
M. Tharek
School of Biological Sciences, Universiti Sains Malaysia, 11800 Minden, Penang, Malaysia
K. Dzulaikha
School of Biological Sciences, Universiti Sains Malaysia, 11800 Minden, Penang, Malaysia
S. Salwani
School of Biological Sciences, Universiti Sains Malaysia, 11800 Minden, Penang, Malaysia
H.G. Amir
School of Biological Sciences, Universiti Sains Malaysia, 11800 Minden, Penang, Malaysia
N. Najimudin
School of Biological Sciences, Universiti Sains Malaysia, 11800 Minden, Penang, Malaysia
Biotechnology
Year: 2011 | Volume: 10 | Issue: 6 | Page No.: 521-527







ABSTRACT
The aims of this study were to isolate endophytic diazotrophs from leaf tissues of oil palm (Elaeis guineensis Jacq.) and to understand the endophytic migration and colonization ability in a selected host plant (rice seedlings; Oryza sativa var. MR220). Moreover, observing the production of plant-cell wall degrading enzymes would aid in the understanding of the endophytic migration. These enzymes are vital for endophytes invasion and spread within the host plant. The endophytic diazotrophs were isolated and identified via partial 16S rDNA sequence analyses. The N2-fixing ability and IAA production of the isolates were determined by Acetylene Reduction Assay (ARA) and Salkowski’s colorimetric test. A total of six isolates were tested positive for N2 fixation potential (2.5x10-10 to 8.5x10-7 μmol C2H4 cfu-1 h-1) and IAA production. The isolates were identified as Enterobacter sp., Acinetobacter sp., Azospirillum sp., Gluconacetobacter sp. and Phytobacter sp. Ascending migration of the endophyte from roots to leaves of the host was also proven via microscopic observation of Green Fluorescent Protein (GFP)-tagged Enterobacter sp. and production of plant-cell wall degrading enzymes. Thus, this study successfully proven occurrence of culturable endophytic diazotrophs in leaf tissues of oil palm. The endophytic diazotrophs were also classified as high-potential plant growth enhancers through the ability to fix nitrogen, produce phytohormone and the potential for endophytic migration and colonization inside the host plants.
PDF Abstract XML References Citation
Received: June 22, 2011;
Accepted: November 23, 2011;
Published: December 12, 2011
How to cite this article
M. Tharek, K. Dzulaikha, S. Salwani, H.G. Amir and N. Najimudin, 2011. Ascending Endophytic Migration of Locally Isolated Diazotroph, Enterobacter sp. Strain USML2 in Rice. Biotechnology, 10: 521-527.
DOI: 10.3923/biotech.2011.521.527
URL: https://scialert.net/abstract/?doi=biotech.2011.521.527
DOI: 10.3923/biotech.2011.521.527
URL: https://scialert.net/abstract/?doi=biotech.2011.521.527
INTRODUCTION
Endophytic diazotrophic bacteria reside within host plant tissues and are relatively protected from the competitive and high stress environment of the soil. Several endophytic diazotrophs, such as Enterobacter dissolvens, Brevundimonas aurantiaca, Pantoea agglomerans, Pseudomonas sp., Acetobacter diazotrophicus, Azoarcus sp. and Herbaspirillum seropedicae have been successfully isolated from the interior tissues of roots and/or shoots and leaves of different kinds of graminaceous crops (Barraquo et al., 1997; Bacilico-Jimenz et al., 2001; Aslin et al., 2005). These endophytes are capable of spreading systemically via invasion of inner tissues including xylem vessels (Prakamhang et al., 2009). Interest in them has arisen due to their ability to supply substantial amounts of biologically-fixed N to their host plants (Quispel, 1991; Pedraza et al., 2009). This biological reaction which assembles inside the host plant, not only balances the loss of N from soils or ecosystems via the nitrogenase enzyme reduction of gaseous N2 to ammonia (NH3) but also provides a protected self-sustaining N system. In this system, the endophytic diazotrophs control the supply of N and very little or no loss of N occurs. Therefore, it would increase the potential for N supply to the plant due to direct availability of fixed N (Senthilkumar et al., 2008). Other studies have also shown that efficient N2-fixing activities occur in the symbiotic association between endophytic diazotrophs and legumes (Bacilico-Jimenz et al., 2001; Al-Falih, 2002; Mia and Shamsuddin, 2010).
