
Mehpara Maqsood
Cellular Differentiation and Molecular Genetics Section, Department of Botany, Hamdard University, 110062, New Delhi, India
A. Mujib
Cellular Differentiation and Molecular Genetics Section, Department of Botany, Hamdard University, 110062, New Delhi, India
Zahid H. Siddiqui
Cellular Differentiation and Molecular Genetics Section, Department of Botany, Hamdard University, 110062, New Delhi, India







ABSTRACT
An encapsulation protocol to obtain synthetic seeds was established in Catharanthus roseus (L.) G. Don. Hypocotyl derived embryogenic callus containing somatic embryos viz., torpedo, early cotyledonary and cotyledonary were encapsulated in sodium alginate and calcium chloride solution. Sodium alginate acts as an artificial endosperm, providing nourishment to the growing embryos, at the same time it protects embryos from damages and facilitates embryo’s germination later. Different levels of sodium alginate and calcium chloride were used in which perfect bead formation was observed in condition encapsulated with 2.5% sodium alginate and 100 mM calcium chloride solution. Calcium chloride exposure for 15 min improved bead quality by forming uniform and firm beads. Addition of 3% sucrose to sodium alginate solution was also very effective for synthetic seed germination. The encapsulated embryos germinated and produced seedlings at a high rate (84.33±2.1) in 1.34 μM α-Naphthalene acetic acid +1.10 μM 6-Benzylaminopurine amended MS medium. Synthetic seeds were kept at 0, 4 and 25°C as to examine the best storage temperature; preservation at 4°C was found to be the optimum temperature for embryo storage and germination purposes. The encapsulated embryos were preserved up to 10 weeks or more without losing germination abilities. The germination frequency was high (81.0%) after 30 days of storage which however reduced with extended storage time. Plantlets obtained from synseeds were morphologically similar to that of mother plant.
PDF Abstract XML References Citation
Received: October 05, 2011;
Accepted: December 17, 2011;
Published: February 15, 2012
How to cite this article
Mehpara Maqsood, A. Mujib and Zahid H. Siddiqui, 2012. Synthetic Seed Development and Conversion to Plantlet in Catharanthus roseus (L.) G. Don. Biotechnology, 11: 37-43.
DOI: 10.3923/biotech.2012.37.43
URL: https://scialert.net/abstract/?doi=biotech.2012.37.43
DOI: 10.3923/biotech.2012.37.43
URL: https://scialert.net/abstract/?doi=biotech.2012.37.43
INTRODUCTION
Catharanthus roseus (L.) G. Don of the family Apocyanaceae is one of the most widely investigated medicinal plants. Medicinal plants are the traditional source of drugs (Jha et al., 2011). The plant has been a fundamental source of a wide variety of alkaloids of economic value for pharmacological enterprises (Loyola-Vargas et al., 2007). C. roseus is immensely valuable as it produces over 130 alkaloids, several of which are anticancerous in nature (Moreno et al., 1995). Vinblastine and vincristine are perhaps the most important alkaloids because of their anti-cancerous property. Under normal circumstances, the yield of these two compounds is low. In plant biotechnology, different tissues have been used to establish culture and the alkaloid content has been analyzed in C. roseus (Whitmer et al., 1998; Batra et al., 2004). Although, the technique of somatic embryogenesis has been reported in a wide range of plants (Thorpe, 1995; Mujib and Samaj, 2006) the embryogenic tissue has never been utilized in alkaloid enrichment programme in C. roseus. Recently, however, Aslam et al. (2009) and Junaid et al. (2010) screened and isolated a few cell/tissue lines that overproduced alkaloids. Maintaining the superior lines of embryogenic cultures for longer period and overproducing some of the valuable alkaloid compounds, would thus be highly desirable. Fatima et al. (2009) established a vitrification-based cryopreservation protocol in C. roseus wherein embryogenic cultures were preserved in vitro by using preculture, pretreatment and liquid nitrogen medium. The same embryogenic culture may be used to produce synthetically coated seeds to conserve superior cell lines on a large scale as was reported earlier in other groups of plants (Redenbaugh et al., 1993). Normally plants propagate by seeds but it is a slow process; seeds and seedlings become harmed due to unfavourable conditions of nature (Salah Uddin et al., 2005). Propagation and conservation of medicinal plants is one of the major concerns, thus immediate efforts are required in this regard (Quraishi et al., 2011). Micropropagation has been extensively used for rapid multiplication of many plant species, however, its wider use is restricted often by high percentage of plant loss when transferred to ex vitro environment (Deb and Imchen, 2010). Synthetic seeds are defined as tissues (somatic embryos, shoot buds, cell aggregates or other organs) that are artificially encapsulated and are used as a seed that possess the ability to regenerate into a plant under in vitro or ex vitro conditions even after storage (Capuano et al., 1998). However, the production of synthetic seeds requires efficient encapsulation of somatic embryos. Choice of the tissue to be employed also needs fundamental deliberation. There have been many studies on encapsulation of different plant materials; somatic embryos were also tried recently in several plant systems like in sugarcane (Nieves et al., 2003), Paulownia elongate (Ipekci and Gozukimizi, 2003) and Daucus carota (Latif et al., 2007). However, less attention has unfortunately been paid to encapsulation of non-embryogenic vegetative propagules such as nodal segments, axillary buds, shoot tips, hairy roots or callus (Zych et al., 2005).
