
Bidhan Roy
Department of Genetics and Plant Breeding, Regional Research Station, Terai Zone, Uttar Banga Krishi Viswavidyalaya, Pundibari, Cooch Behar 736 165, West Bengal, India
S. K. Noren
College of Post Graduate Studies, Central Agricultural University, Umiam 793 103, Meghalaya, India
Asit B. Mandal
Directorate of Seed Research, Khushmaur, P.O. Kathuli, Maunath Bhanjan 275 101, Uttar Pradesh, India
Asit K. Basu
Department of Seed Science and Technology, Bidhan Chandra Krishi Viswavidyalaya, Mohanpur, Nadia 741 252, West Bengal, India







ABSTRACT
Abiotic stresses have become an integral part of crop production. One or other persist either in soil or in atmosphere. With the ultimate goal to raise the crop plants with better suitability towards rapidly changing environmental inputs, intense efforts are needed employing physiological, biochemical and molecular tools to improve tolerance ability under abiotic stresses. Attempts have been taken by plant breeders to develop tolerant varieties of different crops for specific abiotic stress. Appreciable improvement also has been done by the molecular biologists regarding to perturbations in gene expression and protein during stress. Employing transgenic technology, functional validation of various target genes involve in diverse processes, such as signaling, transcription, ion homeostasis, antioxidant defense etc. for enhanced abiotic stress tolerance has been attempted in various model system and some of them have been extended to crop plants. The information in the areas of gene and genetic engineering for improvement of crop plants against abiotic stresses are lying unorganized in different articles of journals and edited books. This information has been compiled in this review article in organized way with up-to-date citations, which will provide comprehensive literatures of recent advances.
PDF Abstract XML References Citation
Received: September 02, 2010;
Accepted: September 30, 2010;
Published: February 22, 2011
How to cite this article
Bidhan Roy, S. K. Noren, Asit B. Mandal and Asit K. Basu, 2011. Genetic Engineering for Abiotic Stress Tolerance in Agricultural Crops. Biotechnology, 10: 1-22.
DOI: 10.3923/biotech.2011.1.22
URL: https://scialert.net/abstract/?doi=biotech.2011.1.22
DOI: 10.3923/biotech.2011.1.22
URL: https://scialert.net/abstract/?doi=biotech.2011.1.22
INTRODUCTION
Most crops growing under field conditions are often being exposed to various abiotic stresses. The complex field environment with its heterogenic conditions and global climate change are increasing day by day but a few of them challenges facing modern agriculture (Mittler and Blumwald, 2010). A combination of plant breeding approaches will likely be needed to improve significantly the abiotic stress tolerance of crops in the field. A number of abnormal environmental parameters, such as drought, salinity, cold, freezing, high temperature, waterlogging, high light intensity, UV-radiation, nutrient imbalances, metal toxicities, nutrient deficiencies, climate change etc. are collectively termed as abiotic stress. Only 10% of world's arable land may be categorized as free from stress. The rapid change in environmental conditions are likely to override the adaptive potential of plants, these environmental changes mainly originated from anthropogenic activities causing soil and air pollution, thus plants are exposed to natural climatic or edaphic stresses. Among abiotic stresses drought is the main abiotic factor as it affects 26% of arable area. Water stress is a single most severe, limitation to the productivity of rice in the rainfed ecosystem (Windawsky and O’Toole, 1990). Mineral toxicity/deficiencies are second in importance. Among mineral toxicity, salinity is wide spread and is estimated to affect 10% of the world land surface (Richards, 1995). Increasing salinization of arable land is expected to have devastating global effects, resulting in 30% land losses within the next 25 years and up to 50% by the year 2050 (Wang et al., 2003a). Soil acidity is another major problem common to tropical regions, which constitutes about 3.95 billion ha of land (FAO, 1991). Acid soils caused by combination of aluminium (Al) and manganese (Mn) toxicity are major constraints to soil fertility and crop productivity. Al toxicity problems are of enormous importance in production of rice, maize and sorghum. Low temperature stress accounts about 15% crop area worldwide and such stress mainly prevails in temperate zone.
There is serious concern for food security of developing countries, which demands a conscious effort to improve production from areas commonly exposed to abiotic stresses. As mindless urbanization and industrialization swallow fertile lands and overuse of pesticides puts the environment jeopardy, researchers are designing crops that could tolerate abiotic stresses. The limitations to increase crop yield on existing cropped areas can be partially overcome with greater production inputs manipulation, which are costly, laborious and beyond the reach of small and marginal farmers. Thus, there is an urgent need to develop varieties that can not only withstand high levels of abiotic stresses but can also maintain optimum yield levels. In conventional breeding programme, genetic markers are being used by plant breeders, which are of morphological traits and controlled by single locus. The morphological markers are not always useful for selection for abiotic stresses. Therefore, molecular breeding (biochemical and/or DNA markers) is advancing as a new chapter for quick improvement in crops suitable for problem soils. Molecular control mechanisms for abiotic stress tolerance are based on the activation and regulation of specific stress-related genes. These genes are involved in the whole sequence of stress response, such as signaling, transcriptional control, protection of membranes and proteins and free-radical and toxic compound scavenging. The complex plant response to abiotic stress, which involves many genes and biochemical-molecular mechanisms, is schematically represented in Fig. 1. Furthermore, reproductive barriers limit transfer of favourable alleles from interspecific and intergeneric sources. Transgenic development is another straight forward technology to improve crop yield in abiotic stress affected land (Roy and Basu, 2009). The development of tolerant crops by genetic engineering, on the other hand, requires the identification of key genetic determinants underlying stress tolerance in plants and introducing these genes into crops. Introduction of molecular change by genetic engineering takes less time compared to plant breeding methods: only desired gene(s) can be transferred, whereas, in conventional breeding approach is associated with simultaneous transfer of undesired gene(s). For re-cultivation of degraded soils and reclamation of industrial sites, stress tolerant plants are required. The advent of plant transformation may have placed within the grasp of the possibility of engineering greater abiotic stress tolerance in plants.
