
H.S. Dalla-Santa
Bioprocess Engineering and Biotechnology Division, Federal University of Parana, Curitiba, Parana, Brazil
R. Rubel
Bioprocess Engineering and Biotechnology Division, Federal University of Parana, Curitiba, Parana, Brazil
F.M.D. Vitola
Bioprocess Engineering and Biotechnology Division, Federal University of Parana, Curitiba, Parana, Brazil
J.A. Rodriguez-Leon
Departament of Bioingenieria, Ingenieria Quimica, 1968, Universidad de La Habana, Cuba
O.R. Dalla-Santa
Laboratory of Microbiology, Department of Food Engineering, Midwest State University, Guarapuava, Parana, Brazil
D. Brand
Bioprocess Engineering and Biotechnology Division, Federal University of Parana, Curitiba, Parana, Brazil
D.C. Alvarez
Laboratory of Microbiology, Department of Food Engineering, Midwest State University, Guarapuava, Parana, Brazil
R.E.F. Macedo
Laboratory of Agricultural and Livestock Technology, Pontifical Catholic University of Parana, S�o Jose dos Pinhais, Parana, Brazil
J.C. Carvalho
Bioprocess Engineering and Biotechnology Division, Federal University of Parana, Curitiba, Parana, Brazil
C.R. Soccol
Bioprocess Engineering and Biotechnology Division, Federal University of Parana, Curitiba, Parana, Brazil
Biotechnology
Year: 2012 | Volume: 11 | Issue: 3 | Page No.: 144-153







ABSTRACT
Agaricus brasiliensis mycelium is rich in antitumoral molecules such proteoglycans and ergosterol and this is the first study to report production of mycelium and ergosterol by A. brasiliensis in solid-state fermentation. The aim of this research was to study the biomass content and several growth parameters of A. brasiliensis mycelium on grounded wheat grain as the substrate in solid-state fermentation with ergosterol as the main growth indicator. The growth parameters were determined by the respirometric activity in the columns of a bioreactor with forced aeration and the outlet air composition was determined by gas chromatography. Ergosterol was extracted, measured by liquid chromatography and used for calculating biomass production. Then, the respirometric activity and ergosterol production data were used to estimate biomass production with the program Fersol (F). The growth parameters resulted in an achieved specific growth velocity of 0.016 h-1 at 18 h and biomass yield (YX/O) = 0.160 g of biomass g-1 of consumed O2 at 302 h of fermentation. The analysis resulted in ergosterol values of 6.71 mg g-1 of fungi biomass (Dry Weight: DW) from submerged fermentation and 1.95 mg g-1 of solid-state-fermented wheat grains. The correlation with biomass production allowed us to estimate a value of 0.29 g g-1 of fungi biomass per gram of the solid-state-fermented wheat grains. The importance of this study is allow calculation of the fungi biomass percentage in solid-state-fermented wheat grains and determination of the growth parameters of the main mycelium A. brasiliensis in this substrate, which can be milled to obtain a bio-flour to produce nutraceutical foods with beneficial effects due the presence of ergosterol which has recognized antitumor activity.
PDF Abstract XML References Citation
Received: February 06, 2012;
Accepted: May 24, 2012;
Published: July 03, 2012
How to cite this article
H.S. Dalla-Santa, R. Rubel, F.M.D. Vitola, J.A. Rodriguez-Leon, O.R. Dalla-Santa, D. Brand, D.C. Alvarez, R.E.F. Macedo, J.C. Carvalho and C.R. Soccol, 2012. Growth Parameters of Agaricus brasiliensis Mycelium on Wheat Grains in Solid-state Fermentation. Biotechnology, 11: 144-153.
DOI: 10.3923/biotech.2012.144.153
URL: https://scialert.net/abstract/?doi=biotech.2012.144.153
DOI: 10.3923/biotech.2012.144.153
URL: https://scialert.net/abstract/?doi=biotech.2012.144.153
INTRODUCTION
A. brasiliensis is a medicinal and edible mushroom with strong antitumor activity and immunostimulant properties (Stamets, 2000; Wasser, 2011). Pharmaceutical industries are interested in A. brasiliensis for the development of drugs and nutraceuticals because of these pharmacological activities (Okwulehie et al., 2007). This fungus produces several important bioactive molecules such as proteoglycans, lectins and ergosterol which are found in the fruiting body, mycelium and diffuse through the fermentation medium (Hamedi et al., 2007; Fan et al., 2007). Polysaccharide and ergosterol are the most investigated bioactive molecules produced by A. brasiliensis because of their health benefits (Lima et al., 2008). Recent research has shown that ergosterol from A. brasiliensis exhibits antitumor activity such as inhibition of angiogenesis and reduction of tumor growth (Takaku et al., 2001). This mushroom which is also known as Himematsutake, Sun-mushroom, A. blazei, A. subrufescens, or A. brasiliensis and refer to refer as A. brasiliensis in the present work, based on recent studies that clarify the problems of nomenclature of this specie (Wasser, 2011).
Ergosterol is a sterol who does not occur in plant or animal cells, however is abundant in yeast and specially in the membrane of fungi (Weete, 1973). Ergosterol plays an important role in the human body as a precursor of vitamin D which is formed in response to the exposure of ultraviolet radiation (sunlight) to the sterols present in the skin. Vitamin D enhances calcium absorption in animals for the formation of strong bones and teeth, prevention of rickets and osteoporosis and maintenance of immune activity (Stamets, 2005). There is an increasing worldwide demand for ergosterol for the production of pharmaceutical-grade, food-grade and beverage-grade vitamin D2, than the levels of vitamin d are low in a lot of foods (Pal et al., 2011). Because of this demand, Submerged Fermentation (SF) was recently assessed for optimizing ergosterol and or polysaccharide production by A. brasiliensis (Fan et al, 2003; Zou, 2005; Gao and Gu, 2007; Hamedi et al., 2007). SF is reportedly the best process for obtaining fungal mycelium and the desired biomolecules because of the shorter fermentation time, lesser space requirement and better controlled fermentation parameters (Karmakar and Ray, 2010; Fan et al., 2007; Gao and Gu, 2007).