The present study has been undertaken to develop an efficient encapsulation method for plantlet regeneration of C. roseus from the synthetic seeds produced from hypocotyls derived somatic embryos. The effect of encapsulation matrix, mineral nutrients, plant growth regulators and temperature on the conversion ability of synthetic seeds was also monitored.
MATERIALS AND METHODS
Plant material: Plants of Catharanthus roseus (L.) G. Don. cv. Nirmal were collected from Jamia Hamdard (Hamdard University campus, New Delhi) herbal garden and was identified (by Dr. M.P. Sharma, Plant Taxonomist, Department of Botany, Hamdard University). The voucher specimen was deposited in the Herbarium of the same department. For initiating fresh culture 20-25 seeds from these plants were surface sterilized by immersion in 70% (v/v) ethanol for 30 sec and in 0.5% sodium hypochlorite with a drop of triton-X for 5 min followed by three rinses with double distilled water for over 5 min each.
Somatic embryogenesis: Seeds were subsequently placed in Magenta-7 vessels containing 50 mL of MS medium (Murashige and Skoog, 1962) without any growth regulators. Seeds were germinated into seedlings and various parts (nodal stem, leaf, root and hypocotyls) were used as explants. All the explants were incubated on a basal MS medium (pH = 5.7) and grown under 16 h photoperiod provided by 40 W cool-white fluorescent tubes (100 μmol-2 sec-1). Hypocotyls of seedlings induced embryogenic callus when supplied with 4.52 μM 2, 4-D while the other plant parts produced non-embryogenic callus. The embryogenic callus differentiated into all four types of somatic embryos in due course of time. In the present study, mature cotyledonary somatic embryos were used for encapsulation work.
Encapsulation: Somatic embryos of C. roseus were encapsulated in different concentrations of sodium alginate (CDH, New Delhi, India; 2, 2.5, 3, 3.5 and 4%) and CaCl2.2H2O (75, 100 and 125 mM) solution (Merck, Mumbai, India). Alginate solution was prepared by adding 3% sucrose and different concentrations of sodium alginate (w/v) in double distilled H2O and later sterilized at 121°C. Embryos were mixed with sodium alginate solution for a few seconds, picked up by pipette and placed in sterile aqueous solution of calcium chloride (CaCl2.2H2O) for varying time (5, 10, 15 and 20 min) for hardening. With occasional agitation on a rotary shaker, it resulted in bead formation. Beads were taken out by decanting off the CaCl2 solution and washed with sterilized water. Freshly prepared beads were transferred to MS medium fortified with different concentrations of Plant Growth Regulators (PGRs). The whole procedure was carried out under strict aseptic conditions.
Statistical analysis: The data on the effects of PGRs on synthetic seeds conversion into plantlet with and without storage were analyzed by one-way analysis of variance (ANOVAs) followed by Least Significant Difference (LSD). Values are means of five replicates from two experiments with six to nine embryos in each replicate. The presented mean values were separated using LSD at p≤0.05.