Interaction between endophytic diazotrophs and host plants can also have a profound effect on crop health and yield (Alam et al., 2003). This interaction includes inducing plant defense mechanisms, producing antibiotics, out-competing pathogens and providing growth-stimulating substances such as phytohormones and enzymes (Jolly et al., 2010). In addition, endophytic diazotrophs often enhance plant growth promotion compared to bacteria restricted to the rhizosphere and root surface (Compant et al., 2005; Sofi and Wani, 2007). Since endophytic diazotrophs are beneficial for use as inocula and mostly originate from soil (Taghavi et al., 2009), understanding their ascending migration ability from roots to leaves is therefore crucial. The localization of the endophytes in plant tissues can be studied in detail with the help of reporters such as Green Fluorescent Protein (GFP) which has been proven to be a simple and effective marker system for studying plant-microbe interactions (Singh et al., 2009). Moreover, observing the production of plant-cell wall degrading enzymes such as endoglucanase and endopolygalacturonase would aid in the understanding of endophytic migration, as these enzymes are vital for invasion and spread within the host plant (Reinhold-Hurek et al., 2006; Hardoim et al., 2008). Due to these interests, the aims of this study were to isolate endophytic diazotrophs from inner leaf tissues, to identify potential isolates for N2 fixation, phytohormone production and to observe endophytic migration and colonization ability of diazotrophs in the host plant.
MATERIALS AND METHODS
Isolation of endophytic diazotrophs: The study was conducted from July 2005 to Dec 2010 at School of Biological Sciences and School of Dental Sciences, Universiti Sains Malaysia. Endophytic diazotrophs were isolated from the internal leaf tissues of oil palm (Elaeis guineensis Jacq.). The leaf samples were surface-sterilized with 50 mL of sodium hypochlorite solution (75%, v/v) for 10 sec, 25 mL of ethanol (95%, v/v) for 30 sec and washed twice with 50 mL of sterile distilled water for 30 sec to ensure that they were free of any contaminants. The leaves were crushed with a knife and forceps to expose the endophytic diazotrophs (Aslin et al., 2005). Each crushed sample was serially diluted with saline solution and later inoculated into vials containing 10 mL of N-free malate (Nfb) semi-solid medium (Moutia et al., 2003; Aslin et al., 2005). Presence of diazotrophs was indicated by the formation of pellicles in Nfb semi-solid medium after 72 h of incubation. A loopful of pellicle was streaked onto Red Congo solid media and incubated under aerobic conditions at 30°C for 48 h. Gram stain and motility of the pure isolates were then observed. The pure isolated colonies were tested for their ability to fix N2 on the basis of Acetylene Reduction Assay (ARA) and to produce phytohormone via Salkowski’s colorimetric test.
Acetylene reduction assay (ARA): The N2-fixing ability of each isolate was determined by Acetylene Reduction Assay (ARA). Each isolate was inoculated into 30 mL airtight bottles containing 10 mL Nfb semi-solid medium. After 72 h of incubation, 5% (v/v) of the head space air was replaced with acetylene gas (99.8% purity) and the vials were reincubated for 24 h to complete reduction of acetylene to ethylene (Prakamhang et al., 2009). The samples were assayed for ethylene using a gas chromatography system (G-3000 Hitachi Ltd., Japan) fitted with a carboxen 1004 micro pack, 2mx1/16 inch (0.16 cm) stainless steel column and Flame Ionization Detector (FID). The N2-fixation activity was determined from the ethylene concentrations recorded and the viable cell counts of the isolates tested (μmol C2H4 cfu-1 h-1) (Aslin et al., 2005).
