
Mohammed S. El-Hersh
Microbial Activity Unit, Department of Microbiology, Soils, Water and Environment Research Institute, Agricultural Research Center, P.N. 12619, Giza, Egypt
Wesam I.A. Saber
Microbial Activity Unit, Department of Microbiology, Soils, Water and Environment Research Institute, Agricultural Research Center, P.N. 12619, Giza, Egypt
Husain A. El-Fadaly
Department of Microbiology, Faculty of Agriculture, Damietta University, Egypt
Biotechnology
Year: 2014 | Volume: 13 | Issue: 6 | Page No.: 252-262







ABSTRACT
Bacillus subtilis ATCC 11774 was exploited to produce amino acids on a medium containing olive cake as non-traditional carbon and nitrogen source. Initially, the potential proteolysis of B. subtilis ATCC 11774 was sequentially optimized for protease production based on statistical strategy. In which, the two level Plackett-Burman design revealed peptone and olive cake as the significant factors affecting protease production. Optimization of both constituents in addition to pH using central composite design maximized protease production (30.3 U mL-1 min-1) by an overall 4.59-fold increase as compared to the non-optimized (6.6 U mL-1 min-1) medium, with marked evidence for the biosynthesis of total free amino acids, being 3403 μg mL-1, accompanied with the reduction in final culture pH down to 4.3. The differentiation of the filtrate of optimized protease production medium using amino acid analyzer revealed glutamic acid (1185.9 μg mL-1) as the highest synthesized amino acid. Other minor amounts of amino acids were also detected. Interestingly, this is the first report on the production of amino acids from olive cake-based medium.
PDF Abstract XML References Citation
Received: September 22, 2014;
Accepted: October 28, 2014;
Published: May 04, 2019
How to cite this article
Mohammed S. El-Hersh, Wesam I.A. Saber and Husain A. El-Fadaly, 2014. Amino Acids Associated with Optimized Alkaline Protease Production by Bacillus subtilis ATCC 11774 Using Statistical Approach. Biotechnology, 13: 252-262.
DOI: 10.3923/biotech.2014.252.262
URL: https://scialert.net/abstract/?doi=biotech.2014.252.262
DOI: 10.3923/biotech.2014.252.262
URL: https://scialert.net/abstract/?doi=biotech.2014.252.262
INTRODUCTION
Amino acids are important group of organic compounds, required for building cells, tissues and organs. There are two categories of amino acids: Non-essential amino acids and the essential amino acids that cannot be synthesized de novo (from scratch) by the organism and therefore must be supplied in its diet for protein synthesizing or general homeostasis. The discrimination between essential and non-essential amino acids is not fully clear, as some amino acids can be produced from others. For instance, the sulfur-containing amino acids, methionine and homocysteine, can be converted into each other but neither can be synthesized de novo in humans (Harper et al., 1984; Fernstrom, 1990; Blomstrand, 2001; Ventura et al., 2012; Falade and Adewusi, 2013). That is in turn, disclose the importance of both groups to the organism.
Proteases play the central role in this respect, since the production of amino acids and other nitrogenous compounds are mainly directed by the action of several kinds of proteases (EC, 3.4.21.24). Microbial proteases are one of the most widely studied group of enzymes and gaining more importance than conventional chemicals that cleave peptide, because of the cheaper production cost and use of renewable resources, they have wide applications e.g., in several industries of food and pharmaceutical. Large proportion of commercially available alkaline proteases are derived from Bacillus strains (Kumar and Takagi, 1999). Synthesis of bio-products by microorganisms is greatly influenced by medium components and physical factors (Potumarthi et al., 2007). For the economization of the production process, cheap raw materials and overproduction are essential, the latter can be achieved both by genetic manipulation, that may not be stable and by media engineering, which is the better alternative for the economic overproduction, especially, when using cheap sources of carbon and nitrogen.
Olive Cake (OC) is an inexpensive biomass that is accumulated in large quantities as solid residue after olive oil extraction, representing a serious environmental problem. Besides, OC is considered a rich source of a wide array of biologically active compounds, which may induce the microbial growth, as well as formation of various compounds (Rodis et al., 2002; Alu'datt et al., 2010) e.g., amino acids. Therefore, reducing the environmental impact of OC and/or harness its potential economic value is of great importance. One of the possible approaches is using OC as a main nutritional component combined with other factors to maximize protease production for the biosynthesis of amino acids as the final target.
In order to optimize the fermentation process in biotechnology, a number of optimization techniques could be used for understanding of production kinetics. The statistical approaches offer ideal ways in this respect (Gupta et al., 2002). In preliminary studies, the Plackett-Burman designs allow for the screening of main factors from a large number of process variables, where, the principal objective is to select variables that can be fixed or eliminated in further optimization processes (Reddy et al., 2008). As well as, Response Surface Method (RSM) is one of the popularly used optimization procedures, mainly developed based on full factorial Central Composite Design (CCD). RSM helps identify the effective factors, study interactions, select optimum conditions and quantify the relationships between one or more measured responses and the vital input factors using limited number of experiments. Central composite designs contain three groups of design points; two-level factorial or fractional factorial, axial and central points, to estimate the coefficients of the Quadratic Model (Oskouie et al., 2008).