Solid-State Fermentation (SSF) requires low capital investment because of low energy and waste output (Onyango et al., 2011; Jamal et al., 2012) and is, therefore, advantageous over SF. Furthermore, sometimes, the produced metabolites by SSF are more thermostable than those produced by SF, such α-amylase (Regulapati et al., 2007). Other advantages are the economy and reduction in contamination problems due low-water indices used in the substrate which can be used for nutrition and support for the microorganism growth (Soccol and Vandenberghe, 2003; Rubel et al., 2008). Furthermore, this method can be used to produce functional foods such as cereals when the substrates are nutritionally valuable and free of toxic components (Rubel et al., 2010). The use of A. brasiliensis mycelium produced by SF are recently used for formulation of nutraceutical foods (Ribeiro and Salvadori, 2003; Camelini et al., 2005). On the other hand, recently studies verified the beneficial biological effects of SSF wheat grain ingestion with the mushroom mycelium in mice (Dalla-Santa et al., 2009, 2010).
However, a correlation factor to determine the amount of biomass in the wheat grains or ergosterol and mycelium production by A. brasiliensis during SSF has not been described before. Therefore, the aim of the present research was to study the biomass content and several growth parameters of A. brasiliensis mycelium grown during SSF on grounded wheat grains with ergosterol as the main growth indicator.
MATERIALS AND METHODS
Microorganism and inoculum preparation: The strain used in this work was A. brasiliensis LPB-03 which is preserved in the Biotechnological Processes Laboratory of the Biotechnology and Bioprocess Engineering Division (Federal University of Parana) (Dalla-Santa et al., 2009).
The mushroom strain was grown in Petri dishes on potato dextrose agar for 10 days at 30°C. Following cultivation, 5 pieces of mycelium (1 cm2) were cut and inoculated into 50 mL of fermentation medium containing (in g L-1) glucose (20), yeast extract (3.95), MgSO4.7H2O (0.3) and K2HPO4.3H2O (0.5). The pH was adjusted to 6.0±0.2 (Fan et al., 2003) and the medium was sterilized at 121°C for 15 min. After inoculation of the 5 pieces of mycelium, incubation under shaker conditions was carried out at 30°C and 120 rpm for 7 days. The pre-inoculum was obtained by straining the mycelium produced in this first step through a 0.5 mm nylon mesh with a spatula to break pellets and produce small mycelium pieces. A suspension of broken mycelium was obtained by washing the biomass with 50 mL of distilled sterilized water during filtration. This suspension was used for the second step of the inoculum preparation, in which 50 mL of the suspension was used to inoculate 500 mL (10%, v/v) of fermentation medium containing (in g L-1) glucose (35), yeast extract (2.5), peptone (5), KH2PO4 (0.88) and MgSO4.7H2O (0.5). The pH was adjusted to 5.5 (Tang and Zhong, 2002) and incubation was performed under conditions similar to those described for the pre-inoculum preparation. The cultivated medium was filtered with 500 mL of distilled sterilized water and the obtained mycelium suspension was used as the inoculum for SF and SSF.
In order to estimate the biomass production of A. brasiliensis in SSF, SF was carried out using the same inoculum. The data obtained for ergosterol production in SF were used to determine the correlation factor between biomass and ergosterol production.
Biomass production in SF: Biomass production of A. brasiliensis in SF was carried out in Erlenmeyer flasks using 5% (v/v) of the inoculum prepared as described above. After 7 days of incubation, the biomass was filtered, washed with distilled water and vacuum-dried (10 mmHg, 45°C) for 24 h. About 0.5 g of the dried biomass was collected for analysis of moisture and ergosterol content. All analyses were performed in triplicate.
Ergosterol extraction and analysis from the SF biomass: Ergosterol extraction was performed according to the method described by De Carvalho et al. (2006) with some modifications. Initially, 0.5 g of the biomass produced by SF was weighed, transferred to a glass flask and 2 mL of analytical-grade ethanol and 1 mL of NaOH (2 M) were added.
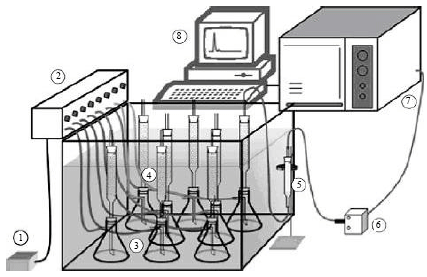 | |
| Fig. 1: | Column setup for respirometric studies, 1: Air pump, 2: Air distribution system, 3: Humidifiers 4: Fermentation columns which were immersed in a water bath with controlled temperature, 5: Drying column attached to a column exit, 6: Sampling valve, 7: Gas chromatograph and 8: Computer with data acquisition and control software |
The material was shaken and incubated for 1 h at 70°C with periodic agitation followed by the addition of 2 mL of HCl (1 M), 1 mL of KHCO3 (1 M) and 2 mL of n-hexane with subsequent stirring. The mixture was agitated, transferred to glass tubes and centrifuged at 3,000 g for 10 min at 20°C to facilitate the separation of light and heavy phases. The light phase (n-hexane) was separated and 2 more extractions with 2 and 1 mL of n-hexane were carried out. The organic extracts were pooled and filtered through a 0.22 μm membrane (GS ester cellulose), evaporated in vacuo at 200 mmHg and 35°C and resuspended in 200 μL of n-hexane.