Phytohormone assay: Ability of the diazotrophs to produce IAA was determined via Salkowski’s colorimetric test. The isolates were cultured in Nfb medium supplemented with 0.5 g L-1 tryptophan. The bacterial cultures were harvested after 48 h of incubation and centrifuged at 15000 rpm for 15 min. The supernatant (2 mL) was mixed with 3 mL of Salkowski’s reagent (150 mL concentrated H2SO4+250 mL distilled H2O+7.5 mL of 0.5 M FeCl3) and left in the dark for chemical reactions and color development for 30 min (Patten and Glick, 2002). The color intensity was measured by spectrophotometer at 530 nm wavelength.
Identification of endophytic diazotrophs: Endophytic diazotrophs were identified via partial 16S rDNA sequence analyses. Amplification of partial 16S rDNA sequence was performed using forward primer UP2 5’-GGG CCC CCG YCA ATT CCT TTG ART TT-3’ and reverse primer URP 5’-GTG CCA GCM GCC GCG GTA A-3’, as described by Bavykin et al. (2004). The presence of PCR products was visualized by electrophoresis on agarose gels (1%, v/v) stained with ethidium bromide and purified by using QIAquick (Qiagen) following the manufacturer’s instructions. Purified PCR product was ligated with pGEM-T Easy Vector Systems (Promega, USA), transformed into E. coli JM109 and cultured on Luria-Bertani agar media consisting of ampicillin (100 mg mL-1), IPTG (0.1 M) and X-Gal (50 μg mL-1). Recombinant plasmids from white colony screening were extracted using Plasmid Miniprep Kit Promega, USA. The insert (partial 16S rDNA gene) was confirmed by digestion using Not1 and EcoR1 restriction enzymes (Choo et al., 2003). Recombinant plasmids containing the partial 16S rDNA gene were sequenced by Macrogen Laboratories (Seoul, South Korea) and referred to the gene bank database at National Center for Biotechnology Information (NCBI) via nucleotide-nucleotide Basic Local Alignment Search Tool (BLASTn) program to identify the genus and species of the isolated strains.
Fluorescent labeling of endophytic diazotrophs by electroporation: DNA sequence encoding the Green Fluorescent Protein (GFP) was incorporated into the isolated endophytic diazotrophs to confirm their endophytic ability. The isolates were cultured in 10 mL of nutrient broth. A sample (1 mL) of each subculture was transferred into a 1 L conical flask containing 200 mL of nutrient broth medium and incubated at 30°C with shaking for 48 h. Cells at stationary phase were chilled on ice separately and harvested by centrifugation at 3 000 x g for 10 min at 4°C. Each pellet was then washed twice with 200 mL of cold, sterilized distilled water and resuspended in 500 μL of cold, sterile 10% glycerol-water. The cell suspension was dispensed in 100 μL aliquots and stored at -80°C until use. The competent cells (100 μL) were thawed on ice and 200 ng of purified GFP delivery plasmid pGFPCR was added to the cells and mixed quickly. The mixture was then incubated on ice for 15 min and transferred to a sterile, prechilled cuvette (0.2 cm interelectrode gap) and placed in the Gene Pulser II apparatus equipped with a Pulser Controller (Bio-Rad Laboratories, Tokyo, Japan). The electroporation unit was used at the following settings: 12.5 kV cm-1, 25 μF and 200 Ω. Following the pulse, the cells were immediately diluted with 1 mL of nutrient broth medium, transferred to the sterilized tubes and incubated at 30°C for 3 to 4 h. From each tube, 100 μL of culture cells was plated onto selective nutrient agar plates containing 100 μg of ampicillin per mL. After the plates were incubated for 48 h at 30°C, green fluorescence emission was examined under a UV transilluminator.