RESULTS
Effect of sodium alginate percentage on synthetic seed formation and conversion rate: Somatic embryos (Fig. 1a) kept in 2.5% sodium alginate and 100 mM calcium chloride produced synthetic seeds (Fig. 1b) with firm coat, suitable for handling which also favoured easy development of plantlets. In contrast, at low concentration of sodium alginate, uniform and sufficiently firm beads were not formed, resulting in a reduced frequency of conversion. Maximum conversion frequency (83.33±2.48%) was noted in embryos encapsulated with 2.5% alginate (Table 1).
| Table 1: | The effect of different concentrations of sodium alginate on conversion frequency (%) of synthetic seeds in C. roseus. The MS medium contained 2.20 μM BAP |
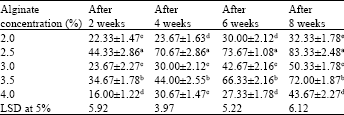 | |
| Values are Mean±SE of at least 3 replicates with 6 embryos in each replicate. Within each column, values are followed by the letters are not significantly different at p≤0.05 level according to LSD test | |
 | |
| Fig. 1a-f: | Encapsulation of somatic embryos and plant regeneration in C. roseus. (a) Somatic embryos (arrows indicating cotyledonary embryos) in C. roseus, (b) Bead formation, (c) Beads cultured on MS medium for germination, (d, e) Germination of synseeds, arrows indicating alginate shell and (f) Whole plantlet regenerated from synthetic seed. (1-a and 1-d Bar = 2 mm, 1-b and 1-c Bar = 1 mm, 1-e Bar = 3 mm and 1-f bar = 0.5 cm) |
At 2% sodium alginate conversion frequency was low. Miniature plantlets emerged out by bursting of alginate capsule layers by the growing embryos (Fig. 1c, e).
Effect of molar strength of calcium chloride and its exposure time on conversion rate: Encapsulated embryos were also exposed to different levels of calcium chloride in order to optimize the right concentration. Calcium chloride at 100 mM proved effective for spherical bead formation, it also hastened conversion frequency markedly (81±1.56%). In 125 mM calcium chloride, least conversion (55±1.25%) was recorded (Fig. 2). We also observed that the embryos kept in calcium chloride for 15 min produced firm, round and uniform beads. Embryo exposure below and above 15 min induced soft and harder beads respectively with poor conversion ability. In all cases, conversion started to occur within 10-15 days of incubation. The development of root and shoot was simultaneous and showed rapid growth of plantlet within 6-7 weeks time. Some of the synthetic seeds failed to convert into plantlet and turned brown within the alginate matrix after 10-15 days of culture.
Effect of plant growth regulators on conversion rate: The synthetic beads were cultured directly on MS medium amended with PGRs at various concentrations 0.64, 1.34 and 2.69 μM NAA; 0.53, 1.10 and 2.20 μM BAP; 0.35, 0.72 and 1.44 μM GA3.
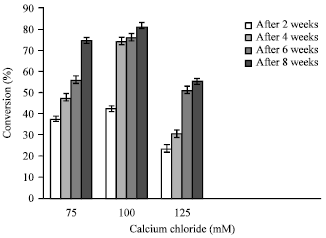 | |
| Fig. 2: | The effect of different calcium chloride concentrations (mM) on the conversion frequency of synthetic seeds (2.5% sodium alginate) |
Other PGR combinations, i.e., 0.64 μM NAA+0.53 μM BAP, 1.34 μM NAA+ 1.10 μM BAP and 2.69 μM NAA+2.20 μM BAP were also used to test the conversion efficiency of encapsulated embryos. Plantlet conversion from synthetic seeds was low (13.33-35.33%) on medium amended singly with NAA, BAP and GA3 and with increase in time, conversion frequency improved up to 75.33%.
| Table 2: | Effect of different concentrations and combinations of PGRs on conversion frequency (%) of synthetic seeds (2.5% sodium alginate, 100 mM calcium chloride and 15 min exposure time) on MS medium |
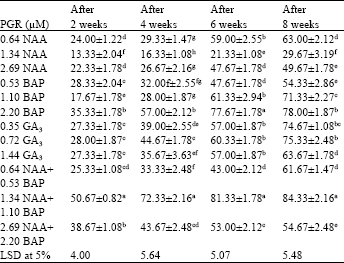 | |
| Values are Means±SE of at least 5 replicates with 8 embryos in each replicate. Within each column, values are followed by the letters are not significantly different at p≤0.05 level according to LSD test | |
MS medium amended with a combination of 1.34 μM NAA and 1.10 μM BAP showed maximum conversion (84.33±2.16%) of synthetic seeds while it was very low in 1.34 μM NAA (29.67±3.19%) added medium (Table 2).