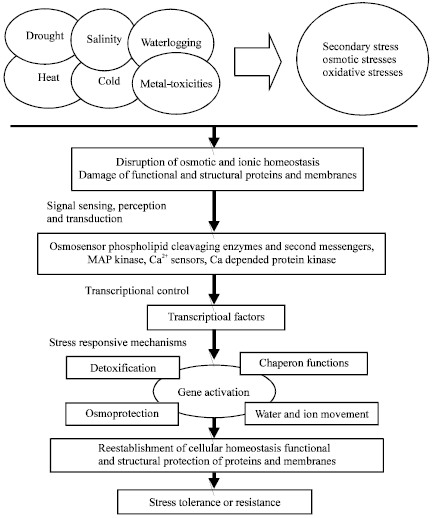 | |
| Fig. 1: | Plant response to abiotic stress to develop tolerance or resistance |
SALT TOLERANCE
Salt tolerance is an important trait that requires overcoming salinity induced reduction in plant productivity. The genetic response of plants to abiotic stresses is complex involving simultaneous expression of a number of genes. Plant genetic engineering techniques could be effectively utilized to exploit some of the untapped potentials to increase the harvestable crop yield. It involves specific gene manipulation either through over expression or silencing of alien/native genes. A number of genes induced in response to salinity have been identified from a range of organisms adapted to stressful environment. If a salt tolerant gene is identified which can lead to betterment of the crops, it is possible to transfer that progress in transgenic research for inclusion salinity stress tolerance, which has been presented in Table 1.
Transgenic research has made significant progress in crop genetic improvement with the advent of modern rDNA technologies. A large number of transgenics in diverse crops are on large-scale cultivation. Moreover, multiple genes can be stacked or transformed to a stock of interest through genetic transformation. There are a large number of genes found to be instrumental and there are many functional targets for engineering tolerance to salinity. Few of the genes of importance are briefed below.
Ion transporter and Antiporter genes: A salt concentration of 200 mM is equivalent to 40% of the salt concentration of sea water and will inhibit growth of almost all crop plants.
| Table 1: | List of transgenic research for salt stress tolerance |
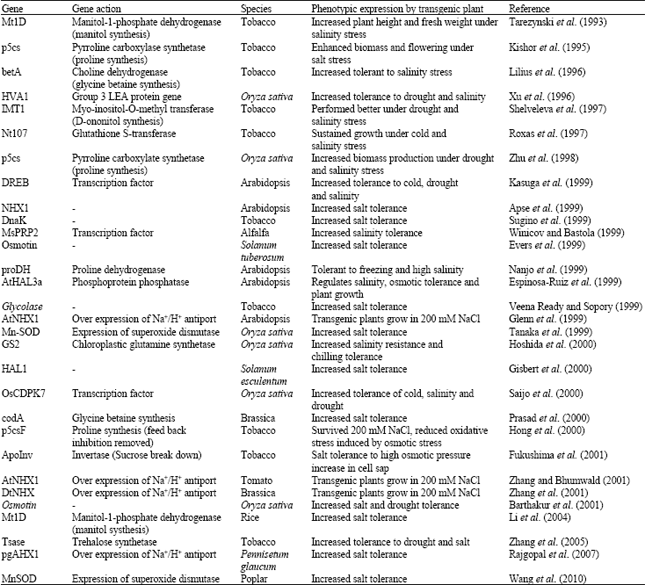 | |
Ion trans porters selectively transport ions and maintain them at physiologically relevant concentrations while Na+/H+ antiporters also play a crucial role in maintaining cellular ion homeostasis, thus permitting plant survival and growth under saline conditions. The Na+/H+ antiporters catalyze the exchange of Na+ for cytoplasmic pH, sodium levels and cell turgour (Serrano et al., 1998).
A construct containing the DtNHX gene, coding for a vacuolar Na+/H+ antiport from Arabidopsis thaliana, was introduced into the genome of Brassica napus cv. Westar by Zhang et al. (2001). Over expression of the vacuolar Na+/H+ antiport did not affect the growth of transgenic plants since similar growth was observed when wild type and transgenic plants were grown in the presence of 10 mM NaCl. While growth of wild type plants was severely affected by the presence of 200 mM NaCl in the growth solution, transgenic plants grew, flowered and produced seeds. They noted that the transgenic plants grown at 200 mM NaCl produced number of seeds similar to those of wild-type plants grown at low salinity. Moreover, qualitative and quantitative analysis of oil content showed no significant differences between seeds from wild type plants grown at low salinity and transgenic plants grown at high salinity. They also observed that the transgenic plants accumulated up to 6% Na without altering the yield and oil content. Transgenic Brassica juncia plants over expressing pgNHX1 withstand 300 mM salt stress till the seed setting stage and exhibited normal growth phenotype without much loss in seed yield (Rajgopal et al., 2007).
Glenn et al. (1999) have engineered transgenic Arabidopsis plants that overexpress AtNHX1, a vacuolar Na+/H+ antiport, which allowed the plants to grow in 200 mM NaCl. Zhang and Bhumwald (2001) reported the genetic modification of tomato plants to overexpress the Arabidopsis thaliana AtNHX1 antiport, which likewise allowed those plants to grow in the presence of 200 mM NaCl. Besides providing farmers with a cash crop for salted lands, such, plants may also pull salt out of soils, enabling other crops to thrive again. Wheat productivity is severely affected by soil salinity due to Na+ toxicity to plant cells. Xue et al. (2004) generated transgenic wheat expressing a vacuolar Na+/H+ antiport gene AtNHX1. The transgenic wheat lines exhibited improved biomass production. The field trial revealed that the transgenic wheat lines produced higher grain yield and heavier and larger grains in the field of saline soil. The transgenic rice accumulated a lower level of Na+ and higher level of K+ in the leaves than the non-transgenic plants under saline environment.
The Escherchia coli nahA gene encodes a Na+/H+ antiporter, which plays critical role in ion homeostasis has been transferred into rice (Oryza sativa L. sp. Japonica) by Wu et al. (2005). The transgenic plants showed better germination rate, growth and average yield per plant than control. They also reported higher sodium and proline content in transgenic lines, implying that nhaA over-expression enhance osmoregulation by activating the bio-synthesis of proline.
Compatible solutes: Compatible solutes accumulate in organisms in response of osmotic stress. The primary function of compatible solute is to maintain cell turgour and thus the driving gradient for water uptake. Studies also indicated that compatible solutes can act as free radical scavengers or chemical chaperon by directly stabilizing membrane and/or proteins (Diamant et al., 2001). The Δ'adi (Choline oxidase) gene isolated from the soil bacterium Arthrobacter globiformis converts choline to glycine betaine via betain aldehyde (Deshnium et al., 1995). This gene encodes choline oxidase, the enzyme that converts choline to glycinebetaine. Transgenic potato plants expressing the oxalate oxidase enzyme were produced by Turhan (2005) using Agrobacterium-mediated genetic transformation. His findings revealed a relatively higher salt tolerance ability of transgenic than the non-transgenic genotypes in vitro. However, the glasshouse results were less consistent, but some transgenic genotypes showed superior yield characteristics to the non-transgenic under salinity.