Hence, the present investigation aimed to study the optimization of extracellular alkaline protease production, as indicator for amino acids synthesis by Bacillus subtilis ATCC11774, on cheap medium components containing olive cake as a new substrate, using CCD of RSM and differentiation of amino acids associated with optimized process of protease production.
MATERIALS AND METHODS
Inoculum preparation and fermentation medium: Bacilllus subtilis ATCC® 11774™ was obtained from American Type Culture Collection (ATCC), Illinois, USA. The bacterial strain showed proteolytic activity during growth on milk agar. The stock culture of the bacterium strain was stored in 30% glycerol at -7°C. Prior to each experiment, the bacterium was sub cultured from the frozen stocks onto nutrient agar medium. Bacterial cells from 24 h aged culture slants were scraped using sterile distilled water. The final inoculum concentration was adjusted to count 108 CFU mL-1.
The composition of the proposed submerged liquid state fermentation medium contained (g L-1), glucose (18), peptone (8), casein (4), KH2PO4 (2), olive cake (4) and corn steep liquor (CSL) (4), distributed in 250 mL Erlenmeyer flasks, each with 45 mL of the fermentation medium. The initial medium pH was adjusted to 7, then sterilized at 121°C for 15 min. After cooling, flasks containing media were inoculated with 5 mL of the previously prepared inoculum and incubated at 35°C, with rotary shaking at 150 rpm. The incubation period was determined based on the time course profile. The fermented medium of each trial was, then, centrifuged at 5000 rpm for 15 min. The resultant supernatant was tested for protease, Total free Amino Acids (TAA) and final culture pH.
Fermentation designs
Plackett-burman design: The six independent variables of medium components (glucose, peptone, casein, KH2PO4, olive cake and CSL) were screened at high (+1) and low (-1) levels as well as the center points (0), using the two-level Plackett-Burman design (Table 1) in twelve combinations plus three center points (Plackett and Burman, 1946). The design was performed in triplicates and the average of protease activity and final culture pH were treated as responses.
| Table 1: | Plackett-Burman design with actual and codified values of medium components and corresponding protease production and final pH by B. subtilis ATCC 11774 |
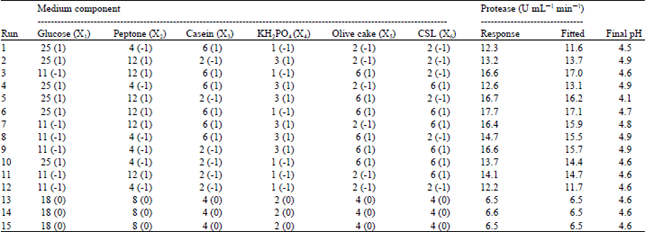 | |
| Number between parentheses is the corresponding coded value | |
The main effect of each variable was simply calculated as the difference between the average of measurements made at the high level (+1) and the average of measurements observed at the low (-1) level of that factor, using the following first order model equation:
where, Xi is the effect of the tested variable, Mi+1 and Mi-1 represent variable response from the trials where the variable (Xi) measured was present at high and low concentration, respectively and N is the total number of trials.
Central composite design: Fermentation factors affecting protease production were optimized using the full Central Composite Design. The significant variables (peptone, X2 and olive cake, X5) determined based upon Plackett-Burman experiment, in addition to the initial pH (X7) of the culture medium were further investigated for studying the interaction among the three variables. The other medium components were omitted from the fermentation medium. Each of the three factors was examined at five different levels at the center point and an axial point located at a specified distance (α = 1.682) from the design center in each direction on each axis, representing 5 levels for each factor. According to the applied design, 20 combinations were executed, the actual and code levels are shown in Table 2 and the observations of the three factors were fitted to the following second order polynomial quadratic model:
where, Y is the predicted response, β0 model constant, Xi and Xj independent variables; βi is linear coefficients; βij is cross product coefficients and βii is the quadratic coefficients.
Determinations
Protease assay: The proteolytic activity was quantitatively assayed in a mixture consisted of 1 mL culture supernatant as a crude protease and 1 mL substrate solution and (2% casein in 0.1 M tris-HCl buffer at pH 7.5), then incubated at 37°C for 20 min. The released amino acids were determined following the method of Lupin et al. (1982). One unit (U) of protease activity was defined as the amount of the enzyme resulting in the release of 0.5 μg of tyrosine equivalent min-1 under the assay conditions, using tyrosine as a standard.
Determination of total free amino acids: Two milliliter of culture supernatant was added to 2 mL of 10% TCA followed by centrifugation at 5000 rpm for 15 min to obtain clear filtrate. The TAA was determined following the method of Lupin et al. (1982). Another set of trials was used as controls, which were carried out using non-inoculated media subjected to the same fermentation conditions. After determination of TAA, each control was subtracted from its corresponding trial sample. Tyrosine was used as a standard.