Different storage temperatures and conversion rate of synthetic seeds: Encapsulated embryos stored at 4°C showed maximum conversion frequency than the embryos stored at 0 and 25°C (Table 3). At 4°C, 57.33±1.78% synthetic seeds were converted into plantlets in about 4 weeks time that improved to 81.67±3.89% after 8 weeks of incubation. Embryos that were stored at 25°C dried out more rapidly and showed low rate of conversion. Our studies also showed that synthetic seeds stored at 0°C quite successfully preserved the encased embryo and at this low temperature conversion ability was also maintained.
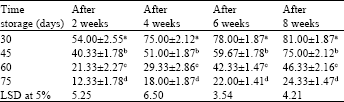
Effect of storage time (duration) on conversion of synthetic seeds: Table 4 compares the conversion frequencies of synthetic seeds stored at 4°C for various time periods. Synthetic seeds transferred to medium after 30 days of incubation showed maximum conversion frequency (81.00±1.8%) which however, reduced to 46.33±2.16% after 60 days of incubation. The conversion frequency was further lowered (24.33±1.47%) when the encapsulated embryos were kept for longer (75 days) period of time. Although, the conversion percentages gradually reduced with time, C. roseus synthetic seeds can be preserved for 10 weeks or more under in vitro conditions utilizing low temperature.
| Table 3: | Conversion frequency of synthetic seeds (2.5% sodium alginate and 100 mM calcium chloride and 15 min exposure time) in C. roseus after storage at different temperatures. MS medium fortified with optimized 1.34 μM NAA+1.10 μM BAP |
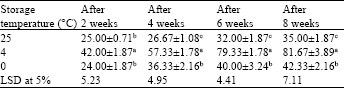 | |
| Values are Means±SE of at least 5 replicates with 6 embryos in each replicate. Within each column, values are followed by the letters are not significantly different at p≤0.05 level according to LSD test | |
| Table 4: | Effect of various storage time on conversion frequencies of synthetic seeds (2.5% sodium alginate, 100 mM calcium chloride and 15 min exposure time) at 4°C. MS medium contained optimized 1.34 μM NAA+1.10 μM BAP |
 | |
| Values are Means±SE of at least 5 replicates with 9 embryos in each replicate. Within each column, values are followed by the letters are not significantly different at p≤0.05 level according to LSD test | |
DISCUSSION
Somatic embryogenesis acts as an alternative to organogenesis for regeneration of plants. In this work we have used sodium alginate and calcium chloride for encapsulation of embryos that would subsequently be preserved at low temperatures and their viability was assessed later on. The method employs a highly asceptic procedure which shall in turn ensure the protection from eventual contamination of the culture. Different combinations of sodium alginate and calcium chloride have been employed to efficiently optimize the conversion ability of the encapsulated embryos. Maximum encapsulation was seen by treating the embryos with 2.5% sodium alginate and 100 mM calcium chloride. In plant systems like mulberry (Patnaik et al., 1995) and Hyoscyamus muticus L. (Pandey and Chand, 2005), 3.0% sodium alginate along with 70-80 mM calcium chloride were used for encapsulation and showed maximum conversion capacity. A 2 and 4% solution of sodium alginate upon complexation with 100 mM calcium chloride produced clear transparent firm capsules with an ion exchange duration of 30 min in Flickingeria nodosa (Dalz) Seidenf (Nagananda et al., 2011). It has been shown that the percentage of sodium alginate employed for maximum conversion ability depends on the species of the plant under investigation (Redenbaugh et al., 1986). We have observed that a low percentage of sodium alginate resulted in the formation of tender and weak beads that have comparatively low conversion capacity. The presence of high percentage of sodium alginate resulted in the formation of hard beads that had a low conversion frequency in conformity with observation of Mohanraj et al. (2009). Several authors observed similar results in other investigated species like Papaya (Castillo et al., 1998) in which uniform beads were formed on 2.5% sodium alginate added medium.
