixture turned out to be the lowest (82.7%), while the ones planted in a 4:1 ratio mixture of peat and sand revealed the most successful acclimatization (97%). To our knowledge comparison of various substrates during acclimatization of F. excelsior plants was not carried out earlier.
In the present study, it was shown that single-node explants are the most optimal for multiplication of common ash. Rooting medium containing 30 g L–1 sucrose and PFD of 25 μmol m–2 sec–1 are the best combination for root formation of microshoots and subsequent acclimatization under ex vitro conditions. Almost 100% of plant survival in vitro in the greenhouse was achieved. In summary, an efficient technology for mass propagation of F. excelsior plants was developed via optimization of a number of parameters. This technology can be used both for large scale plant production for forest plantations and for rapid multiplication of unique genotypes.
ACKNOWLEDGMENTS
This work was supported by the Ministry of Education and Science of The Russian Federation (Project № 14.613.21.0007 from 26.08.2014, unique identifier RFMEFI61314X0007).
REFERENCES
- Van Sambeek, J.W. and J.E. Preece, 2007. In vitro Propagation of Fraxinus Species. In: Protocols for Micropropagation of Woody Trees and Fruits, Jain, S.M. and H. Haggman (Eds.). Chapter 17, Springer, Berlin, Germany, ISBN: 978-1-4020-6351-0, pp: 179-192.
Direct Link - Chalupa, V., 1990. Micropropagation of hornbeam (Carpinus betulus L.) and ash (Fraxinus excelsior L.). Biol. Plant., 32: 332-338.
CrossRefDirect Link - Hammatt, N. and M.S. Ridout, 1992. Micropropagation of common ash (Fraxinus excelsior). Plant Cell Tissue Organ. Cult., 31: 67-74.
Direct Link - Mitras, D., P. Kitin, I. Iliev, D. Dancheva and A. Scaltsoyiannes et al., 2009. In vitro propagation of Fraxinus excelsior L. by epicotyls. J. Biol. Res.-Thessalon., 11: 37-48.
Direct Link - Schoenweiss, K. and A. Meier-Dinkel, 2005. In vitro propagation of selected mature trees and juvenile embryo-derived cultures of common ash (Fraxinus excelsior L.). Propag. Ornam. Plants, 5: 137-145.
Direct Link - Capuana, M., G. Petrini, A. Di Marco and R. Giannini, 2007. Plant regeneration of common ash (Fraxinus excelsior L.) by somatic embryogenesis. In vitro Cell. Biol.-Plant, 43: 101-110.
CrossRefDirect Link - Capuana, M., 2013. In vitro propagation of ash (Fraxinus excelsior L.) by somatic embryogenesis. Methods Mol. Biol., 994: 213-221.
CrossRefPubMedDirect Link - Dancheva, D. and I. Iliev, 2015. Factors affecting adventitious shoot formation in Fraxinus excelsior L. Propag. Ornam. Plants, 15: 10-20.
Direct Link - Du, N. and P.M. Pijut, 2009. Agrobacterium-mediated transformation of Fraxinus pennsylvanica hypocotyls and plant regeneration. Plant Cell Rep., 28: 915-923.
CrossRefPubMedDirect Link - Stevens, M.E. and P.M. Pijut, 2014. Agrobacterium-mediated genetic transformation and plant regeneration of the hardwood tree species Fraxinus profunda. Plant Cell Rep., 33: 861-870.
CrossRefPubMedDirect Link - Palla, K.J. and P.M. Pijut, 2015. Agrobacterium-mediated genetic transformation of Fraxinus americana hypocotyls. Plant Cell Tissue Organ Cult., 120: 631-641.
CrossRefDirect Link - Lebedev, V. and K. Schestibratov, 2013. Effect of natural and synthetic growth stimulators on in vitro rooting and acclimatization of common ash (Fraxinus excelsior L.) microplants. Nat. Sci., 5: 1095-1101.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Lloyd, G. and B. McCown, 1980. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. Proc. Int. Plant Propagator's Soc., 30: 421-426.
Direct Link - El Boullani, R., A. Elmoslih, A. El Finti, A. El Mousadik and M.A. Serghini, 2013. Effect of decapitation and size of explants on in vitro multiplication rate of globe artichoke. Acta Hort., 983: 325-329.
CrossRefDirect Link - Hung, C.D. and S.J. Trueman, 2011. Topophysic effects differ between node and organogenic cultures of the eucalypt Corymbia torelliana x C. citriodora. Plant Cell Tissue Organ. Cult., 104: 69-77.
CrossRefDirect Link - Hamad, A.M. and R.M. Taha, 2009. Effect of explants density on the in vitro proliferation and growth of separated and cluster shoots of smooth cayenne pineapple (Ananas comosus L. Merr.). Asian J. Plant Sci., 8: 313-317.
CrossRefDirect Link - Kim, M.S., N.B. Klopfenstein and B.M. Cregg, 1998. In vitro and ex vitro rooting of micropropagated shoots using three green ash (Fraxinus pennsylvanica) clones. New For., 16: 43-57.
CrossRefDirect Link - Kadota, M. and Y. Niimi, 2004. Influences of carbon sources and their concentrations on shoot proliferation and rooting of 'Hosui' Japanese pear. HortSci., 39: 1681-1683.
Direct Link - Bahmani, R., O. Karami and M. Gholami, 2009. Influence of carbon sources and their concentrations on rooting and hyperhydricity of apple rootstock MM.106. World Applied Sci. J., 6: 1513-1517.
Direct Link - Kozai, T., Y. Xiao, Q.T. Nguyen, F. Afreen and S.M.A. Zobayed, 2005. Photoautotrophic (sugar-free medium) micropropagation systems for large-scale commercialization. Propag. Ornam. Plants, 5: 23-34.
Direct Link - Mascarello, C., R. Melis, E. Mantovani and B. Ruffoni, 2009. Evaluation of the role of light intensity and hormonal treatments during in vitro rooting of myrtus micropropagated plantlets related to acclimatization performances. Acta Hortic., 812: 379-386.
CrossRefDirect Link - Du, N. and P.M. Pijut, 2008. Regeneration of plants from Fraxinus pennsylvanica hypocotyls and cotyledons. Scientia Hortic., 118: 74-79.
Direct Link - Navarrete, N.E., J.W. Van Sambeek, J.E. Preece and G.R. Gaffeny, 1989. Improved Micropropagation of White Ash (Fraxinus americana L.). In: Seventh Central Hardwood Forest Conference, Rink, G. and C.A. Budelsky (Eds.)., USDA Forestry Service, St. Paul, MN., USA., pp: 146-149.
Direct Link - Premkumar, A., A. Barcelo-Munoz, F. Pliego-Alfaro, M.A. Quesada and J.A. Mercado, 2002. Influences of exogenous sucrose on juvenile avocado during in vitro cultivation and subsequent ex vitro acclimatization. Trees, 16: 569-575.
CrossRefDirect Link - Jimenez, V.M., E. Guevara and S. Masis, 2011. Effect of macronutrients and sucrose concentration on in vitro growth of Drosera capensis L. (Droseraceae) plants and evaluation of six substrates for acclimatization. Propag. Ornam. Plants, 11: 34-39.
Direct Link - Markovic, M., M. Popovic and D. Vilotic, 2013. Micropropagation of Dianthus deltoides L. through shoot tip and nodal cuttings culture. Arch. Biol. Sci., 65: 17-22.
CrossRefDirect Link