Quantitative analysis of individual amino acids using amino acid analyzer: For quantitative differentiation of individual amino acids, sample of clear filtrate obtained from optimized fermented medium was determined according to the method described by Fontaine and Eudaimon (2000), using the amino acid analyzer Sykam GmbH, Eresing, Germany, to estimate the type and concentration of each amino acid.
| Table 2: | Coded and actual values of the three independent variables used in central composite design matrix with experimental and fitted values of protease production |
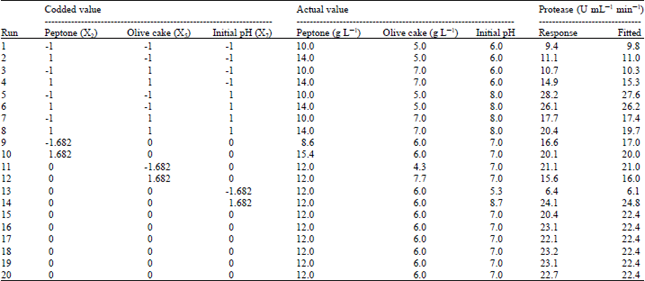 | |
Non-inoculated medium with the same composition, subjected to the same fermentation conditions, was also injected into the amino acid analyzer to serve as control, then subtracted from its corresponding trial sample.
Statistical analysis: All experiments were performed in triplicates and the average of data was calculated. The statistical analysis of the results was performed with the aid of statistical software packages Minitab (version 17, Minitab Inc., USA) and Design Expert (version 7, State-Ease, USA).
RESULTS
Among several strains, B. subtilis was found to be the potential source of protease (data not shown), in which B. subtilis ATCC 11774 was qualitatively tested for potential proteolysis, as the first step in amino acids production. The bacterial strain showed activity during growth on milk agar plates, in which the clear zone of 24 h old culture was 20 mm in diameter.
Time course of bioconversion process: Accordingly, the proteolytic activity, TAA production and the final culture pH of the selected strain were evaluated periodically, to quantitatively determine the optimum incubation period. Figure 1 shows that protease and TAA biosynthesis were detected earlier after 12 h of incubation on broth medium. Maximum productions were observed after 48 h of incubation by the tested bacterial strain being 6.6 U of protease and 1348.1 μg mL-1 of TAA and then continually reduced to the end of fermentation period was recorded. On the other side, the final pH showed opposite trend. The simple correlation coefficient (R, at p<0.05) reveals that the final culture pH showed significant negative correlation with both protease production (r = -0.947) and TAA (r = -0.963). Oppositely, significant positive correlation was detected between protease production and TAA (r = 0.985). This data suggests the dependency of TAA production, mainly on proteolytic activity, rather than the inter-transformation of various cell metabolites. Therefore, the next steps considered protease production and reduction in the final culture pH as indicator for TAA accumulation.
Screening the medium components using Plackett-Burman: Plackett-Burman design has been adopted at two levels for screening the potential importance of six variables on protease production and final pH. The design matrix selected for the screening of significant variables for reduction in final culture pH and protease production and the corresponding responses are shown in Table 1, in which, a high degree of similarity was observed between the predicted (fitted) and experimental (response) values of protease. The run number 6 recorded the highest protease production (17.7 U). The adequacy of the protease production and final culture pH models were calculated and the variables evidencing statistically significant effects were screened by ANOVA (Table 3). The main effect of protease recoded significant p-value (0.003).
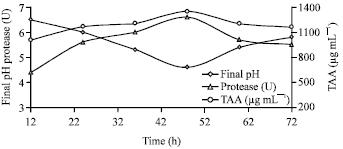 | |
| Fig. 1: | Profile of protease, TAA and final culture pH by B. subtilis ATCC 11774 during 3 days incubation period |
| Table 3: | Estimated effect, regression coefficient and ANOVA of Plackett-Burman matrix of medium components for the resulted protease and reduction in final culture pH by B. subtilis ATCC 11774 (coded units) |
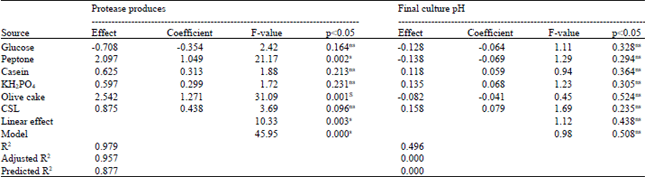 | |
| s: Significant, ns: Not significant at p<0.05 | |
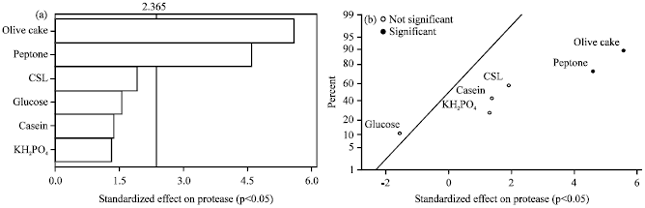 | |
| Fig. 2(a-b): | (a) Pareto chart and (b) Normal plot of the standardized effects of the different medium components on the protease production by B. subtilis ATCC 11774 |
Olive Cake and peptone with p-values of 0.001 and 0.002, in sequence, were determined to be the most significant factors, the overall model also showed significant p-value. That was confirmed by the high values of coefficient of determination (R2), adjusted R2 and predicted R2, being 0.979, 0.957 and 0.877, respectively, indicating the validity of the data, subsequently their model. The values of predicted R2 and adjusted R2 are in reasonable agreement. On the other hand, the final culture pH reduced, vigorously, to the acidic side, but no dependency could be rely on such criteria, because of the insignificant model and low values of different kinds of R2.