The concentration of the complexing agent calcium chloride has also shown a significant impact on conversion capacity in this present case. A 15 min exposure to calcium chloride induced firm, round and uniform beads. It is known that the capsule hardiness depends upon optimal ion exchange of Na+ and Ca2+, however, this may vary with different propagules as well as with different plant species (Rai et al., 2009). It was also observed that the nutrient supplemented to the alginate matrix reduced the viscosity and the ability of the gel to form solid beads (Daud et al., 2008). We noticed that polymer concentration, alginate viscosity, calcium concentration and exposure time are fundamental in determining a successful encapsulation technique.
The present study also revealed the influence of plant growth regulators on the conversion of embryos. We noticed highest conversion rate when the matrix was supplied with 1.34 μM NAA+1.10 μM BAP. Combination of NAA and BAP also showed increase in number of regenerated shoots in Taxus baccata (Abbasin et al., 2010), tea roses (Noriega and Sondahl, 1991) but in some cases resulted in increased callus induction for example in tomato (Harish et al., 2010). Contrary to this, sometimes BAP alone proved best for the shoot induction as in Hibiscus sabdariffa L. (Gomez-Leyva et al., 2008). It has been reported that the encapsulated somatic embryos may survive for a period of at least 4 weeks when stored at 4°C and no loss in conversion capacity was observed (Lulsdorf et al., 1993). Similar results were also reported in Olive (Micheli et al., 1998) and in apple rootstock (Sicurani et al., 2001). In our study, synthetic seeds stored at 4°C showed a higher resistance to storage than the naked embryos and showed maximum conversion capacity. An increase in temperature resulted in desiccation of embryos and loss in conversion rate. The release from cold-stratification of normal seeds also has been demonstrated in different Bunium persicum populations (Sharifi and Pouresmael, 2006). Encapsulated somatic embryos of Olive stored at 2 or 4°C for 2-3 months showed high conversion rates of 61% (Micheli et al., 1998). It has been suggested that several compounds such as amino acids, ammonium compounds, sugars, sugar alcohols are possibly produced by cultivated plant cells in response to stress (osmotic, salt, low temperature, desiccation etc.) provide protection to cells (Hincha et al., 2006). It is, however, very difficult to explain the mechanism by which a compound or group of compounds acting together while giving tolerance to cells. In this present investigation, it appeared that the added alginate and sugar in capsule might be involved as functional compound in making cells tolerant to freezing. Nieves et al. (2001) observed that hydrated synthetic seeds were difficult to store at room temperature because of depleted nutritive reservoir and lack of quiescence that lowered conversion percentage. In alfalfa, at 4°C even, the storage life of synthetic seeds was quite short (Redenbaugh et al., 1987) as the conversion rates were poor in Asparagus cooperi (Ghosh and Sen, 1994) and in Ecalyptus citrisdora (Muralitharan and Mascarenhas, 1995). However, in Santalum album, the encapsulated embryos retained their conversion (18%) ability for over 6 weeks or more after storage at 14°C (Rao et al., 1993). In the present study, synthetic seeds showed highest frequency of conversion after 30 days of storage which was, however, reduced when encapsulated embryos were kept for extended period of time indicating that with increased storage periods conversion frequency is reduced. In normal seeds also, the conversion frequency decreases with increase in time as found in Bunium persicum (Sharma and Sharma, 2010). The decline in conversion from encapsulated embryo into plants may be related to both oxygen deficiency in the gel bead and its rapid drying (Swamy et al., 2009). The exploitation of stable and regenerative embryogenic masses is an attractive tool for the fast production of clonal plants including transgenics where synthetic seed technology can preserve those mother tissues. In C. roseus, encapsulation technology is therefore important to conserve important cell lines in which one plantlet may be produced from one bead after short to medium term storage (60-75 days).
CONCLUSION
Present study reports the feasibility of using somatic embryos for germplasm conservation. The study further envisages the optimization of encapsulation matrix for large scale preparation of synthetic seeds. The encapsulated somatic embryos can be stored at low temperature which helps to preserve elite germplasm for medium to long term basis. On requirement, plantlets can be recovered directly without any induction of callus phases, avoiding somaclonal variation among regenerated population.