Ascending endophytic migration of diazotroph: In studying the colonization process of ascending migration ability from roots to leaves, rice seedling was used as a model system. In this study, endophytic diazotrophs that had been successfully tagged with Green Fluorescent Protein (GFP) were used as inoculums (108 cells mL-1). The inoculum (2 mL per respective seedling) was inoculated on the lower portion of the roots of each 7-day-old rice seedling (Oryza sativa var. MR220) which was then transferred into a 50 mL test tube containing 1 mL of N-free liquid medium under gnotobiotic (sterile) conditions. At 12 h and 24 h after inoculation, the seedlings were harvested and surface sterilized with 70% ethanol for 30 s to examine the in planta colonization of the endophyte (GFP-tagged-Enterobacter sp. strain USML2). Colonization was observed by viewing the excised roots, stem bases and leaf tips with a Leica Microsystems TCS SP2 Confocal Laser Scanning Microscope (CLSM). The GFP-tagged bacteria were excited with a 488 nm Argon laser line (Verma et al., 2004). The strain was then assayed for plant-cell wall degrading enzyme production to observe its capability to invade and migrate in the plant tissues.
Plant-cell wall degrading enzyme production: Production of plant-cell wall degrading enzymes (endoglucanase and endopolygalacturonase) was assayed in triplicates using modified methods by Compant et al. (2005). Kim-Wimpenny (KW) solid medium with D-glucose, containing either 0.2% (w/v) carboxymethyl cellulose (CMC) or 0.5% polygalacturonic acid, was spot inoculated with the selected isolate and incubated at 30°C for 3 days. Bacterial cells were then removed from the plates by scraping the colonies with a sterile loop. The surface of the agar was then rinsed with sterile distilled water to wash away the scraped colonies. CMC-containing plates were stained with Congo Red (0.1%; w/v) for 30 min. This was followed by several washes with 1 M sodium chloride (NaCl) solution to improve the contrast. Similarly, the polygalacturonic acid-containing plates were stained with ruthenium red (0.1%) and washed with 1 M NaCl solution. Presence of endoglucanase activity was determined by the appearance of a clear yellowish halo around the inoculated area. Endopolygalacturonase activity was observed by the appearance of intense purple-red halos on a colourless background at the point (spot) where the bacteria were inoculated.
RESULTS
Isolation and characterization of endophytic diazotrophs: A total of 14 Gram-positive and 16 Gram-negative endophytic bacteria were isolated from surface sterilized leaves of oil palm (data not shown). All strains formed veil-like pellicles in semi-solid Nfb after 72 h of incubation. Thus, this indicates the capability of the endophytes to grow under microaerophillic conditions. However, based on the ARA results, only six strains (USML2, USML5, USML8, USML9, USML11 and USML15) were able to fix atmospheric N2 in a range of 2.2x10-10 to 8.5x10-7 μmol C2H4 cfu-1 h-1 within 24 h of incubation under microaerophilic conditions (Table 1).
| Table 1: | Acetylene Reduction Assay (ARA), indole-3-acetic acid (IAA) production and identification of locally-isolated endophytic diazotrophs from inner leaf tissues of oil palm |
 | |
Among the six strains, isolate USML5 exhibited the highest N2 fixation ability (8.5x10-7 μmol C2H4 cfu-1 h-1). These six endophytic diazotrophs were also capable in producing phytohormone in a range of 17.3 to 53.7 mM (Table 1). Based on partial 16S rDNA sequence analyses, the endophytic diazotrophs were closely related to Enterobacter sp. (USML2), Acinetobacter sp. (USML5), Azospirillum sp. (USML8), Phytobacter sp. (USML9), Azospirillum sp. (USML11) and Gluconacetobacter sp. (USML15) (Table 1).