The magnitude of an effect of different tested medium components on protease production was estimated using Pareto chart (Fig. 2a). The chart displays the absolute value of the effects and a reference line. Effects of olive cake and peptone showed potential importance, since both of them extended past the reference line. The normal plot of the effects (Fig. 2b) shows the importance of the tested variables, in which variables that do not fall near the fitted line (olive cake and peptone) are more important than variables centered around zero (CSL, glucose, casein and KH2PO4), which tend to be smaller than the significant ones. Only glucose exerted a negative effect, whereas the other variables exerted positive effects on protease production. So, the insignificant variables were eliminated from the fermentation medium. The optimum levels of the two significant variables (olive cake and peptone), which recorded positive effect were further determined by the Central Composite Design (CCD) of RSM design.
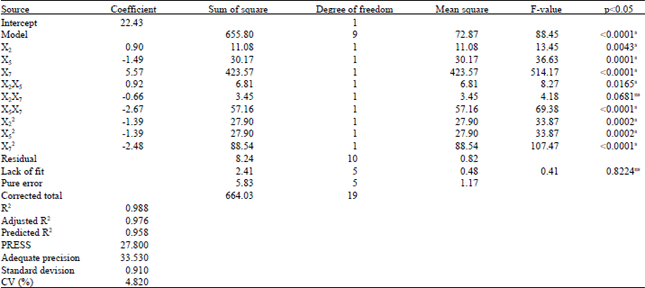
Quadratic response surface optimization of protease production: The CCD was adopted to explain the interaction among the three independent variables; viz., peptone and olive cake, selected based on Plackett-Burman screening experiment, in addition to the initial culture pH, on protease production by B. subtilis. The response and fitted protease production as a function of CCD experiments are presented in Table 2. The run number 5 was the highest one. The data was fitted using the second order polynomial equation. The analysis of variance (Table 4) indicates that the model terms of X2, X5, X7, X2, X5, X5, X7, X22, X52 and X72 were significant (p<0.05), but the interaction of X2X7 was not significant.
The model recorded significant F-value (88.45), with high values of R2 (0.988), adjusted R2 (0.976) and predicted R2 (0.958), indicating the validity of the data. The values of predicted R2 and adjusted R2 are in reasonable agreement. In addition, the model has an adequate precision value of 33.5. The lack of fit F-value (0.41) is not significant relative to the pure error. The model shows standard deviation of 0.91, meaning low variation or dispersion from the average of protease production. Coefficient of Variation (CV), which describes the variation of a testas a percentage of the mean recorded 4.82%. Finally, the model could be effectively used to measure the particular model fits at each point in the design, since the predicted residual sum of squares (PRESS) values lowered to 27.80. The parity plot (Fig. 3) confirms a satisfactory correlation between the experimental and predictive values of protease production.
In order to determine the optimal levels of each variable for maximum protease production, three-dimensional response surface graphs were constructed by plotting the protease production on the z-axis against any two independent variables, while maintaining the other variables at their central levels. As is shown in Fig. 4a, an increase in protease secretion was observed when the peptone and olive cake concentrations were increased, then concomitant decline in enzyme production was observed.
| Table 4: | Analysis of variance (ANOVA) for response surface quadratic model for production of protease (coded units) |
 | |
| s: Significant; ns: Not significant at p<0.05 | |
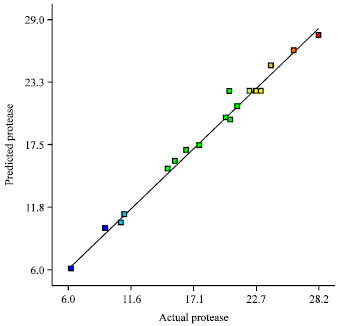 | |
| Fig. 3: | Parity plot showing the distribution of experimental vs. predicted values of protease production |
A linear profile was observed in Fig. 4b, as the peptone concentration and initial culture pH values increase. However, as is shown in Fig. 4c with olive cake and pH, protease production could not increase further with increasing olive cake, but increased continiously with the the higher pH values.
Based on ANOVA, X2X7 is the only insignificant model, it is already known that, if there are many insignificant model terms (not counting those required to support hierarchy), model reduction may improve the model. In the present, only one individual model is insignificant, therefore, there is no need to simplify the second-order polynomial equation for protease production. Accordingly, the fitted value was calculated from the final equation in terms of coded values as follows:
Protease (Y) = 22.43+0.9(X2)-1.49(X5)+5.57(X7)+0.92 (X2X5)-0.66(X2X7)-2.67(X5X7)-1.39(X22)-1.39(X52)-2.48(X72)
The experimental data was fitted and the solver was applied to optimize the proteolysis process. The solver was set to maximize protease as response and, at the same time, as indicator for the biosynthesis of amino acids, whereas, the independent variables; peptone was set to be minimized and olive cake and pH within their tested ranges. The optimum levels of each variable were determined by the solver to be 8.64 g L-1 o f peptone, 4.32 g L-1 of olive cake with initial culture pH at 8.7.