REFERENCES
- Abbasin, Z., S. Zamani, S. Movahedi, G. Khaksar and B.E.S. Tabatabaei, 2010. In vitro micropropagation of yew (Taxus Baccata) and production of plantlets. Biotechnology, 9: 48-54.
CrossRefDirect Link - Batra, J., A. Datta, D. Singh, S. Kumar and J. Sen, 2004. Growth and terpenoids indole alkaloid production in Catharanthus roseus hairy root clones in relation to left- and right-termini-linked Ri T-DNA gene integration. Plant Cell Rep., 23: 148-154.
CrossRefDirect Link - Castillo, B., M.A.L. Smith and U.L. Yadava, 1998. Plant regeneration from encapsulated somatic embryos of Carica papaya L. Plant Cell Rep., 17: 172-176.
CrossRefDirect Link - Daud, N., R.M. Taha and N.A. Hasbullah, 2008. Artificial seed production from encapsulated micro shoots of Saintpaulia ionantha Wendl. (African Violet). J. Applied Sci., 8: 4662-4667.
CrossRefDirect Link - Deb, C.R. and T. Imchen, 2010. An efficient in vitro hardening technique of tissue culture raised plants. Biotechnology, 9: 79-83.
CrossRefDirect Link - Fatima, S., A. Mujib, S.A. Nasim and Z.H. Siddiqui, 2009. Cryopreservation of embryogenic cell suspensions of Catharanthus roseus L. (G) Don. Plant Cell Tiss. Org. Cult., 98: 1-9.
CrossRefDirect Link - Ghosh, B. and S. Sen, 1994. Plant regeneration from alginate encapsulated somatic embryos of Asparagus cooperi baker. Plant Cell Rep., 13: 381-385.
CrossRefDirect Link - Harish, M.C., S. Rajeevkumar and R. Sathishkumar, 2010. Efficient in vitro callus induction and regeneration of different tomato cultivars of India. Asian J. Biotechnol., 2: 178-184.
CrossRefDirect Link - Hincha, D.K., E. Zuther, M. Hundertmark and A.G. Heyer, 2006. The Role of Compatible Solutes in Plant Freezing Tolerance: A Case Study on Raffinose. In: Cold Hardiness in Plants: Molecular Genetics, Cell Biology and Physiology, Chen, T.H.H., M. Uemura and S. Fujikawa (Eds.). CAB International, London, UK., pp: 203-218.
Direct Link - Ipekci, Z. and N. Gozukimizi, 2003. Direct somatic embryogenesis and synthetic seed production from Paulownia elongate. Plant Cell Rep., 22: 16-24.
CrossRefDirect Link - Jha, T.B., A. Dafadar and R.K. Chaudhuri, 2011. Somatic embryogenesis in Swertia chirata buch. ham. ex wall.-A multipotent medicinal herb. Asian J. Biotechnol., 3: 186-193.
CrossRefDirect Link - Aslam, J., A. Mujib, S.A. Nasim and M.P. Sharma, 2009. Screening of vincristine yield in ex vitro and in vitro somatic embryos derived plantlets of Catharanthus roseus L. (G) Don. Sci. Hort., 119: 325-329.
CrossRefDirect Link - Aslam, J., A. Mujib, Z. Fatima and M.P. Sharma, 2010. Variations in vinblastine production at different stages of somatic embryogenesis, embryo and field-grown plantlets of Catharanthus roseus L. (G) Don, as revealed by HPLC. In vitro Cell. Dev. Biol.-Plant, 46: 348-353.
CrossRefDirect Link - Latif, Z., A.N. Idrees and S. Riazuddin, 2007. Indigenous production of synthetic seeds in Daucus carota. Pak. J. Bot., 39: 849-855.
Direct Link - Loyola-Vargas, V.M., R.M. Galaz-Avalos and R. Ku-Cauich, 2007. Catharanthus biosynthetic enzymes: The road ahead. Phytochem. Rev., 6: 307-339.
CrossRef - Mohanraj, R., R. Ananthan and V.N. Bai, 2009. Production and storage of synthetic seeds in Coelogyne breviscapa lindl. Asian J. Biotechnol., 1: 124-128.