Glycinebetaine is an extremely efficient osmoprotectant widely distributed among plants. It is a quaternary ammonium compound belonging to the organic solutes, which accumulate in higher plants in response to osmotic stress induced by drought, high salinity or low temperature (McNeil et al., 2000). The ability to synthesize and accumulate glycinebetaine contributes to salt and drought tolerance (Grumet and Hanson, 1986). Transformation of Arabidopsis thaliana with Δ'adi gene resulted in enhanced salt tolerance of transgenic Arabidopsis plants (Hayashi et al., 1997). Thus synthesis of glycinebetaine in transgenic plants in vitro, as a result of the expression of Δ'adi gene, might be very useful in improving the ability of crop plants to tolerate salt stress (Hayashi et al., 2001). Sakamoto et al. (1998) reported transgenic rice expressing the Δ'adi gene in the chloroplast and the cytosol recovered to normal growth at a faster rate than the wild type after an initial growth inhibition under salt and low temperature stress.Lilius et al. (1996) transformed tobacco plants with Δ'adi gene, which encodes for choline dehydrogenase protein (responsible for conversion of choline to betaine aldehyde) from E. coli. The Δ'adi gene introduction rendered the transgenic tobacco plants tolerant to high concentration of salt. Betaine aldehyde dehydrogenease (BADH) is the second enzyme in this pathway responsible for conversion of betaine aldehyde to glycinebetaine.
Tobacco has been transformed with the Δ'adi and betB genes, involved in the glycinebetaine (betaine) pathway of Escherichia coli (Holmstrom, 1998). The transgenic plants produced and accumulated osmolyte and exhibited enhanced stress tolerance as shown by enhanced growth under salt stress and improved recovery of PS-II after salt stress. High tolerance to NaCl was also obtained transferring bacterial beta gene to cabbage (Brassica oleracea var. capitata) cultivar Golden Acre through Agrobacterium-mediated genetic transformation of hypocotyl explants (Bhattacharya et al., 2004).
Manitol gene: Osmotically shocked cells synthesize and accumulate massive amount of osmoprotectory compounds. Such compounds possibly help the cells to lower their osmotic potential and to draw water from the outside medium. Manitol as an osmoprotectory compound is primarily found in microbes. By introducing manitol-1-phosphate dehydrogenease gene (mt1D) isolated from E. coli (Tarezynski et al., 1993) showed over-expression of manitol in tobacco plants. These transgenic plants showed tolerance to high NaCl levels (250 mM). Seeds of transgenic Arabidopsis transformed with mt1D gene under control of CaMV 35 promoter over produced manitol and germinated in a medium supplemented with high amount of NaCl (Thomas et al., 1995). Li et al. (2004) introduced mt1D gene into upland rice (Oryza sativa var. japonica) by microprojectile bombardment. Growth rate of transgenic plants was significantly higher than the control on MS medium containing 1% NaCl. Non-transgenic plants died after 35 days. They reported less membrane damage and low Na+/K+ ratio than the control under salt stress.
LEA protein: Osmotic stresses induce Late-embryogenesis-abundant (LEA) proteins in vegetative tissues of plants. The functions of LEA proteins are largely unknown. Nevertheless, their considerable synthesis during the late embryogenesis, their induction by stress and their structural characteristics permit the prediction of some of their functions. It has been suggested that LEA type proteins act as water binding molecules, in ion sequestration and membrane stabilization. LEA-proteins are encoded by Responsive to Dehydration (RD), Early Responsive to Dehydration (ERD), cold inducible (KIN), cold regulated (COR) and responsive to ABA (RAB) genes in different plant species. Xu et al. (1996) found that the hva1 gene, which encodes for a specific class of LEA proteins, when overexpressed in rice leads to increased salt tolerance. This study demonstrated that subcellular compartmentalization of the biosynthesis of glycinebetaine was a critical step in attaining enhancement of tolerance for salinity and water stress. Tansformation of Chinese cabagge (Brassica campestris ssp. pekinensis) by overexpression a B. napus Group 3 LEA gene enhanced tolerance to salinity and drought (Park et al., 2005).
Trehalose: Trehalose is a non-reducing disaccharide of glucose that functions as a protectant in the stabilization of biological structure and enhances the tolerance of organisms to abiotic stress. Zhang et al. (2005) transformed tobacco plants with trehalose synthetase (Tsase) gene for manipulating abiotic stress tolerance. They reported higher trehalose accumulation in transgenic plants as compared to non-transgenics. The finding suggested that the transgenic plants transformed with Tsase gene can accumulate higher levels of trehalose and have enhanced tolerance to drought and salt stresses.
H+-pyrophosphatase (H+-Ppase) gene: An H+-Ppase gene named TsVP involved in basic biochemical and physiological mechanisms was cloned from Thellungiella halophila. Transgenic tobacco overexpressing TsVP had 60% greater dry weight than wild-type tobacco at 300 mM NaCl (Gao et al., 2006). Their findings suggested that over expression of H+-Ppase causes the accumulation of Na+ in vacuoles instead of in the cytoplasm and avoids the toxicity of excess Na+ in plant cells.
WATERLOGGING TOLERANCE
The major changes in the waterlogged soils are physical, biological and chemical changes. Upon flooding, the pore spaces (filled with air) in the soil become saturated with water, as a result, the soil swells. Since, the exchange of air between the atmosphere and the soil is impeded and since the water particles are held by soil particles and prevent from percolating downward and escaping. The absence of soil air in waterlogged condition causes a change in the varieties of microbes, microscopic, organisms which live in the soil. Anaerobic microbes tend to be much slower; less efficient decomposer of organic matter than the aerobic microorganisms. Consequently, the rate of decay of organic matter tends to be slow in flooded soils.
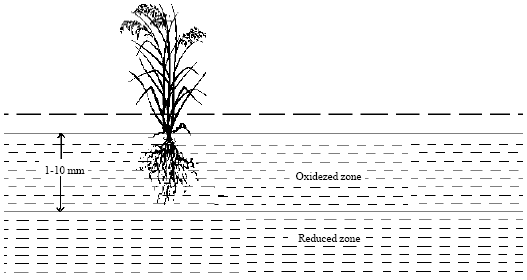 | |
| Fig. 2: | Chemical zones of flooded soil |
| Table 2: | Transgenics developed in different crop plants against waterlogging tolerance |
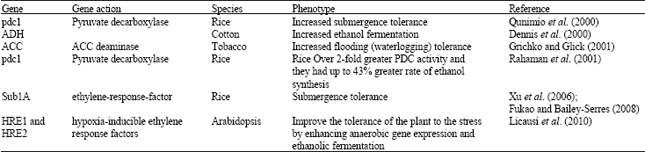 | |
Also the end products produced by anaerobic decomposition differ some are toxic to crop plants; particularly those released during the first two weeks after decomposition begins and the toxicity produced during decomposition may stunt the growth of crop plants. Flooded soils develop two distinct chemical zones (Fig. 2); oxidized and reduced zones. The upper oxidized zone (1-10 mm) absorbs oxygen from the water, turns brown in colour and reacts to nitrogen like an unfolded soil. The lower reduced zone, which extends down as far as the water, is extremely low in available oxygen, turns dark blue or gray in colour and takes on chemical properties quite different from those of oxidized layer above.