According the prediction equation, the fitted value of protease was calculated to be 29.9 U mL-1 min-1. It should be considered that the polynomial model is a reasonable approximation of the true functional relationship on a relatively small region of the entire space of the independent variables. Accordingly, the calculated value of protease production was experimentaly validated and the activity reached 30.3 U mL-1 min-1, recording 1.71 fold increase, incomparison with the highest run (No. 6) in Plackett-Burman experiments.
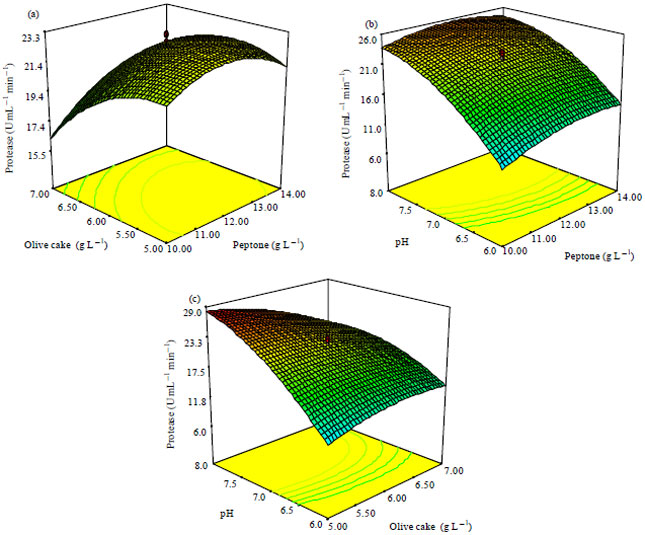 | |
| Fig. 4(a-c): | Three-dimensional response surface plot for protease production showing the interactive effects of (a) Peptone and olive cake at hold value: pH = 7, (b) Peptone and pH at hold value: olive cake = 6 g L-1 and (c) Olive cake and pH at hold value: Peptone = 12 g L-1 |
The total amino acids in the optimized culture was determined, recording 3403 μg mL-1, this was accompanied with reduction in the final pH down to 4.3. So, the differentiation of the amino acids content was performed using amino acid analyzer to discover their profile.
Amino acids profile: The profile of the amino acids associated with optimized protease production were investigated (Fig. 5) by amino acids analyzer. The overall recovered amino acids reached 6801.5 μg mL-1, varied between essential and non-essential categories. The amino acids obtained in descending order were glutamic, leucine, aspartic acid, proline, lysine, glycine, alanine, valine, phenylalanine, isoleucine, threonine, serine, tyrosine, arginine, methionine, cysteine and histidine.
DISCUSSION
Amino acids are valuable component in everyday nutrition; they are extremely required at various concentrations, for nearly every organism in every growth stage. Work on production of amino acids is rare, especially, when using cheap source for the production process. Amino acids are the final products of proteases action, so protease production and reduction in final culture pH are the initial and direct indicators for the release of amino acids, during the several stages of fermentation. Selection of appropriate strain for such mission is the important point. However, on the other hand, the medium manipulation was found to be more stable and effective procedure than genetic manipulation for maximizing the proteolysis by the selected microbe.
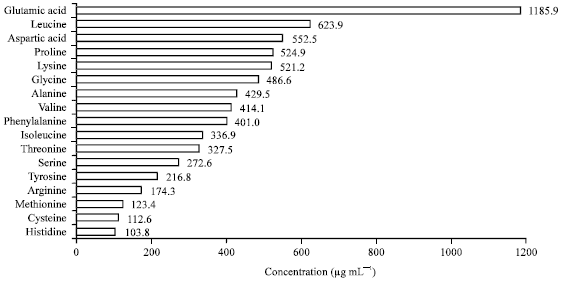 | |
| Fig. 5: | Amino acids profile, recovered by the amino acids analyzer, associated with liquid state fermentation medium by B. subtilis ATCC 11774 |
Herein, B. subtilis is good suitable strain and vital source of these enzymes, subsequently, the production of amino acids. This was obvious on milk agar plates, so, B. subtilis ATCC 11774, was confirmed as proteolytic strain. Time course of the fermentation process by B. subtilis ATCC 11774 revealed 2 days as the best optimum fermentation period; this was true for both protease production and TAA, accompanied with the maximum reduction on final culture pH. Protease production and TAA are positively correlated and both are negatively correlated with final culture pH, these data are expected, because of the catalytic action of protease leads to the formation of amino acids that have acidic nature, leading to the drop in pH. However, 2 days is reasonably accepted in the biotechnology field.