Analysis of the organic extract was performed in a ProStar high-performance liquid chromatograph (HPLC; Varian, Inc.) with a C18 column and a photodiode array (PDA) detector set at 282 nm. HPLC-grade solvents were used (Sigma). The conditions of elution and chromatographic analysis were set according to a previous report (De Carvalho et al., 2006). The peak areas which were used to quantify ergosterol values, were obtained from the chromatographic analysis of the ergosterol standards and from the calibration curve. This resulted in a linear regression and the formula:
ERG = 56051A+4E(06) |
where, ERG is ergosterol (ppm) and A is the peak area, with R2 = 0.9984.
Biomass production in SSF columns: Airflow and initial moisture levels for the SSF columnar system were optimized in a previous experiment (data not shown) and these conditions were used for subsequent respirometric and kinetic studies.
Wheat grains were ground in a knife mill and sieved to obtain particles with 0.8-2 mm diameter for use as the substrate for SSF. The granulometry was determined by a previous test which indicated that particles smaller or greater than 0.8-2 mm would be difficult for mycelium penetration and further growth. The substrate was soaked in water for 12 h and sterilized at 121°C for 50 min. The initial pH of the substrate was adjusted to 5.9±0.1 with 0.1 N HCl and the initial moisture was adjusted to 39% with sterile water mixed with the inoculum suspension at an inoculation rate of 5% (v/w). The inoculated substrate was packed into 12 columns (40-mm diameter and 200-mm height) with a bed height of 120 mm (volume, 150 mL). Each column was filled with 45 g of the inoculated substrate and incubated for 14 days at 30°C. The fermentation system consisted of glass columns immersed in a water bath at 30°C and an airflow of 0.22 N mL g-1 substrate min per column (milliliter of air in standard conditions, per gram of wet substrate, per minute) (Raimbault, 1998) (Fig. 1). The columns were closed at both ends with cotton plugs, connected to humidifiers and the aeration rate was adjusted twice a day using a rotameter (Cole-Parmer).
This procedure was performed simultaneously for kinetic studies of ergosterol and the corresponding biomass production (columns numbered from 1-10). The outlet gases were analyzed for online respirometry (columns 11 and 12). The data obtained in this analysis were used to determine the growth parameters of A. brasiliensis mycelium in SSF.
Ergosterol extraction and analysis from the SSF biomass: For the kinetic studies on biomass production, the entire columnar content from 1-10 was withdrawn at 0, 4, 18, 24, 36, 48, 72, 100, 196 and 302 h of fermentation and analyzed for pH, moisture and water activity. Samples (1.0 g) were vacuum-dried (200 mmHg, 40°C) for 12 h, pulverized in an electric mill and analyzed for ergosterol content using the same procedures described for ergosterol obtained by SF. All analyses were performed in duplicate and data of biomass and fermented wheat grains were presented in Dry Weight (DW).
Estimation of the correlation factor between ergosterol and biomass production: The data about ergosterol content from the biomass obtained by SF was used to correlate the biomass production from SSF from the ergosterol content of the samples.
Respirometric analysis: The respirometric analysis of A. brasiliensis growth on wheat grains was carried out in 2 columns (numbered 11 and 12 with initial moisture of 39% and airflow of 0.22 N mL/g/min). The outlet gases were passed through desiccant silica gel columns and analyzed by Gas Chromatography (GC) (Shimadzu GC-8A; Shimadzu Co., Japan). This system was linked to a program for chromatograph control and integration (Chroma Biosystèmes, France) followed by analysis with the Fersol mathematical model (Rodriguez-Leon et al., 1988). The GC system had a Porapak 80/100 column at 60°C with 2 m length, helium as the carrier gas, a thermal conductivity detector and a sample injection volume of 300 μL for GC. The gases used for system calibration were air, CO2 (0.0)/O2 (21.0)/N2 (79.0); mixture 1, CO2 (5.0)/O2 (5.0)/N2 (90.0) and mixture 2, CO2 (10.0)/O2 (15.0)/N2 (75.0). In these conditions, the retention time of each component was 0.95 min for CO2, 5.72 min for O2 and 8.02 min for N2. Exhaust gas analysis was performed every hour with 2 replicates and Oxygen Update Rate (OUR) and CO2 production results were collected to establish a relationship between the online respirometric parameters in the columnar bioreactor with forced air and the mycelium growth. The results of the respirometric analysis were averaged for the two columns used in the assay.
Kinetic parameters: The kinetic parameters were calculated as described by Pandey et al. (2001) and biomass production by ergosterol analysis was used to obtain its yield based on O2 consumption (YX/O) (g of biomass g-1 of consumed O2).
The same biomass data was correlated to dependent variables using Fersol software (Rodriguez-Leon et al., 1988) and all data received the suffix F. The CO2 production results, OUR determination and ergosterol content from the kinetic study during the fermentation period were used to correlate the biomass produced in SSF at a certain time (Xn)F which was calculated by assuming values for its yield based on O2 consumption (YX/O)F (g of biomass g-1 of consumed O2) and the biomass maintenance coefficient (mX)F using Fersol.
Analytical procedures: The pH of the solid substrate was determined with a digital pH meter by mixing 5 g of the sample with 50 mL of distilled water. Substrate moisture was determined following standard procedures (AOAC, 2002). Water activity (aw) was measured using AquaLab CX-2.