Genetic engineering provides opportunities for germplasm improvement as well as evaluating the impact of different mechanisms of tolerance to waterlogging without confounding effects of complete changes in the background. Two different approaches have been used to try and identify limiting factors in the response to waterlogging. First is the under-expression of single candidate genes, e.g., for ethanol synthesis, using sense and anti-sense constructs. Second is the over-expression of transcription factors (Dennis et al., 2000). It was anticipated that both approaches may have a beneficial effect in switching on the longer term adaptation response to low oxygen stress. Few achievements in transgenic development have been presented in Table 2.
Preliminary results of rice transgenic containing the cotton adh cDNA suggested that the over expressing pdc do not show increased tolerance to submergence. Qunimio et al. (2000) found that Taipei 309 transferred with pdc1 linked to a constitutive 35S promoter had up to 3-fold higher PDC activities and ethanol synthesis rates when exposed to anoxia compared to non-transformed controls. They also reported that increasing ethanol production up to 6-fold in a range of transgenic lines exposed to anoxia was correlated with an 8-fold increase in percentage survival of lines during submergence under hypoxic conditions. In contrast, Rahaman et al. (2001) studied Taipei-309 transformed with pdc1 and found that two transgenic lines had over 2-fold greater PDC activity and they had up to 43% greater rate of ethanol synthesis, however, survival of seed lines exposed to anoxia was even less than that of non-transformed plants. Therefore, the results of rice transgenic need to be repeated for its potential extrapolation.
Transgenic cotton plants containing the ADH cDNA driven by a constitutive 35S promoter showed 10-30 fold increase in ADH activity and a significant increase in the rate of ethanol fermentation (Dennis et al., 2000). Cotton plants with rice pdc1 cDNA driven by a constitutive 35S promoter produce more pdc protein but had only marginal more PDC activity. They also observed that neither pdc or adh transgenic cotton plants, nor plants containing both constructs showed increased tolerance of hypoxia stress.
DROUGHT TOLERANCE
Water becomes an increasingly scarce and precious commodity. It is thus essential to improve water use efficiency in agriculture. The development of crop varieties with increase tolerance to drought, both by conventional breeding methods and genetic engineering is important to meet global food demand with less water. Conventional breeding and marker assisted selection are being used to develop varieties more tolerant to water stress. These methods are time taking and resource consuming and germplasm dependent. Genetic engineering is alternative because of its potential to improve abiotic stress tolerance more rapidly. The techniques for gene transformation of crop plants have been applied for identification of genes responsible for drought resistance and their transfer. Genes expressed during stress are anticipated to promote cellular tolerance to dehydration through protective functions in the cytoplasm, cell membrane, alternations of cellular water potential to promote water uptake, control of ion accumulation and further regulation of other gene(s). A comprehensive study on transgenic plants summarized and nature of tolerance has been listed in Table 3.
| Table 3: | Genes/transgenic plants for drought tolerance |
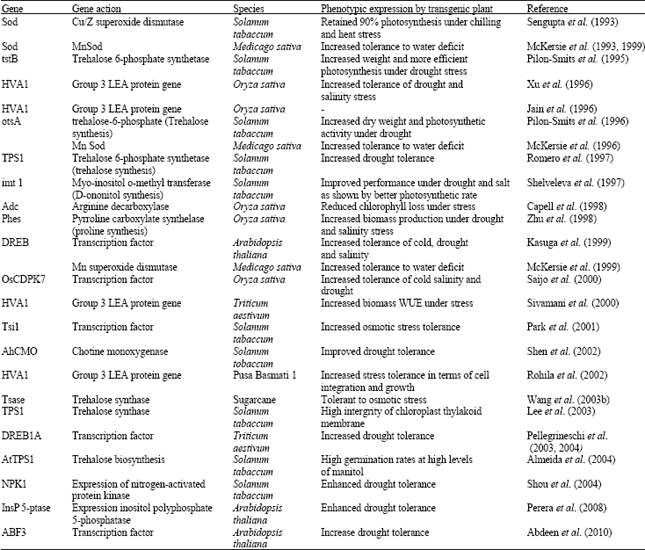 | |
Trihelose biosynthesis: Bacterial trehalose-6-phosphate (OtsB) genes introduced in tobacco showed better growth under drought stress (Pilon-Smits et al., 1995). Transgenic plants also showed a better capacity to retain water and preformed more efficient photosynthesis under stress. The gene TPS1 found in yeast encodes for trehalose-6-phosphate synthetase and is involved in biosynthesis of trehalose. The transgenic tobacco plants containing the yeast TPS1 gene exhibited multiple alteration and improved drought tolerance (Romero et al., 1997). By determining the effect of withholding irrigation on the death and damage of leaf (Romero et al., 1997), it has been shown that the transgenic plants have increased drought resistance. Further, Holmstrom (1998) transferred tobacco, resulting in the accumulation of trehalose and improved drought resistance. It is suggested that the improved drought tolerance is due to enhanced water retention capacity. Serrano et al. (1998) reported that yeast regulatory gene involved in stress tolerance, TPS1, have been expressed in transgenic plants. Almeida et al. (2004) transformed Nicotiana tabaccum with Arbidopsis thaliana gene (AtTPS1), which is involved in trehalose biosynthesis, by Agrobacterium-mediated genetic transformation. They observed high germination rates at higher levels of manitol than did wild type plants.
The trehalose synthetase gene (TSase ) gene from Grifola frondosa was transferred into sugarcane (Wang et al., 2003b). The high frequency phosphinothricine-resistant plants were obtained from the transformation. Some transgenic plants showed multiple phenotypic alterations and some plants showed improved tolerance to osmotic stress. Another trehalose phosphate synthetase (TPS1) gene from yeast was introduced into tobacco chloroplast (Lee et al., 2003). Stable integration of TPS1 into chloroplast genome of tobacco was confirmed by PCR and Southern blots analysis. Transgenic chloroplast thylakoid membranes showed high integrity under osmotic stress as evidenced by the retention of chlorophyll even when grown in 6% PEG6000, whereas, chloroplasts of untransformed plants were bleached. Thus, they suggested that trehalose functions by protecting biological membranes rather than regulating water potential.