Accordingly, a study has been designed for screening different medium components for proteolytic activity by B. subtilis ATCC 11774. Due to its ready applicability and aptness, the statistical screening, Placket-Burman has been adopted at two levels, to select the significant medium components necessary for enhancing protease production as a direct indicator, as well as reduction in pH as an indirect indicator for the biosynthesis of amino acids. The model term of protease production showed significant model, represented by a large variation in protease production (6.5-17.7 U) from that mandated by the Plackett-Burman design experiments suggested a need for further optimization. On the other side, the insignificant model of the final culture pH may be back to its narrow range (4.1-4.9). This reduction in the final culture pH along all tested runs of Plackett-Burman gives an evidence for the presence of compounds with acidic nature, i.e., the secretion of TAA, along all the 15-tested run, in adequate amounts that led to semi equal reduction of the final pH. This is a good indicator for the biosynthesis of amino acids, but unfortunately, does not allow fair selection of the best variable among the various medium components. The high similarity between the fitted and response values of protease production reflects the accuracy of the design. In addition, the p-value of the main effect recoded 0.003. Both peptone and olive cake were the major factors affecting protease production, by which their p-values being 0.001 and 0.002, respectively. The lower p-value than 0.05 the more significant effect on production of protease. The adjusted R2 and predicted R2 recorded high values in protease production as compared with the final culture pH. R2, adjusted R2 and predicted R2 are measures of how well the fitness of the data. These values can help selection the model with the best fit, the higher of their values, up to 1, the more fitness of the data. The final culture pH was excluded as an evaluation response, because of its high p-values of the model and low values of different kinds of R2, in addition to the insignificant model terms. Only, protease was considered as the selection criterion of medium components, both peptone and olive cake were the significant factors in this respect.
The complex substrate of nitrogen and carbon is better for protease production than simple ones (Kole et al., 1988), so, the complex content of olive cake (organic matter, crude proteins, fibers and amino acids N) (Molina-Alcaide and Yanez-Ruiz, 2008) may affect protease response. However, the other tested factors were less effect, where the data in pareto chart and normal plot of the standardized effects show the negative effect of glucose and the non-significant positive effect of casein. The metabolism of glucose may affect on protease response as catabolic repression (Kole et al., 1988). Contrarily, the best fermented carbon and nitrogen sources, utilized by Mucor mucedo DSM809 for protease production, were glucose and casein (Yegin et al., 2010), the later was found to contain casamino acids, which play a functional role in inducing protease production (Chauhan and Gupta, 2004). In addition, CSL was stimulating for protease production by Bacillus sp. RKY3 (Reddy et al., 2008). Commonly, there is no general medium for protease production by different microbial strains. However, Plackett-Burman, achieved 2.68-fold increase in protease production compared to the initial activity (6.6 U mL-1 min-1). Based upon the previous rules, peptone and olive cake were selected for further optimization studies.
The interaction effect among the significant variables (peptone and olive cake) in addition to the initial culture pH were studied by the CCD. Fitting data, with a second order polynomial function indicates the accuracy of the model. The analysis of variance indicates that all model terms were significant at p<0.05), except the interaction of X2X7, meaning that there was no significant effect to the interaction between peptone and initial pH. Thus, this term could be eliminated from the equation of prediction. High F-value (88.45) implies the significant of the overall model. Data obtained from ANOVA indicate that R2 is 0.988 i.e., the model can accurately estimate 98.8% of variation of protease production that is explained by the factors. The adjusted R2 (0.976) and predicted R2 (0.958) are measures of how well the fitness of the data. These values can help selection the model with the best fit. Predicted R2 is more useful than adjusted R2 for comparing models and can prevent overfitting the model. Overfitting refers to models that appear to explain the relationship between the predictor and response variables for the data set used for model calculation, but the new observations cannot be accurately predicted. Large value of predicted R2 suggests models of greater predictive ability. However, the higher of their values, the more accuracy of the relationships between the variables (factors) and response (protease). The assessment of the model’s predictive ability of PRESS recorded small value of 27.80, in general, the smaller the PRESS value, the better the model’s predictive ability. PRESS is used to calculate the predicted R2, which is generally more intuitive to interpret. Both of predicted R2 and PRESS reduce the model overfitting, since calculation of predicted R2 is based on values outside the model estimation. The value of adequate precision (>4), which is an index of the signal to noise ratio, suggesting that the model can be used to navigate the design space; this is an essential prerequisite for a model to be a good fit, together with the non-significant lack of fit F-value (0.41), as fitting of model is required. The low standard deviation value (0.91) indicates that the protease production points tend to be very close to the mean, also, indicates the low variation or dispersion from the average exists and the protease points are not spread out over a large range of values. The accuracy of the experimentation procedure, presented by the CV (4.82%), gives indication of adequate method performance, since CV of 5.0% or less is good. All the previous statistatical calculations confirmed a satisfactory adjustment of the quadratic model to the experimental data. To ensure the validity of the obtained polynomial model, the parity plot was drawn, wherein, the points clustered of experimental and predictive values of protease production around the diagonal line, indicates the good fit of the model.
The response of protease to pH values could be due to perturbation effects of pH on the gradient intracellular flux distributions, as well as, due to the difference in proton electrochemical generation (Calik et al., 2002). Suganthi et al. (2013) found that the optimum protease production by B. licheniformis TD4 was at pH 8, whereas, earlier study pointed out that the optimum pH for operating condition from a quadratic form of RSM model was 7.6 (Dutta et al., 2004). These results are similar to that obtained in the case production of protease by Bacillus sp. RGR-14 (Chauhan and Gupta, 2004). The response of the bacterial strain to olive cake may be back to the phenolic compounds that have been identified in olive fruits, in which olive oil contains 2%, while 98% is present in olive mill waste (Rodis et al., 2002), suggesting induction of the biosynthesis of microbial products.