Construction of GFP-tagged endophytic diazotrophs: In this study, all six endophytic diazotrophs (USML2, USML5, USML8, USML9, USML11 and USML15) were tagged with GFP via electroporation. This was done to study their ascending endophytic migration ability in the plant host. However, the GFP gene encoding GFP was successfully introduced only into Enterobacter sp. strain USML2. This was probably because the other strains were resistant to ampicillin which is already present in the pGFPCR. The cells of GFP-tagged-Enterobacter sp. strain USML2 exhibited strong green fluorescence, when observed under an epiflourescence microscope (Fig. 1a). Controls that were not exposed to the electric pulse did not produce any transformants.
Ascending endophytic migration of GFP-tagged-Enterobacter sp. Strain USML2: Behavior of endophytic bacteria that have potential use as inocula for plant promotion has been a major subject of this study. Since inocula are introduced on the soil or specifically on the roots, understanding the ascending endophytic migration from roots to leaves of the plant host (rice seedlings) is essential. In this study, GFP-tagged-Enterobacter sp. strain USML2 showed concentrated colonization initially at the root tips (Fig. 2c). Green fluorescent emission was the brightest at lateral root junctions of 7 day-old rice seedlings at 12 h after inoculation (Fig. 2c). The colonization also ascended into intracellular spaces of the stem base (Fig. 2b), though no colonization was observed in the leaf tip (Fig. 2a). At later incubation period (24 h), the endophytic diazotroph not only colonized into the whole root but also the stem base and leaf tip (Fig. 2d-g). The capability of Enterobacter sp. strain USML2 in ascending migration from root to leaf of the rice seedlings was probably due to its ability in producing the plant-cell wall degrading enzymes endopolygalacturonase (Fig. 1b) and endoglucanase (Fig. 1c). These enzymes play an important role in promoting colonization and ascending migration of endophytes from roots to leaves of the plant hosts.
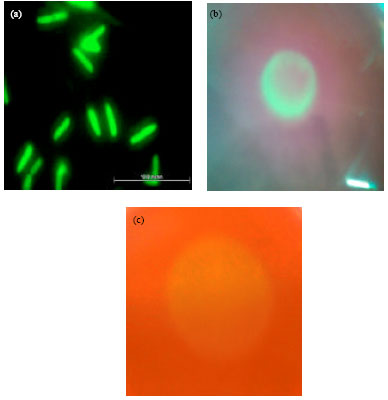 | |
| Fig. 1(a-c): | Enterobacter sp. strain USML2 (a) cells tagged with Green Fluorescent Protein (GFP) (b) production of endopolygalacturonase enzyme and (c) production of endoglucanase enzyme |
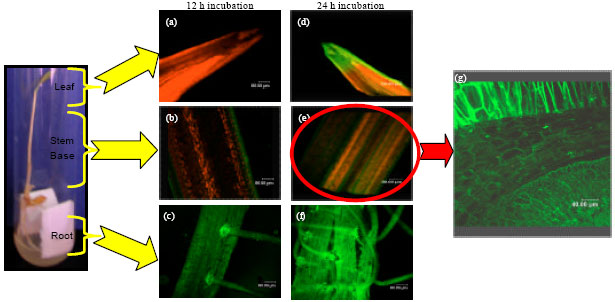 | |
| Fig. 2(a-g): | Fluorescent micrographs showing confocal laser scanning microscopy images of GFP-tagged-Enterobacter sp. strain USML2 in (a) Leaf at 12 h, (b) Stem base at 12 h, (c) Root at 12 h, (d) Leaf at 24 h, (e) Stem base at 24 h, (f) Root at 24 h and (g) Shows colonization ascended into intracellular spaces of stem base at 24 h |
DISCUSSION
Isolation and Characterization of Endophytic Diazotrophs: Endophytic bacteria often originate from the soil (Prakamhang et al., 2009). They live inside the plant host for at least part of their life cycle (Hardoim et al., 2008). In this study, 30 strains of endophytic bacteria were successfully purified due to the fact that the bacteria were isolated from the interior surface-sterilized leaf tissues of oil palm. Moreover, it is a challenge to isolate endophytic bacteria from inner leaf tissues because it is believed that the endophytic population in the leaf tissues is much less compared to in the roots (Prakamhang et al., 2009). Furthermore, it is not surprising that endophytic bacteria can survive inside plant tissues where O2 is limited (Minamisawa et al., 2004). Among the 30 isolated endophytes, only six strains were proven as diazotrophic bacteria. This was based on their capability to fix atmospheric N2. Earlier studies reported that N2 fixation activity was also detected in leaf consortia and indicated the ability of endophytic diazotrophs to fix N2 in planta (Prakamhang et al., 2009). These endophytic diazotrophs were able to synthesize phytohormone such as indole-3-acetic acid (IAA) which is a type of plant hormone that can affect plant growth promotion. In addition, phytohormone production not only plays an important role in plant development but also in activation of the plant defense system and functions as a signaling molecule in bacterial communication (Hardoim et al., 2008).