RESULTS
Correlation between ergosterol and the biomass obtained by SF: The ergosterol content of mushrooms can be correlated with the fungal biomass (Klamer and Baath, 2004). In the present study, the following correlation was obtained:
| (1) |
This value was used to calculate the biomass production in SSF using the ergosterol content produced by this fermentation method.
Correlation between ergosterol and the biomass obtained by SSF: The biomass in SSF was estimated using an indirect method because it is difficult to separate the biomass from the substrate. Indirect methods are based on measurement of cell components or respirometric balance while accounting for biomass composition and stoichiometry of the process (Soccol and Vandenberghe, 2003).
Ergosterol analysis of wheat grains without fermentation resulted in a very low value, i.e., 1.74 μg g-1 which was subtracted from the ergosterol content found in the fermented samples. The results obtained from samples from the 14th day of fermentation were expressed as:
| (2) |
The calculation of the fungi biomass per gram of fermented dried wheat grains was obtained from the correlation between Eq. 1 and 2 and resulted in a maximum value of 0.29 biomass g-1 of fermented wheat grains (DW) at the 14th day of fermentation.
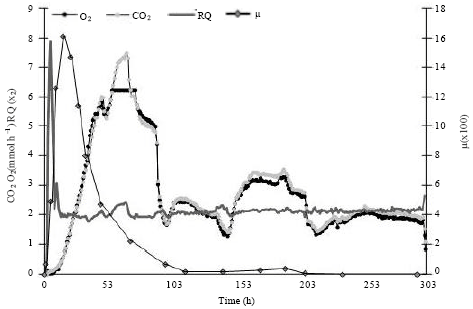 | |
| Fig. 2: | Respirometric data of Agaricus brasiliensis mycelium on wheat grains in the column, Column has 45 g of wheat grains and 39% initial moisture with forced aeration, RQ: Respiratory quotient and μ: specific growth rate |
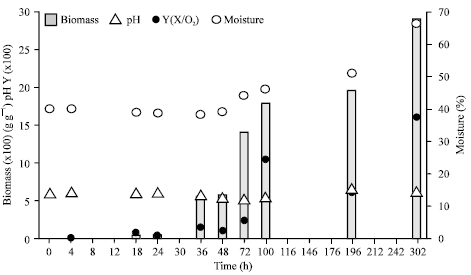 | |
| Fig. 3: | Growth parameters of Agaricus brasiliensis mycelium on wheat grains in the column, Column has 45 g of wheat grains and 39% initial moisture with forced aeration |
Kinetic studies of A. brasiliensis growth and the SSF biomass from respirometric analysis: The respirometric data and biomass production of A. brasiliensis mycelium in SSF are shown in Fig. 2 and 3, respectively. The results were obtained from a whole column containing 45 g of wheat grains with 39% initial moisture. The biomass yield coefficient was calculated as the ratio of biomass production per amount of O2 consumed and is presented in Fig. 3.
The Aw values ranged from 0.96-0.97 even with alterations in moisture content which increased gradually during the fermentation period (Fig. 3) and reached a value of 66% at the 14th fermentation day.
The initial pH value was ~6.0 and showed slight variation during the fermentation period (302 h, Fig. 3). The pH decreased slowly to 5.16 at 72 h of fermentation followed by an increase to values proximate to the initial pH (6.1) at the end of the fermentation period.
The data for biomass, produced CO2 and consumed O2 obtained during SSF were applied to the Fersol program and are presented in Table 1. The predicted biomass at a certain time (Xn)F involved the assumption of values for its yield based on the O2 consumption (YX/O)F and the coefficient of biomass maintenance (mx)F. The software allowed determination of the equation coefficients by successive approximation.
| Table 1: | Online respirometric parameters and biomass production by A. brasiliensis mycelium by solid-state culture in a column coupled with forced aeration |
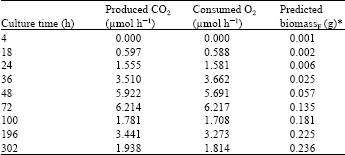 | |
| *The values of time, biomass, produced CO2 and consumed O2 were used to estimate biomassF production in the column (Wheat grains: 45 g, Initial moisture: 39%, Air flow: 0.22 N mL g-1 substrate/min per column, Diameter: 40 mm and Bed height: 120 mm) with forced aeration through the Fersol program | |
Based on the values of fermentation time and biomass obtained from ergosterol correlation and consumed O2, the following data were obtained using Fersol: predicted biomass yield (YX/O)F = 1.949 g of biomass g-1 of consumed O2; coefficient of biomass maintenance (mx)F = 0.0027 g of consumed O2/(g of produced biomass. h); specific growth rate (μF) = 0.086/h and correlation coefficient (R2F) = 0.955.
The development of the fungal mycelia was described according to the RQ values, O2 consumption and CO2 and biomass production (Fig. 2, 3). The ergosterol detection was very low at the 4th h with 0.006 mg g-1 of wheat grains and the production of CO2 started at the sixth hour of fermentation. From 6 h to 12 h, the RQ value increased and reached a maximum of 3.94 at 11 h of fermentation (Fig. 2) which resulted from higher CO2 production and little O2 uptake in the system. After 12 h, the RQ value decreased to ~1, indicating aerobic respiratory activity according to the theoretical value expected for this parameter (Fig. 2). CO2 production was associated with O2 consumption. During the growth phase, the mycelium spread over the substrate surface, showing a white cover resembling cotton. Ergosterol production began to increase at the 18th h of fermentation, with the values varying from 0.03-0.94 mg g-1 of fermented wheat grains (dry weight) at the 72nd h (Fig. 3). The O2 uptake rate and CO2 production were very high between 57 and 76 h, indicating intense metabolism of the fungus (Fig. 2). The maximum CO2 production occurred at 69 h of fermentation, reaching the maximum rate of 7 mmol h-1. The development of A. brasiliensis during this phase resulted in a maximum specific growth velocity of 0.016 h-1 at 18 h. CO2 production decreased after 78 h, whereas O2 consumption increased gradually until 98 h. During the fermentation periods from 105-126 h and from 156-213 h, an increase in CO2 production and O2 consumption was observed (Fig. 2). Ergosterol analysis resulted in a continuous increase with the maximum value achieved at the 14th fermentation day, at 302 h, with 1.95 mg g-1 of fermented wheat grains (dry weight) (Fig. 3).