The protease production recovered after optimization (30.3 U mL-1 min-1) recorded 4.59-fold increase in comparison with initial value (6.6 U mL-1 min-1) recorded with B. subtilis ATCC 11774. At this point, there was reasonable content of TAA being 3403 μg mL-1. This, in turn, encouraged differentiating and discovering the profile of the amino acids in the crude extract of the fermented culture.
Several amino acids were detected in the filtrate, accompanied with marked reduction in final pH and high activity of protease production. Some of the detected amino acids, regarded as essential for humans, were detected i.e., phenylalanine, valine, threonine, isoleucine, methionine, leucine, lysine and histidine. However, not all essential amino acids were synthesized in the fermented medium by the tested bacterium e.g., tryptophan. Additionally, the filtrate contained some amino acids that are required in specific growth stage, like cysteine (sulphur-containing amino acids), tyrosine (aromatic amino acids) and arginine that are required by infants and growing children (Narayan et al., 2011). Other amino acids (arginine, cysteine, glycine, glutamine, proline, serine and tyrosine) are considered conditionally essential, meaning they must be supplied exogenously in certain cases to specific populations, who do not synthesize them in adequate amounts, i.e., they are not required in normal situation in the diet. An example would be with the phenylketonuria (PKU) disease. Individuals living with PKU must keep their intake of phenylalanine extremely low to prevent metabolic complications including mental retardation; these patients require tyrosine in their diets, since they cannot synthesize tyrosine from phenylalanine, so tyrosine becomes essential in the diet of PKU patients (Lou et al., 1987).
The importance of the present results arises from the formation of most of essential amino acids (histidine, isoleucine, leucine, lysine, methionine, phenylalanine, threonine and valine) in the fermented medium, except tryptophan, which was not detected, additionally, the results showed relatively adequate content of branched chain amino acids (BCAAs); leucine, isoleucine and valine in fermented medium. BCAAs are proteinogenic amino acids and belonging to the essential amino acids for humans and account for ~35% of the essential amino acids, required to stimulate the building and reduce breakdown of muscle proteins and also account for ~40% of the preformed amino acids required by mammals. However, BCAAs that are ingested in excess are quickly disposed of, where the animal and human cells have a tightly controlled enzymatic system for BCAAs degradation. It was suggested that changes in plasma amino acids concentration may play a role in central fatigue by influencing the synthesis of neurotransmitters, particularly 5-HT in the brain. That is why some people use BCAAs to prevent fatigue and improve concentration and treatment of some diseases (Harper et al., 1984; Fernstrom, 1990; Blomstrand, 2001; Falade and Adewusi, 2013). However, free amino acids and other nitrogen-containing compounds are usually associated with enzymatic hydrolysis of protein (Ventura et al., 2012).
All the previously mentioned essential amino acids were detected in the fermented OC filtrate contained with some of non-essential amino acids. That wide variation of the amino acids makes the product rich and somehow complementary as amino acids source for nutrition. In addition, the wide collection of obtained amino acids suggesting the presence of several kinds of proteases such as serine, metallo, carboxyl, acidic, neutral and alkaline proteases, which differ in their structure or properties of the active site (Raj et al., 2012). Finally, the content of amino acids found in the filtrate was associated with high protease production, indicating that such amino acids were formed in situ degradation of medium protein.
CONCLUSION
The pronounced proteolytic activity of B. subtilis ATCC 11774 was directed to the biosynthesis of amino acids, it was found to produce appreciable amount of protease. The statistical optimization approach revealed peptone, OC and pH as the major factors affecting protease production. However, the proposed medium contained olive cake, which represents cheap source for microbial growth. This economically optimized medium could be used for the biosynthesis of both protease and amino acids, since both of them are correlated and have wide applications. Protease and amino acids from olive cake is coinciding with the attempts of many investigators to find cheap alternative substrate for the synthesis of bio-products. As far as we know the present study is the first work in this respect and introduces dual valuable end-products from single fermentation medium. However, additional work is needed to separate single amino acids, especially glutamic acid from the obtained combination of amino acids.
REFERENCES
- Alu'datt, M.H., I. Alli, K. Ereifej, M. Alhamad, A.r. Al-Tawaha and T. Rababah, 2010. Optimisation, characterisation and quantification of phenolic compounds in olive cake. Food Chem., 123: 117-122.
CrossRefDirect Link - Calik, P., E. Bilir, G. Calik and T.H. Ozdamar, 2002. Influence of pH conditions on metabolic regulations in serine alkaline protease production by Bacillus licheniformis. Enzyme Microbial. Technol., 31: 685-697.
CrossRefDirect Link - Chauhan, B. and R. Gupta, 2004. Application of statistical experimental design for optimization of alkaline protease production from Bacillus sp. RGR-14. Process Biochem., 39: 2115-2122.
CrossRefDirect Link - Dutta, J.R., P.K. Dutta and R. Banerjee, 2004. Optimization of culture parameters for extracellular protease production from a newly isolated Pseudomonas sp. using response surface and artificial neural network models. Process Biochem., 39: 2193-2198.