LEA protein: Transgenic rice carrying barley HVA1 gene had shown drought resistance (Xu et al., 1996). Gene HVA1 encodes for a group of three LEA proteins which get accumulated in vegetative organs during drought condition. Transgenic rice showed enhanced accumulation of the HVA1 protein, which led to higher growth rates, delayed stress-related damage systems and improved recovery from the removal of stress conditions (Xu et al., 1996). The transgenic rice plants exhibited constitutive high expression of HVA1 protein ranging from 0.3-2.5 and 0.3-1.0% of the total soluble protein in leaf and root, respectively. Transgenic wheat plants containing the HVA1 gene showed consecutive expression of the transgene resulting in improvement of growth characteristics under water deficient conditions, more biomass and more efficient water use (Sivamani et al., 2000). Rohila et al. (2002) transformed Pusa Basmati 1 with HVA1 to increase tolerance against abiotic stresses. They developed transgenic lines which showed increased stress tolerance in terms of cell integrity and growth after imposed salt-and water stresses. Their findings exhibited high levels of LEA3 accumulation in the leaves of transgenic Pusa Basmati 1 rice plants might have conferred the significant increase in tolerance against drought and salt stresses.
Fructan synthesis: The bacterial gene sacB found in Bacillus subtilis encodes for levan sucrase, which takes part in fructan synthesis. Fructan promotes the process of root branching, thus increasing root surface and water uptake. Pilon-Smits et al. (1995) have shown that over-expression of scaB gene from Bacillus subtilis leads to high level of fructans in transgenic tobacco cells, leading in higher capacity for osmotic adjustment and this is associated with increased drought tolerance. The additional carbohydrate gained may lead to deeper rooting and greater water-uptake (Pilon-Smits et al. 1995; Blum et al., 1996; Schellenbaum et al., 1999). The transgenic tobacco that produces bacterial fructans was produced and examined for growth of the transgenic plants was significantly higher both on fresh weight and dry weight basis under drought stress compared to the wild type tobacco. The transgenic tobacco exhibited significantly more biomass accumulation in roots under drought stress and higher non-structural carbohydrate content under all conditions.
Proline accumulators: Proline accumulation has been demonstrated to be associated with abiotic stress (Delauney and Verma, 1993). Some genes related to proline synthesis, transport and accumulation have been identified (Delauney and Verma, 1993). Transgenic tobacco overexpressing P5CS gene transferred from mothbean exhibited a high level of enzyme and produced 10-18 fold more proline than control plant (Kishor et al., 1995). Over-production of proline enhanced root biomass and flower development under drought condition. Zhu et al. (1998) also found that transgenic rice with P5CS gene enhanced root biomass and flower development under water stress. Transgenic soybean plants transferred with antisense P5CR (L-Δ'-pyrroline-5-carboxylate reductase) gene showed increased proline accumulation, leading to higher tolerance to water stress (De Ronde et al., 2000).
Polyamine biosynthesis genes: In plants, polyamine, accumulate under several abiotic stress stimuli, including drought and salt. It has been suggested that this increase in polyamine concentration could be considered as an indicator of plant stress. With the availability of genes responsible for polyamine biosynthesis such as ADC (encodes for arginine decarboxylase), it is now possible to manipulate polyamine content using sense and antisense constructs of these genes in transgenic plants. Engineering of the plant polyamine biosynthesis pathway has concentrated mostly on two species, tobacco and rice (Kumar and Minocha, 1998; Capell and Christou, 2004). They suggested that further studies are required to understand the tolerance ability of these genes. The increase in putrescine levels in plants under stress might be the cause of stress-induced injury or alternatively a mean of protection against stress. Roy and Wu (2001) expressing oat adc cDNA in rice under control of an ABA-inducible promoter resulted in transgenic rice plants with increased biomass when grown under salt stress. Capell and Christou (2004) have generated a diverse rice germplasm with altered polyamine content. Transgenic rice plant expressing the SAMDC DNA accumulated spermidine and spermine in seed at 2-3 fold higher levels compared to wild type. In another set of experiment, they have obtained ten fold putrescine accumulation in transgenic rice plants laboring oat adc cDNA compared to wild type. Spermidine and spermine de-novo synthesis in transgenic plants under drought stress is corroborated by the activation of the rice SAMDC gene. Transcript levels for rice SAMDC reach their maximum levels at 6 days after stress induction. Such increase in the endogenous spermidine and spermine pools of transgenic plants not only regulates the putrescine response, but also exerts an anti-senescence effect at the whole plant level, resulting in phenotypically normal plants. Wild type plants, however, are not able to raise their spermidine and spermine levels after 6 days of drought stress and consequently exhibit the classical drought stress response (Capell and Christou, 2004).
Choline monooxygenase (CMO) catalyzes the committed step of glycinebetaine (GlyBet) biosynthesis in many flowering plants. Over-expression of AhCMO improved drought tolerance in transgenic tobacco when cultured in medium containing PEG6000 (Shen et al., 2002).
Dehydration responsive transcription factor: Plant genomes contain a large number of Transcription Factors (TFs). TFs are key regulatory proteins that enhance or repress the transcriptional rate of their target genes by binding to specific promoter regions upon activation or deactivation of upstream signaling cascade. Most of these transcriptional factors belong to a few large multigene families. Individual members of the same family often respond differently to various stress stimuli, on the other hand, some stress responsive may share the same transcriptional factor (Chen et al., 2002; Kreps et al., 2002). The Dehydration-responsive Element (DRE) and C-repeat binding factors (CBF) was identified as a cis-acting element regulating gene expression in response to dehydration (salt, drought and cold stresses) in Arabidopsis (Van Rensburg and Kruger, 1994). The dehydration-responsive transcription factors DREB and C-repeat binding factors (CBF) bind to DRE and CRT cis-acting elements that contain the same motif (CCGAC). Members of the CBF/DREB1 family, such as CBF1, CBF2 and CBF3 (or DREB1B, DREB1C and DREB1A, respectively) are themselves stress-inducible. DREB/CBF proteins are encoded by AP2/EREBP multigene families and mediate the transcription of several genes. DREB1A, a transcription factor that recognizes dehydration response elements, has been shown in Arbidopsis thaliana to play a crucial role in promoting the expression of drought tolerant genes (Pellegrineschi et al., 2003, 2004). They have transformed DREB1A gene into wheat. Plant expressing the gene demonstrated substantial resistant to water stress compared with the control under stress condition. Recently, overexpression of CBF4 from barley has been shown to confer salinity, drought and low temperature tolerance in transgenic rice (Oh et al., 2007).