CrossRefDirect Link - Moreno, P.R.H., R. van der Heijden and R. Verpoorte, 1995. Cell and tissue cultures of Catharanthus roseus: A literature survey. II: Updating from 1988 to 1993. Plant Cell Tissue Org. Cult., 42: 1-25.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Nagananda, G.S., N. Satishchandra and S. Rajath, 2011. Regenetion of encapsulated protocorm like bodies of medicinally important vulnerable orchid Flickingeria nodosa (Dalz.) Seidenf. Int. J. Bot., 7: 310-313.
CrossRefDirect Link - Nieves, N., M.E. Martinez, R. Castillo, M.A. Blanco and J.L. Gonzalez-Olmedo, 2001. Effect of abscisic acid and jasmonic acid on partial desiccation of encapsulated somatic embryos of sugarcane. Plant Cell Tissue Organ Cult., 65: 15-21.
CrossRef - Nieves, N., Y. Zambrano, R. Tapia, M. Cid, D. Pina and R. Castillo, 2003. Field performance of artificial seedderived sugarcane plants. Plant Cell Tiss. Organ Cult., 75: 279-282.
CrossRefDirect Link - Pandey, A. and S. Chand, 2005. Efficient plant regeneration from encapsulated somatic embryos of Hyoscyamus muticus L. Ind. J. Biotechnol., 4: 546-550.
Direct Link - Patnaik, S.K., Y. Sahoo and P.K. Chand, 1995. Efficient plant retrieval from alginate-encapsulated vegetative buds of mature mulberry trees. Sci. Hortic., 61: 227-239.
CrossRefDirect Link - Quraishi, A., S.K. Jadhav and S. Gupta, 2011. In vitro clonal propagation of Cassia tora L. (Coffee Pod): A medicinal plant. Biotechnology, 1062: 546-550.
CrossRefDirect Link - Rai, M.K., P. Aathana, S.K. Singh, V.S. Jaiswal and U. Jaiswal, 2009. The encapsulation technology in fruit plants-a review. Biotechnol. Adv., 27: 671-679.
CrossRefDirect Link - Redenbaugh, K., B.D. Paasch, J.W. Nichol, M.E. Kossler and P.R. Viss and K.A. Walker, 1986. Somatic seeds encapsulation of asexual plant embryos. Biotechnology, 4: 797-801.
CrossRefDirect Link - Salah Uddin, M., K. Nasirujjaman, S. Zaman and M.A. Reza, 2005. Regeneration of multiple shoots from different explants viz. shoot tip, nodal segment and cotyledonary node of in vitro grown seedlings of Peltophorum pterocarpum (DC.) backer EX K. Heyne. Biotechnology, 4: 35-38.
CrossRefDirect Link - Sharifi, M. and M. Pouresmael, 2006. Breaking seed dormancy in Bunium persicum by stratification and chemical substances. Asian J. Plant Sci., 5: 695-699.
CrossRefDirect Link - Sharma, R.K. and S. Sharma, 2010. Effect of storage and cold-stratification on seed physiological aspects of Bunium persicum: A threatened medicinal herb of trans-Himalaya. I. J. Bot., 6: 151-156.
CrossRefDirect Link - Sicurani, M., E. Piccioni and A. Standardi, 2001. Micropropagation and preparation of synthetic seed in m.26 apple rootstock i: Attempts towards saving labor in the production of adventitious shoot tips suitable for encapsulation. Cultures, 66: 207-216.
CrossRef - Swamy, M.K., S. Balasubramanya and M. Anuradha, 2009. Germplasm conservation of patchouli (Pogostemon cablin Benth.) by encapsulation of in vitro derived nodal segments. Int. J. Biodivers. Conserv., 104: 224-230.
Direct Link - Whitmer, S., C. Canel, D. Hallard, C. Goncalves and R. Verpoorte, 1998. Influence of precursor availability on alkaloid accumulation by transgenic cell line of Catharanthus roseus. Plant Physiol., 116: 853-857.
Direct Link - Zych, M., M. Furmanowa, A. Krajewska-Patan, A. Lowicka, M. Dreger, and S. Mendlewska, 2005. Micropropagation of Rhodiola kirilowii plants using encapsulated axillary buds and callus. Acta Biol. Cracoviensia Ser. Bot., 47: 83-87.
Direct Link