DISCUSSION
Biotechnological applications of SSF are widespread and are one of the oldest processes known for production of foods such as tempeh, koji processing, cheese production and many secondary metabolites such enzymes and more recently fermentable sugars aiming the production of bioethanol (Anusha et al., 2012; Hong et al., 2011; Murad and Azzaz, 2010; Sherief et al., 2010; Pandey et al., 2001). However, several technical problems related to SSF have not been solved during the scale-up process, such as difficulty controlling temperature, pH, moisture, substrate concentration, or pO2 during cultivation under limited water availability (Holker et al., 2004; Singhania et al., 2009). The observed increase in the moisture levels from initial values of 39% to 66% at the 14th day of SSF in the kinetic study (Fig. 3) was possibly due to the humidity of the air flushed through the columnar system. In contrast, SSF performed in trays by Dalla-Santa et al. (2010) resulted in maintenance or reduction of the moisture levels when fermentation was conducted in humidified or normal air conditions, respectively.
It is also difficult to maintain pH values during SSF, as observed by the varying pH values during fermentation (Fig. 3) which seems characteristic of this microorganism. Similar changes were detected in another study of SF with A. brasiliensis, with pH values varying from 5.6-4.4 on the second day and reaching 7.84 on the sixth day (Gern, 2005) which the author suggested was correlated with acid production and concomitant glucose consumption. The pH variance was more intense in the above-mentioned study than in our study which is presumably due to the differences in the mycelium grown, nutrient absorption related to the fermentation type (SF versus SFF) and to the chemical difference of the substrate (liquid complex medium versus wheat grains). The wheat grains used in the present study are rich in protein and it is well known that protein-rich substrates can act as buffers and inhibit excessive pH changes (Chisti, 1999).
It is important to follow the kinetics of growth in relation to the metabolic activity in order to measure the biomass in SSF. In SSF, the microorganisms are closely bound to the solid-state matrix, resulting in difficult biomass separation and measurement. Therefore, the biomass is usually measured indirectly with minor or major error by determining cell components such as glucosamine, ergosterol, proteins, or nucleic acids and the development of adequate monitoring and control processes are required (Pandey et al., 2001; Chen et al., 2005). An advantage of determining ergosterol content is that it can be recovered and separated by HPLC and easily quantified by spectrophotometry, providing a sensitive biomass index at low levels of growth (Raimbault, 1998). On the other hand, estimation of O2 intake and CO2 evolution rates are recently considered the most accurate determination of microorganism growth (Pandey et al., 2007). Therefore, it was used the ergosterol determination with a final value of 0.29 g of biomass per gram of fermented wheat grains (dry weight) (Fig. 3) and the respirometric analysis (Fig. 2, Table 1) and both methods efficiently measured and estimated fungi biomass from A. brasiliensis in the grounded wheat grains in SSF.
Owing to the lack of published research on A. brasiliensis in SSF, the results obtained in the present work were compared with those obtained with Monascus, since both these fungi have similar habitats; however, the fact that each fungus has a specific metabolism was also considered. A pre-fermentation procedure that includes increasing the inoculum concentration or inoculation ratio has been suggested to reduce the long lag phase of 60 h observed in Monascus (De Carvalho et al., 2006). In the present study, use of the broken mycelium suspension resulted in an adaptive phase of less than 6 h of fermentation, with CO2 production at 6 h and an increased RQ value. This short lag phase is important for 2 reasons: it reduces contamination which is one of the most recurrent problems in spawn production and the fermentation time is reduced which is very important at the industrial scale. In normal spawn production, pieces of solid medium covered with developed mycelium are inoculated on grains, followed by incubation for 20-30 days, whereas the broken suspension method usually enables the efficient development of mushrooms in SSF within 14-18 days. In this study, this method resulted in extremely rapid development of A. brasiliensis. After 4 days, the entire substrate was recovered by the fungal mycelium. Active fermentation of the fungus was detected from the high and rapid increase in the RQ value (3.94) for A. brasiliensis at 11 h, whereas the maximum RQ value for Monascus was <2 in another study (De Carvalho et al., 2006).
Because there is a dearth of studies on Agaricus for comparison of the growth rate based on O2 consumption and CO2 production, the results were compared with others published for other Ascomycetes or Basidiomycetes in other fermentations systems. Monascus sp., developed in the same type of columnar system (De Carvalho et al., 2006) reached 0.086/h of maximum specific growthF at 140 h, whereas A. brasiliensis showed intense growth and specific growth velocity of 0.016/h at 18 h, as also observed by the total and rapid coverage of mycelium on the wheat particles. Gibberella fujikuroi and gibberellic acid (GA3) production using coffee husk and cassava bagasse in a column bioreactor resulted in a maximum specific growth rateF of 0.052 h-1 between 24 and 48 h of fermentation (Machado et al., 2004).