The NAC gene family members encode one of the largest families of plant specific TFs and are expressed in various developmental stages, tissues and environmental factors. Hu et al. (2006) isolated and characterized NAC gene SNAC1. SNAC1 over-expressing transgenic rice plants showed significantly improved drought resistance under field conditions and strong tolerance to salt stress (Hu et al., 2006).
The phosphoinositide pathway and inositol-1,4,5-trisphosphate (InsP3) are implicated in plant responses to stress. To determine the downstream consequences of altered InsP3-mediated signaling, Perera et al. (2008) generated transgenic Arabidopsis thaliana plants expressing the mammalian type I inositol polyphosphate 5-phosphatase (InsP 5-ptase), which specifically hydrolyzes soluble inositol phosphates and terminates the signal. Rapid transient Ca2+ responses to a cold or salt stimulus were reduced by ∼30% in these transgenic plants. Drought stress studies revealed, surprisingly, that the InsP 5-ptase plants lost less water and exhibited increased drought tolerance. The onset of the drought stress was delayed in the transgenic plants and abscisic acid (ABA) levels increased less than in the wild-type plants. Stomatal bioassays showed that transgenic guard cells were less responsive to the inhibition of opening by ABA but showed an increased sensitivity to ABA-induced closure. Transcript profiling revealed that the drought-inducible ABA-independent transcription factor DREB2A and a subset of DREB2A-regulated genes were basally upregulated in the InsP 5-ptase plants, suggesting that InsP3 is a negative regulator of these DREB2A-regulated genes. These results indicated that the drought tolerance of the InsP 5-ptase plants is mediated in part via a DREB2A-dependent pathway and that constitutive dampening of the InsP3 signal reveals unanticipated interconnections between signaling pathways (Perera et al., 2008).
Super oxidedismutase: Within a cell, the SODs constitute the first line of the defense against ROS. It is found in all subcellular locations, like mitochondria, chloroplast microsomes, glyoxysomes, peroxysomes, apoplast and the cytosol. SODs are metaloproteins found in various compartments of plant cells and contain Cu and Zn, Fe or Mn cofactors. Transgenic tobacco plants containing oxidative stress- related genes showed elevated levels of glutathione reductase, superoxide dismutase and ascorbate peroxidase, resulting in enhance drought tolerance (Van Rensburg and Kruger, 1994).
Rice transgenic plants carrying Fe-SOD gene in Annada (Fig. 3a, b) was produced using Agrobacterium-mediated transformation system (unpluished data from thesis of Bidhan Roy). This result also confirmed successful Agrobacterium-mediated genetic transformation in rice and ample potential of this technique in genetic engineering for crop its improvement. Percent regeneration (%) of transformants as obtained in this study is found to be corroborative with the findings of Lin et al. (1995) and Datta et al. (1999, 2000).
Wax production: The gene designated WXP1, is able to activate wax production and confer drought tolerance in alfalfa (Medicago sativa). Overexpression of WXP1 under the control of CaMV355 promoter led to a significant increase in cuticular wax loading on leaves of transgenic alfalfa (Zhang et al., 2005). WXP1 over expression induced a number of wax-related genes.
 | |
| Fig. 3: | Regeneration and maintenance of transgenic plants under in vitro and in vivo containments. (A) Plants turned to be albino on selection medium fortified with 100 mg L-1of kanamaycin and 50 mg L-1 cefotaxime, B) Fertile transgenic plant of Annada on cement pots |
Transgenic leaves showed reduced water loss and chlorophyll leaching- transgenic alfalfa plants with increased cuticular wax showed enhanced drought tolerance demonstrated by delayed wilting after watering has been ceased and quicker and better recovery when the dehydrated plants were re-watered.
A full length rDNA of dehydrin BcDh2 from Boea crassifolia and its antisense nucleotide sequence was transferred into tobacco, var. NC89 under the control of CMV promoter (Ye et al., 2004). Under progressive water stress, the photosynthetic rate, transpiration rate and stomatal conductance of sense and antisense plants decreased. However, those parameters increased after 24 h of watering and the enhanced was higher in sense and antisense plants than the control.
Mitogen-activated protein: Expression of mitogen-activated protein kinase gene (MAPK) genes activates an oxidative signal cascade and lead to the tolerance of freezing, drought, heat and salinity stressing transgenic tobacco. MAPKs perform their function as a part of kinase modules, which is mainly composed of MAPKs, MAPKKs and MAPKKKs. Shou et al. (2004) transformed maize with a tobacco MAPKKK (NPK1). They found that the NPK1 expression enhanced drought tolerance in transgenic maize. Under drought stress, it maintained significantly higher photosynthesis rates than did the non-transgenic control, suggesting that protected photosynthesis machinery is protective from dehydration damage.
ABA Signaling: Protecting crop yield under drought stress is a major challenge for modern agriculture. One biotechnological target for improving plant drought tolerance is the genetic manipulation of the stress response to the hormone abscisic acid (ABA). In response of water stress, ABA levels in plant greatly increase resulting closure of stomata, thereby reducing the level of water loss through transpiration from leaves and activate response of genes. ERA1, a gene identified in Arabidopsis, encodes β-subunit of a farenesyl-transfarase and involves in ABA signaling. Wang et al. (2005) developed transgenic plants which performed significantly better under water stress, with consistently higher yield over conventional varieties of canola, whereas they observed no difference in performance of transgenic and control under sufficient water.
HEAT TOLERANCE
The successful development of transgenics have been presented in Table 4. The biochemical basis of heat tolerance was investigated by comparing the response of antisense and sense transgenic soybean plant containing L-Delta-1-pyrroline-5-carboxylate reductase gene (De Ronde et al., 2004) with non-transgenic wild-type plant. Upon stress, NADP+ levels decreased in sense plants. Sense plants had the highest ability to metabolize proline after rewatering. They reported dissociation of Oxygen-Evolving Complex (OEC) upon stress. In sense plants, which best resisted the stress, OEC dissociation was by passed by proline feeding electrons into PS II, maintaining an acceptable NADPH level, preventing further damage.
Heat shock proteins: It is important to maintain protein in their functional conformations and preventing aggregation of non-native proteins under stress. Many stress responsive proteins, particularly Heat Shock Proteins (HSPs) have been shown to act as molecular chaperones, which are responsible for protein synthesis, targeting, maturation and degradation in a broad array of normal cellular process. Furthermore, molecular chaperones function in the stabilization of proteins and membranes and in assisting protein folding under stress conditions.