Ascending Migration of Endophytic Diazotroph: Microbial survivability inside plants could be explained based on the ability of diverse groups of endophytes to enter and persist in plants. These endophytes could infect the host plant via wounds caused by microbial or nematode phytopathogens or the stomata found in the leaf tissues. However, infection via root crack entry between displaced epidermal cells at the lateral root junctions is recognized as the main hot spot for bacterial colonization (Hardoim et al., 2008). Thus, this explains the concentrated colonization of GFP-tagged-Enterobacter sp. strain USML2 at the lateral root junctions of the 7 day-old rice seedlings at 12 and 24 h of inoculation. Invasion through these portals of entry will then quickly spread into the intercellular spaces in the root (Feng et al., 2005). Similarly, in this study, colonization also ascended into intracellular spaces of the stem base at 12 h of inoculation, followed by invasion of the whole root, stem base and leaf tip within 24 h of inoculation. The migration probably started with root surface colonization stage, followed by bacterial penetration into the cortical cells and spread to other tissues through xylem and stem.
Feng et al. (2005) observed that even though Azospirillum, Azotobacter and Paenibacillus are rhizosphere and rhizoplane (root surface) bacteria, they are also expected to be isolated from leaves since these bacteria can migrate from roots to shoots via internal tissue colonization. In addition, Roncato-Maccari et al. (2003) also reported the occurrence of rhizosphere and rhizoplane bacteria in leaf tissues of graminaceous plants. Bacterial colonization in healthy plant tissues like leaves, stem base and roots is a normal phenomenon and even the non-endophytic bacteria have been isolated from these plant parts (Lynch, 1990). This is probably because the microbes tend to colonize the plant region that is high in carbon (carbohydrate) concentration such as leaf tissues. Other bacterial strains which have been isolated from leaf tissues include Herbaspirillum and Gluconacetobacter (from sugar cane) and Beijerinkia (from coffee and cacao) (Moutia et al., 2003). The capability of these endophytic bacteria to colonize the interior tissues of their host plants is most probably due the production of plant-cell wall degrading enzymes which are endoglucanase and endopolygalacturonase. These enzymes are known to be responsible in plant invasion via degradation of plant-cell envelope and assist in efficient migration and colonization of the intercellular and intracellular spaces in plant tissues (Bacilico-Jimenz et al., 2001). Since Enterobacter sp. strain USML2 used in this study also produced endoglucanase and endopolygalacturonase enzymes, this thus explains its ability of ascending migration from root to leaf of the host plant. These enzymes are crucial for a successful endophytic colonization and invasion of plant tissues via degradation of the plant-cell wall (Elbeltagy et al., 2001; Reinhold-Hurek et al., 2006).