The yield and biological efficiency for all microorganisms depend on the substrate formulations and fermentation parameters. Many authors specify the importance of the absolute biomass amount for the calculation of growth rates and yields in fermentation (Raimbault, 1998). Ergosterol was used to calculate the biomass and the possible yield which resulted in a biomass yield (YX/O) of 0.160 g of biomass g-1 of consumed O2 at 302 h of fermentation. Soccol and Vandenberghe (2003) obtained good correlations between the values obtained experimentally for the OUR and CO2 involved with the analyzed biomass through protein content analysis using the Fersol program. In the present work it was chosen to use ergosterol determination rather than protein analyses as the high protein content in the wheat grains could induce error because of the possible incorporation of wheat protein into the microorganism. On the other hand, yield can be calculated based on product yield using ergosterol production per amount of consumed O2 (Fig. 3) and the maximum Y(P/O2) obtained value was 1.07 mg of ergosterol g-1 of dried fermented wheat grains mmol-1 O2 at the 14th fermentation day.
The mushroom fruiting body contains remarkably high amounts of phytosterol, the most important of which is ergosterol, whose distribution varies in species and in different parts of the mushroom tissue (Jasinghe and Perera, 2005; Krzyczkowski et al., 2009). For example, variances in the overall ergosterol content of the fruiting body of different types of mushrooms have been reported as follows (mg of ergosterol g-1 of dry weight [DW]): 7.80 (0.35) in button mushrooms or Agaricus, 6.05 (0.07) in shiitake, 4.40 (0.08) in oyster mushrooms and 0.68 (0.14) in enoki mushrooms (Jasinghe and Perera, 2005). Mattila et al. (2002) observed a value of 6.79 mg g-1 DW of ergosterol in shiitake mushrooms and Teichmann et al. (2007) reported that ergosterol contents of dark cultivated mushrooms (A. bisporus/white, A. bisporus/brown, A. bisporus/Portabella, Lentinula edodes and Pleurotus ostreatus) varied from 3.7-5.1 mg g-1 DW to 40-108 mg/100 (Fresh Weight). Krzyczkowski et al. (2009) detected ergosterol peroxide values of 29.32, 17.27 and 12.60 mg/100 g for Boletus edulis (king bolete), Suillus bovinus (Jersey cow mushroom) and B. badius (bay bolete) fruiting bodies, respectively. These authors additionally described the ergosterol values for mycelia for several mushrooms: H. erinaceum, Laetiporus sulphureus (chicken mushroom) and Morchella esculenta (common morel) with 15.98, 10.07 and 13.37 mg/100 g, respectively. Few articles have described the biomass and ergosterol content of the mycelium from the mushroom A. brasiliensis. Li and Shen (2003) used a mixture of malt, yeast extract, glucose, peptone, oatmeal and calcium carbonate as the substrate for mycelium production of A. brasiliensis in SSF (%: malt extract, 2; yeast extract, 2; peptone, 0.5; glucose, 2; water, 45.7, oatmeal, 45.7; calcium carbonate, 2) and obtained ergosterol values ranging from 0.371-0.416 mg g-1 of dry substrate. In the present study, an increase in the ergosterol content of the fermented wheat grains was observed during the fermentation period which reached the maximum value of 1.95 mg of ergosterol per gram of dry fermented material after 302 h (day 14) of fermentation. This result was presumably due to the efficient aeration system which provided high O2 diffusion in the substrate and adequate nutrient content in the wheat grains.
In our study, submerged fermentation yielded a value of 6.71 mg of ergosterol per gram of dried biomass with 20 g of glucose in the Erlenmeyer flask. This value is similar to a study by Zou (2006), reporting a biomass-ergosterol production of 8.8 mg g-1 of DW in an Erlenmeyer flask and 25 mg g-1 DW at 144 h in fed batch fermentation by SF of A. brasiliensis using 20 g of sugar in the medium. Gao and Gu (2007) optimized the ergosterol production of A. brasiliensis in SF and achieved 0.076 g L-1, an increase of 43% compared to non-optimized conditions.
The importance of the present results is related to the recognized beneficial effects of ergosterol on health improvement and tumor prevention (Takaku et al., 2001), the capacity of phytosterols to lower plasma cholesterol and LDL cholesterol (Shin et al., 2003) and the proven beneficial effects of ingestion of wheat grains fermented with A. brasiliensis mycelium in mice by our research group (Dalla-Santa et al., 2009; Dalla-Santa et al., 2010) which suggest that this fermented material is an excellent bioactive ingredient. Moreover, SSF is a cheap and simple method to obtain this functional product. The results presented herein allow calculation of the fungi biomass percentage in solid-state-fermented wheat grains and determination of the growth parameters of the main mycelium A. brasiliensis in this substrate which can be milled to obtain bio-flour, nutraceutical foods.
CONCLUSION
In summary, regarding the results of biomass from ergosterol production in SSF, both ergosterol content and respirometric activity determination methods efficiently estimated the biomass production of A. brasiliensis in SSF. The growth parameters achieved a specific growth velocity of 0.016/h at 18 h and a biomass yield (YX/O) of 0.160 g of biomass g-1 of consumed O2 at 302 h of fermentation. Ergosterol analysis revealed that 6.71 mg of ergosterol g-1 of dried fungi biomass was obtained from SF and 1.95 mg of ergosterol/g of solid-state-fermented wheat grains (dry weight) from SSF. The correlation with biomass production allowed an estimate of 0.29 g of fungi biomass per gram of the solid-state-fermented wheat grains (dry weight) which represent an excellent bioactive material for nutraceutical products due the presence of the ergosterol with recognized beneficial effects.