Ahn and Zimmerman (2006) fused DcHSP17.7 gene (a carrot heat shock protein gene encoding HSP17.1) to a 6XHistidine (His) tag to distinguish the engineered protein from endogenous potato proteins and it was introduced into the potato cultivar Desiree under the control of the cauliflower mosaic virus (CaMV) 35S promoter. The integration was confirmed by Western Blot, which showed constitutive integration of DcHSP17.7 in transgenic potato lines before heat stress. They observed improved cellular membrane stability at high temperature, compared with wild type and vector controlled plants. Transgenic potato lines also exhibited enhanced tuberization in vitro.
COLD TOLERANCE
Classical plant breeding has limited success in imparting cold hardiness to crop plants. Biotechnology with its powerful tolls however may provide answer through the isolation of cold fighting genes and thus may help in the development of crop plants that can withstand freezing temperature. Some of the heat tolerant genes/transgenic plants have been listed in Table 5.
Several comprehensive genomic studies based on DNA micro-array performed in the field of plant cold acclimation, identified large number of cold responsive genes (Seki et al., 2001; Fowler and Thomashow, 2002; Kreps et al., 2002; Seki et al., 2002), such as COR, lipid transfer protein and β-amylase.
| Table 4: | Gene encoding for molecular chaperones and transgenic development |
 | |
| Table 5: | Cold tolerance gens/transgenic plants |
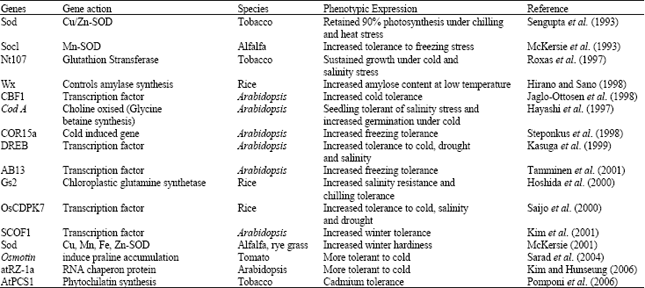 | |
A large number of these genes encode proteins with known enzyme activities, that potentioally contribute to freezing tolerance. Studies revealed that the cold acclimation is controlled by many genes and that cell membranes are particularly vulnerable to cold damage. Lee and Lee (2003) also identified many cold inducible alcohol dehydrogenase, β-amylase and many novel genes.
Dehydration responsive element: Many of the known cold regulated genes were under control of a primary master regulator, CBF/DREB1. Dehydration Response Element (DRE) plays an important role in the response to low temperature. The transcription factor DREB1A specifically interacts with DRE and induces the expression of stress tolerance genes in plant.
Over expression of DREB1A in transgenic Arabidopsis plants activated the expression of many of these stress tolerance genes and resulted in tolerance to freezing (Liu et al., 1998; Kasuga et al., 1999; Seki et al., 2001; Urao et al., 2000). Liu et al. (1998) and Jaglo-Ottosen et al. (1998) activated gene expression by overexpressing a homolog of CBF1, designated DREB1A. Their result indicated that the expression of the CRT/DRE regulation also increase drought tolerance. Liu et al. (1998) observed that the overexpression of DREB1A in transgenic Arabidosis resulted in dwarf phenotype. Fan et al. (2002) cloned an antifreeze gene from carrot and successfully transferred it into tobacco.
Cold regulated LEA protein: It was found that the COR15am the mature COR15a polypeptide, acts directly as cryoprotective protein by inhibiting the formation of hexagonal II phase lipid, a major type of freeze-induced membrane lession in non-acclimated plants (Steponkus et al., 1998). The COR15a gene enhanced the freezing tolerance of chloroplasts in engineered plants by almost 2C, which was nearly one third of the increase seen due to cold acclimation. While, this might not appear as large increase, an improvement of freeze tolerance by 2°C could potentially benefit certain crop plants (Artus et al., 1996). COR85, a group-II LEA protein was shown to be involved in cryoprotection of freezing-sensitive enzymes (Kazuaka and Oeda, 1994).
Overexpression of atRZ-1a did not affect the expression of various cold-responsive genes such as COR6.6, COR15a, COR47, RD29A, RD29B and LTI29 (Kim and Hunseung, 2006). Proteome analyses revealed that overexpression of atRZ-1a modulated the expression of several stress-responsive genes and the transcript levels and RNA stability of these target genes were not affected by atRZ-1a. atRZ-1a successfully complements the cold sensitivity of Escherichia coli lacking four cold shock proteins. These results strongly suggest that atRZ-1a plays a role as an RNA chaperone during the cold adaptation process.
Osmotin: Sarad et al. (2004) developed transgenic tomato with osmotin gene. Their preliminary tests revealed that the transgenic plants are more tolerant to cold than wild types.
Dehydrin: To elucidate the contribution of dehydrins (DHNs) to freezing stress tolerance, Puhakainen et al. (2004) developed transgenic Arabidopsis plants over-expressing multiple DHN genes. The transgenic plants exhibited lower LT50 values and improved survival when express to freezing stress compared to control plants. Thus they concluded that dehydrines contribute to freezing stress tolerance in plants and this could be partially due to dehydrins protective effect on membrane.
Compatible solutes: Parvanova et al. (2004) transformed tobacco to accumulate different compatible solutes (proline, fructans or glycine-betaine) in order to improve its tolerance to low temperature. The elevated parameters of transgenic plants successfully survived againsed freezing stress. Mukhoopadhyay et al. (2004) developed transgenic with OSISAP1 gene confering tolerance to cold, dehydration and salt stress. The gene OSISAP1 was isolated from rice encoding a zinc-finger protein induced under abiotic stresses. Hur et al. (2004) also developed transgenic rice with OsP5CS2 gene. This gene encodes for a protein that is highly homologous to δ-1-proline-5-caboxylate synthetase (P5CS), a proline biosynthesis enzyme. Their result indicated that the OsP5CS2 gene is necessary for plant tolerance to salt and cold tolerance.
Potato plants (Solanum tuberosum cv. Desiree) transformed with yeast invertase gene acquired a higher tolerance to cold temperature as compared to the control plants, apparently due to the changes in sugar ratio produced by the foreign invertase. The expression of Osmyb4 in Arabidopsis thaliana plants showed a significant increase cold and freezing tolerance, measured as membrane or photosystem-II stability and as a whole plant tolerance (Vannini et al., 2004). They demonstrated by transient expression of Myb4 transactivater the PAL2, ScD9, SAD and COR15a cold inducible promoters.