CONCLUSIONS
This study demonstrated the occurrence of culturable endophytes in leaf tissues of oil palm. The endophytes were classified as high-potential plant growth enhancers for non-leguminous host plants through the ability to fix nitrogen and produce phytohormone such as IAA. In addition, besides the ability of the endophyte (Enterobacter sp. strain USML2) to survive in the intercellular spaces, it is also capable in migrating upwards inside the plant from roots to leaves within 24 h. This was probably due to the plant-cell wall degrading enzymes (endoglucanase and endopolygalacturonase) that were crucial for efficient endophytic invasion and colonization of the plant. More research on the specific pathways and biochemical interactions involved in endophytic migration would be beneficial in understanding this important process.
ACKNOWLEDGMENTS
The authors are indebted to the School of Biological Sciences and Institute of Postgraduate Studies, Universiti Sains Malaysia (USM) for fellowship award; the School of Dental Sciences, USM Health Campus, Kelantan for assistance with confocal laser scanning microscopy (CLSM); the Malaysian Ministry of Science, Technology and Innovation (MOSTI) for research funding and Dr. Samuel C. Allen for a critical review of this manuscript.
REFERENCES
- Al-Falih, M.K.A., 2002. Factors affecting the efficiency of symbiotic nitrogen fixation by rhizobium. Pak. J. Biol. Sci., 5: 1277-1293.
CrossRefDirect Link - Alam, R., M.Z. Karim, M.F. Hasan, M.A. Hossain and A.B.M.M. Rahman, 2003. Evaluation of sugarcane clones with biological nitrogen fixation endowment. Asian J. Plant Sci., 2: 216-219.
CrossRefDirect Link - Aslin, C.O., H.G. Amir and L.K. Chan, 2005. Isolation and characterization of diazotrophic rhizobacteria of oil palm roots. Malay. J. Microbiol., 1: 31-35.
Direct Link - Bacilico-Jimenz, M., S. Aguilar-Flores, M.V. del Valle, A. Perez, A. Zepeda and E. Zenteno, 2001. Endophytic bacteria in rice seeds inhibit early colonization of roots by Azospirillum brasilense. Soil Biol. Biochem., 33: 167-172.
CrossRef - Barraquio, W.L., L. Revilla and J.K. Ladha, 1997. Isolation of endophytic diazotrophic bacteria from Wetland rice. Plant Soil, 194: 15-24.
CrossRefDirect Link - Bavykin, D.V., V.N. Parmon, A.A. Lapkin and F.C. Walsh, 2004. The effect of hydrothermal conditions on the mesoporous structure of TiO2 nanotubes. J. Mater. Chem., 14: 3370-3377.
CrossRef - Chi, F., S.H. Shen, H.P. Cheng, J.X. Jing, Y.G. Yanni and F.B. Dazzo, 2005. Ascending migration of endophytic rhizobia, from roots to leaves, inside rice plants and assessment of benefits to rice growth physiology. Applied Environ. Microbiol., 71: 7271-7278.
Direct Link - Choo, Q.C., M.R. Samian and N. Najimudin, 2003. Phylogeny and characterization of three nifH-homologous genes from Paenibacillus azotofixans. Appl. Environ. Microbiol., 69: 3658-3662.
CrossRef - Compant, S., B. Reiter, A. Sessitsch, J. Nowak, C. Clément and E.A. Barka, 2005. Endophytic colonization of Vitis vinifera L. by plant growth-promoting bacterium Burkholderia sp. strain PsJN. Appl. Environ. Microbiol., 71: 1685-1693.
CrossRefPubMedDirect Link - Elbeltagy, A., K. Nishioka, T. Sato, H. Suzuki and B. Ye et al., 2001. Endophytic colonization and in planta nitrogen fixation by a Herbaspirillum sp. isolated from wild rice species. Applied Environ. Microbiol., 67: 5285-5293.
Direct Link - Hardoim, P.R., L.S. van Overbeek and J.D. van Elsas, 2008. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol., 16: 463-471.
CrossRefPubMedDirect Link - Jolly, S.N., N.A. Shanta and Z.U.M. Khan, 2010. Phytoplankton quantification of heterotrophic bacteria and Azospirillum from the rhizosphere of taro (Colocasia esculenta L. Schott.) and the nitrogen fixing potential of isolated Azospirillum. Int. J. Botany, 6: 117-121.