ACKNOWLEDGMENTS
H.S. Dalla-Santa and C.R. Soccol thank the Midwest State University (UNICENTRO) and the National Council of Technological and Scientific Development (Project No. 475798/2003-1), respectively, for financial support and for a scholarship under the Scientific Productivity Scheme.
REFERENCES
- Anusha, N.C., M.S. Umikalsom, T.C. Ling and A.B. Ariff, 2012. Relationship between fungal growth morphologies and ability to secrete lipase in solid state fermentation. Asian J. Biotechnol., 4: 15-29.
CrossRef - Camelini, C.M., M. Maraschin, M.M. de Mendonca, C. Zucco, A.G. Ferreira and L.A. Tavares, 2005. Structural characterization of β-glucans of Agaricus brasiliensis in different stages of fruiting body maturity and their use in nutraceutical products. Biotechnol. Lett., 27: 1295-1299.
CrossRefDirect Link - De Carvalho, J.C., A. Pandey, B.O. Oishi, D. Brand, J.A. Rodriguez-Leon and C.R. Soccol, 2006. Relation between growth, respirometric analysis and biopigments production from Monascus by solid-state fermentation. Biochem. Eng. J., 29: 262-269.
CrossRefDirect Link - Dalla-Santa, H.S., R. Rubel, F.M.D. Vitola, F. Leifa and A.L. Tararthuch et al., 2009. Kidney function indices in mice after long intake of Agaricus brasiliensis mycelia (=Agaricus blazei, Agaricus subrufescens) produced by solid state cultivation. Online J. Biol. Sci., 9: 21-28.
Direct Link - Dalla-Santa, H.S., R. Rubel, L.C. Fernandes, S.J.R. Bonatto and S.R. Bello et al., 2010. Agaricus brasiliensis-enriched functional product promotes in mice increase in HDL levels and immunomodulate to Th1 CD4+T subsets. Curr. Trends Biotechnol. Pharm., 4: 957-970.
Direct Link - Fan, L., A.T. Soccol, A. Pandey, S. Germano, R. Rau, A. Pedroso and C.R. Soccol, 2003. Production of polysaccharide by culinary-medicinal mushroom Agaricus brasiliensis S. Wasser et al., LPB 03 (Agaromycetidae) in submerged fermentation and its antitumor effect. Int. J. Med. Mushr., 5: 17-23.
Direct Link - Gao, H. and W.Y. Gu, 2007. Optimization of polysaccharide and ergosterol production from Agaricus brasiliensis by fermentation process. Biochem. Eng. J., 33: 202-210.
CrossRefDirect Link - Hamedi, A., H. Vahid and F. Ghanati, 2007. Optimization of the medium composition for production of mycelial biomass and exo-polysaccharide by Agaricus blazei Murill DPPh 131 using response-surface methodology. Biotechnology, 6: 456-464.
CrossRefDirect Link - Holker, U., M. Hofer and J. Lenz, 2004. Biotechnological advantages of laboratory-scale solid state fermentation with fungi. Applied Microbiol. Biotechnol., 64: 175-186.
Direct Link - Hong, L.S., D. Ibrahim and I.C. Omar, 2011. Lignocellulolytic materials-as a raw material for the production of fermentable sugars via solid state fermentation. Asian J. Sci. Res., 4: 53-61.
CrossRefDirect Link - Chen, F., Y. Jiang and F. Ouyang, 2005. Development of bioprocess engineering in China. Biotechnology, 4: 1-6.
CrossRefDirect Link - Jamal, P., O.K. Saheed and Z. Alam, 2012. Bio-valorization potential of banana peels (Musa sapientum): An overview. Asian J. Biotechnol., 4 : 1-14.
CrossRefDirect Link - Jasinghe, V.J. and C.O. Perera, 2005. Distribution of ergosterol in different tissues of mushrooms and its effect on the conversion of ergosterol to vitamin D2 by UV irradiation. Food Chem., 92: 541-546.
CrossRefDirect Link - Karmakar, M. and R.R. Ray, 2010. Extra cellular endoglucanase production by rhizopus oryzae in solid and liquid state fermentation of agro wastes. Asian J. Biotechnol., 2: 27-36.
CrossRef - Klamer, M. and E. Baath, 2004. Estimation of conversion factors for fungal biomass determination in compost using ergosterol and PLFA 18:2ω6,9. Soil Biol. Biochem., 36: 57-65.
CrossRefDirect Link - Krzyczkowski, W., E. Malinowska, P. Suchocki, J. Kleps, M. Olejnik and F. Herold, 2009. Isolation and quantitative determination of ergosterol peroxide in various edible mushroom species. Food Chem., 113: 351-355.
CrossRefDirect Link - Fan, L., A.T. Soccol, A. Pandey and C.R. Soccol, 2007. Effect of nutritional and environmental conditions on the production of EPS of Agaricus blazei by submerged fermentation and its antitumor activity. Food Sci. Technol., 40: 30-35.
CrossRef - Lima, L.F.O., S. Habu, J.C Gern, B.M. Nascimento and J.L. Parada et al., 2008. Production and characterization of the exopolysaccharides produced by Agaricus brasiliensis in submerged fermentation. Applied Biochem. Biotechnol., 151: 283-294.
CrossRefDirect Link - Machado, C.M.M., B.O. Oishi, A. Pandey and C.R. Soccol, 2004. Kinetics of Gibberella fujikuroi growth and gibberellic acid production by solid-state fermentation in a packed-bed column bioreactor. Biotechnol. Prog., 20: 1449-1453.
CrossRefDirect Link - Mattila, P.H., A.M. Lampi, R. Ronkainen, J. Toivo and V. Piironen, 2002. Sterol and vitamin D2 contents in some wild and cultivated mushrooms. Food Chem., 76: 293-298.