CrossRefDirect Link - Falade, O.S. and S.R.A. Adewusi, 2013. Comparative analysis of some essential amino acids and available lysine in Acacia colei and A. tumida seeds using chemical methods and an amino acid analyzer. J. AOAC. Int., 96: 102-109.
Direct Link - Fernstrom, J.D., 1990. Aromatic amino acids and monoamine synthesis in the central nervous system: Influence of the diet. J. Nutr. Biochem., 90: 508-517.
CrossRefDirect Link - Fontaine, J. and M. Eudaimon, 2000. Liquid chromatographic determination of lysine, methionine and threonine in pure amino acids (feed grade) and premixes: Collaborative study. J. AOAC Int., 83: 771-783.
Direct Link - Gupta, R., Q. Beg and P. Lorenz, 2002. Bacterial alkaline proteases: Molecular approaches and industrial applications. Appl. Microbiol. Biotechnol., 59: 15-32.
CrossRefPubMedDirect Link - Harper, A.E., R.H. Miller and K.P. Block, 1984. Branched-chain amino acid metabolism. Annu. Rev. Nutr., 4: 409-454.
CrossRefPubMedDirect Link - Kole, M.M., I. Draper and D.F. Gerson, 1988. Production of protease by Bacillus subtilis using simultaneous control of glucose and ammonium concentrations. J. Chem.Technol. Biotechnol., 41: 197-206.
CrossRefDirect Link - Kumar, C.G. and H. Takagi, 1999. Microbial alkaline proteases: From a bioindustrial viewpoint. Biotechnol. Adv., 17: 561-594.
CrossRefPubMedDirect Link - Lou, H.C., C. Lykkelund, A.M. Gerdes, H. Udesen and P. Bruhn, 1987. Increased vigilance and dopamine synthesis by large doses of tyrosine or phenylalanine restriction in phenylketonuria. Acta Paediatrica, 76: 560-565.
CrossRefDirect Link - Lupin, I.V., D. Korner, A. Taufel and H. Ruttloff, 1982. Application of automatic protease determination in a fermenter. Enzyme Microb. Technol., 4: 104-106.
CrossRefDirect Link - Molina-Alcaide, E. and D.R. Yanez-Ruiz, 2008. Potential use of olive by-products in ruminant feeding: A review. Anim. Feed Sci. Technol., 147: 247-264.
CrossRefDirect Link - Narayan, S.B., G. Ditewig-Meyers, K.S. Graham, R. Scott and M.J. Bennett, 2011. Measurement of plasma amino acids by Ultraperformance® liquid chromatography. Clin. Chem. Lab. Med., 49: 1177-1185.
CrossRefDirect Link - Oskouie, S.F.G., F. Tabandeh, B. Yakhchali and F. Eftekhar, 2008. Response surface optimization of medium composition for alkaline protease production by Bacillus clausii. Biochem. Eng. J., 39: 37-42.
CrossRefDirect Link - Plackett, R.L. and J.P. Burman, 1946. The design of optimum multifactorial experiments. Biometrika, 33: 305-325.
CrossRefDirect Link - Potumarthi, R., C. Subhakar and A. Jetty, 2007. Alkaline protease production by submerged fermentation in stirred tank reactor using Bacillus licheniformis NCIM-2042: Effect of aeration and agitation regimes. Biochem. Eng. J., 34: 185-192.
CrossRefDirect Link - Raj, A., N. Khess, N. Pujari, S. Bhattacharya, A. Das and S.S. Rajan, 2012. Enhancement of protease production by Pseudomonas aeruginosa isolated from dairy effluent sludge and determination of its fibrinolytic potential. Asian Pac. J. Trop. Biomed., 2: S1845-S1851.
CrossRefDirect Link - Reddy, L.V.A., Y.J. Wee, J.S. Yun and H.W. Ryu, 2008. Optimization of alkaline protease production by batch culture of Bacillus sp. RKY3 through plackett-burman and response surface methodological approaches. Bioresour. Technol., 99: 2242-2249.
CrossRefPubMedDirect Link - Rodis, P.S., V.T. Karathanos and A. Mantzavinou, 2002. Partitioning of olive oil antioxidants between oil and water phases. J. Agric. Food Chem., 50: 596-601.
CrossRefDirect Link - Suganthi, C., A. Mageswari, S. Karthikeyan, M. Anbalagan, A. Sivakumar and K.M. Gothandam, 2013. Screening and optimization of protease production from a halotolerant Bacillus licheniformis isolated from saltern sediments. J. Genet. Eng. Biotechnol., 11: 47-52.
CrossRefDirect Link - Ventura, A.K., A.S. Gabriel, M. Hirota and J.A. Mennella, 2012. Free amino acid content in infant formulas. Nutr. Food Sci., 42: 271-278.
CrossRefDirect Link - Yegin, S., M. Fernandez-Lahore, U. Guvenc and Y. Goksungur, 2010. Production of extracellular aspartic protease in submerged fermentation with Mucor mucedo DSM 809. Afr. J. Biotechnol., 9: 6380-6386.
Direct Link