CrossRef - Lynch, J.M., 1990. Beneficial interactions between micro-organisms and roots. Biotechnol. Adv., 8: 335-346.
CrossRef - Mia, M.A.B. and Z.H. Shamsuddin, 2010. Nitrogen fixation and transportation by rhizobacteria: A scenario of rice and banana. Int. J. Bot., 6: 235-242.
CrossRefDirect Link - Minamisawa, K., K. Nishioka, T. Miyaki, B. Ye and T. Miyamoto et al., 2004. Anaerobic nitrogen-fixing consortia consisting of clostridia isolated from gramineous plants. Appl. Environ. Microbiol., 70: 3096-3102.
CrossRef - Moutia, J.F.Y., G. Umrit, A.S. Saumtally and K.F. Ng Kee Kwong, 2003. The role of diazotrophic bacteria in the nitrogen nutrition of sugar cane in Mauritius: Preliminary results. AMAS Food and Agricultural Research Council, Reduit, pp: 29-39. http://www.gov.mu/portal/sites/ncb/moa/farc/amas2003/html/s1/s4.htm.
- Patten, C.L. and B.R. Glick, 2002. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Applied Environ. Microbiol., 68: 3795-3801.
CrossRefPubMedDirect Link - Pedraza, R.O., C.H. Bellone, S.C. de Bellone, P.M.F.B. Sorte and K.R.S. dos Teixeira, 2009. Azospirillum inoculation and nitrogen fertilization effect on grain yield and on the diversity of endophytic bacteria in the phyllosphere of rice rainfed crop. Eur. J. Soil Biol., 45: 36-43.
CrossRef - Prakamhang, J., K. Minamisawa, K. Teamtaisong, N. Boonkerd and N. Teaumroong, 2009. The communities of endophytic diazotrophic bacteria in cultivated rice (Oryza sativa L.). Appl. Soil Ecol., 42: 141-149.
Direct Link - Quispel, A., 1991. A critical evaluation of the prospects of nitrogen fixation with non-legumes. Plant Soil, 137: 1-11.
CrossRef - Reinhold-Hurek, B., T. Maes, S. Gemmer, M.V. Montagu and T. Hurek, 2006. An endoglucanase is involved in infection of rice roots by non-cellulose-metabolizing endophyte Azoarcus sp. strain BH72. Mol. Plant Microbe. Interact., 19: 181-188.
PubMed - Roncato-Maccari, L.D.B., H.J.O. Ramos, F.O. Pedrosa, Y. Alquini and L.S. Chubatsu et al., 2003. Endophytic Herbaspirillum seropedicae expresses nif genes in gramineous plants. FEMS Microbiol. Ecol., 45: 39-47.
Direct Link - Senthilkumar, M., M. Madhaiyan, S.P. Sundaram, H. Sangeetha and S. Kannaiyan, 2008. Induction of endophytic colonization in rice (Oryza sativa L.) tissue culture plants by Azorhizobium caulinodans. Biotechnol. Lett., 30: 1477-1487.
CrossRef - Singh, M.K., C. Kushwaha and R.K. Singh, 2009. Studies on endophytic colonization ability of two upland rice endophytes, Rhizobium sp. and Burkholderia sp., using green fluorescent protein reporter. Curr. Microbiol., 59: 240-243.
CrossRef - Sofi, P. and S. Wani, 2007. Prospects of nitrogen fixation in rice. Asian J. Plant Sci., 6: 203-213.
CrossRefDirect Link - Taghavi, S., C. Garafola, S. Monchy, L. Newman and A. Hoffman et al., 2009. Genome survey and characterization of endophytic bacteria exhibiting a beneficial effect on growth and development of poplar trees. Applied. Environ. Microbiol., 75: 748-757.
CrossRef