CrossRefDirect Link - Murad, H.A. and H.H. Azzaz, 2010. Cellulase and dairy animal feeding. Biotechnology, 9: 238-256.
CrossRefDirect Link - Okwulehie, I.C., N.C. Princewill and O.C. Johnpaul, 2007. Pharmaceutical and nutritional prospects of two wild macro-fungi found in Nigeria. Biotechnology, 6: 567-572.
CrossRefDirect Link - Onyango, B.O., V.A. Palapala, P.F. Arama, S.O. Wagai and B.M. Gichimu, 2011. Suitability of selected supplemented substrates for cultivation of Kenyan native wood ear mushrooms (Auricularia auricula). Am. J. Food Technol., 6: 395-403.
CrossRefDirect Link - Pal, U.K., P.K. Mandal, V.K. Rao and C.D. Das, 2011. Quality and utility of goat milk with special reference to India: An overview. Asian J. Anim. Sci., 5: 56-63.
CrossRefDirect Link - Raimbault, M., 1998. General and microbiological aspects of solid substrate fermentation. Electron. J. Biotechnol., 1: 174-188.
Direct Link - Regulapati, R., N.P. Malav and N.S. Gummadi, 2007. Production of Thermostable α-amylases by solid state fermentation: A review. Am. J. Food Technol., 2: 1-11.
CrossRefDirect Link - Ribeiro, L.R. and D.M.F. Salvadori, 2003. Dietary components may prevent mutation-related diseases in humans. Mutat. Res., 544: 195-201.
Direct Link - Rodriguez-Leon, J.A., L. Sastre, J. Echevarria, G. Delgado and W.A. Bechstedt, 1988. A mathematical approach for the estimation of biomass production rate in solid state fermentation. Acta Biotechnol., 8: 307-310.
CrossRefDirect Link - Rubel, R., H.S. Dalla-Santa, L.C. Fernandes, J.H.C. Lima-Filho and B.C. Figueiredo et al., 2008. High immunomodulatory and preventive effects against sarcoma 180 in mice fed with Ling Zhi or Reishi mushroom Ganoderma lucidum (W. Curt.: Fr.) P. Karst. (Aphyllophoromycetideae) mycelium. Int. J. Med. Mushrooms, 10: 37-48.
CrossRef - Rubel, R., H.S. Dalla-Santa, L.C. Fernandes, S.J.R. Bonatto and S. Bello et al., 2010. Hypolipidemic and antioxidant properties of Ganoderma lucidum (Leyss:Fr) Karst used as a dietary supplement. World J. Microbiol. Biotechnol., 27: 1083-1089.
CrossRefDirect Link - Sherief, A.A., N.E.A. El-Naggar and S.S. Hamza, 2010. Bioprocessing of lignocellulosic biomass for production of bioethanol using thermotolerant Aspergillus fumigates under solid state fermentation conditions. Biotechnology, 9: 513-522.
CrossRefDirect Link - Shin, M.J., S.J. Rim, Y. Jang, D. Choi and S.M. Kang et al., 2003. The cholesterol-lowering effect of plant sterol-containing beverage in hypercholesterolemic subjects with low cholesterol intake. Nutr. Res., 23: 489-496.
CrossRefDirect Link - Singhania, R.R., A.K. Patel, C.R. Soccol and A. Pandey, 2009. Recent advances in solid-state fermentation. Biochem. Eng. J., 44: 13-18.
CrossRefDirect Link - Soccol, C.R. and L.P.S. Vandenberghe, 2003. Overview of applied solid-state fermentation in Brazil. Biochem. Eng. J., 13: 205-218.
CrossRefDirect Link - Stamets, P., 2000. Techniques for the cultivation of the medicinal mushroom royal sun Agaricus blazei Murrill (agaricomycetidae). Int. J. Med. Mushr., 2: 151-160.
Direct Link - Stamets, P., 2005. Notes on nutritional properties of culinary-medicinal mushrooms. Int. J. Med. Mushrooms, 7: 103-110.
CrossRefDirect Link - Takaku, T., Y. Kimura and H. Okuda, 2001. Isolation of an antitumor compound from Agaricus blazei Murril and its mechanism of action. J. Nutr., 131: 1409-1413.
Direct Link - Tang, Y.J. and J.J. Zhong, 2002. Fed-batch fermentation of Ganoderma lucidum for hyperproduction of polysaccharide and ganoderic acid. Enzym. Microb. Technol., 31: 20-28.
CrossRefDirect Link - Teichmann, A., P.C. Dutta, A. Staffas and J. Margaretha, 2007. Sterol and vitamin D2 concentrations in cultivated and wild grown mushrooms: Effects of UV irradiation. LWT-Food Sci. Technol., 40: 815-822.
CrossRefDirect Link - Wasser, S.P., 2011. Current findings, future trends and unsolved problems in studies of medicinal mushrooms. Applied Microbiol. Biotechnol., 89: 1323-1332.
CrossRefDirect Link - Weete, J.D., 1973. Sterols of the fungi: Distribution and biosynthesis. Phytochemistry, 12: 1843-1864.
CrossRefDirect Link - Zou, X., 2005. Effects of Zn supplementation on the growth, amino acid composition, polysaccharide yields and anti-tumour activity of Agaricus brasiliensis. World J. Microbiol. Biotechnol., 21: 261-264.
CrossRefDirect Link - Zou, X., 2006. Fed-batch fermentation for hyperproduction of polysaccharide and ergosterol by medicinal mushroom Agaricus brasiliensis. Process. Biochem., 41: 970-974.
CrossRefDirect Link