Heat shock factors: Various studies showed that plant heat shock proteins are not only express in response to heat shock, but also under water, salt, oxidative stress and at low temperature. Li et al. (2003) suggested that Hsf (heat shock factor) gene may play a pivotal role in heat-shock-induced chilling tolerance and constitutive expression of the transcriopton regulated gene in chilling sensitive crops may be useful in improving tolerance against chilling stress. They transferred Arabidopsis thaliana Hsf1b (AtHsf1a) gene into tomato. The transgenic tomato plants harbouring this gene showed increased chilling tolerance.
ALUMINIUM TOXICITY TOLERANCE
Optimum growth and productivity and even cultivation of most of the plants is severely restricted in soils with elevated levels of Aluminium (Al). It is the third most abundant element in the earth’s crust. Al toxicity mainly targets root apex, resulting in inhibited root growth and function (Roy and Mandal, 2005a). As a result, Al toxicity leads to severe impairment in acquisition of water and nutrients from the soil, which results in a significant reduction in crop yield on acid soils (Delhaize et al., 2007).
In plants with genetic resistance to Al toxicity, the Al-exclusion and uptake from root tips have been found to be correlated to their increased capacity to release organic acids such as citric acid, which chelates Al3+ outside the plasma membrane. There are two classes of physiological mechanisms that enable plants to withstand toxic levels of Al in acid soils: exclusion of Al from the root apex and true tolerance to Al in the root and shoot symplasma (Delhaize et al., 2007). Currently, Al-activated exudation of organic acid anions from root apices is the best documented and characterized plant Al tolerance mechanism (Delhaize et al., 2007; Kinraide et al., 2005).
The production of Al-tolerant transgenic plant varieties should be considered an important part of crop management strategies to increase agricultural production on acid soils and to protect forests around strongly acidified industrial regions. At molecular level, identification of tolerant genes to Al-toxicity remains scanty. The first two plant Al tolerant genes are TaALMT1 (Sasaki et al., 2004) from Triticum aestivum and SbMATE from sorghum and barley (Furukawa et al., 2007; Magalhaes et al., 2007). Expression of TaALMT1 transgene facilitated Al-activated malate efflux and increased Al tolerance in both tobacco suspension cells (Sasaki et al., 2004) and transgenic barley plants Delhaize et al., 2004). SbMATE underlies the AltSB locus of sorghum, encodes a plasma membrane-localized citrate transporter that belongs to the multi-drug and toxic compound extrusion family (Magalhaes et al., 2007). Magalhaes et al. (2007) reported that over-expression of this gene in transgenic Arbidopsis plants conferred a significant increase in Al tolerance and root citrate exudation. Liu et al. (2009) have utilized an integrated genetical and physiological analysis to identify an member of the ArbidopsisMATE family, AtMATE as the functional homolog of SbMATE. AtMATE is expressed primarily in root and is induced by Al.
Development of transgenic tobacco plants, tolerant to Al-toxicity was reported by Pan et al. (1994). Mexican scientist also have developed transgenic rice conferring tolerance to high concentration of Al (Conway and Toenniessen, 1999), which could have immense prospects in genetic management of Al-toxicity. Some of other works on Al-tolerant transgenic development have been cited in Table 6.
| Table 6: | Gene/transgenic plants with gene conferring metal toxicity tolerance |
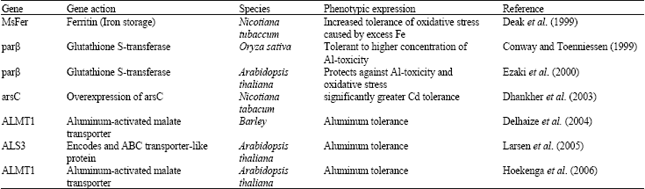 | |
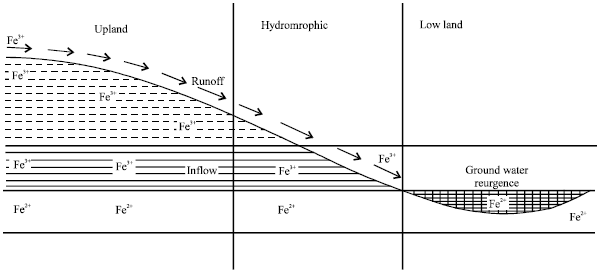 | |
| Fig. 4: | Cross section of toposequence showing movement of iron and reducing process |
IRON TOXICITY TOLERANCE
Fe-toxicity had been reported as one of the major soil constraints of lowland acid soils, inland valley swamps, costal swamps and irrigated lowlands in utisols and oxisols. Fe is also abundantly found in heavy soils (Tanaka and Yamaguchi, 1973). In the humid forest and moist Savana zones of Africa interflow of ferrous ion occurs from upper slopes (Fig. 4). More than 50% lowland rice is being affected with Fe-toxicity in Siera Leone, Liberia, Guinea, Nigeria, Ivory Coast and Senegal. It was also reported in Sri Lanka, Vietnam, Malaysia, India (Kerala, Orissa, West Bengal and Andaman and Nicobar Islands), Brazil, Colombia, Madagascar and Indonesia (Kalimanta and Sumatra), Philippinese (Sahrawat and Singh, 1995). Young acid sulphate soil in the coastal areas also show rampant symptoms and low lying valleys of Andaman and Nicobar Islands constitute a vast area constrained with Fe-toxicity, which is seriously limiting rice production (Mandal and Roy, 2000, 2003; Roy and Mandal, 2005b). In the people's Republic of China gleyic paddy soils are estimated to cover 7.5-8.0 m ha, where ferrous iron toxicity in the soil reduce rice yield by 10-20% depending on the intensity of toxicity and tolerance of the rice cultivar (Li et al., 1991; Wan et al., 2003).
Transgenic development: The transgenic development study in case of Fe-toxicity tolerance is very scanty. Deak et al. (1999) developed transgenic rice with ferritin gene to enhance high iron storage and they suggested that the enhanced Fe storage ability can reduce reactive oxygen species. Ferritin gene has been also transferred into rice (Khalekuzzaman et al., 2006), which enhanced iron level in the endosperm.
CADMIUM TOXICITY TOLERANCE
Cadmium (Cd) is a strongly phytotoxic heavy metals in an increasing environmental problem worldwide. It is one of the most dangerous metal due to its high mobility and the small concentration at which its effects on the plants being to appear. It is released into the environment by the power stations, heating systems, metal-working industries or urban traffic. It is recognized as an extremely significant pollutant due to its high toxicity and large solubility in water (Pinto et al., 2004). Soil solutions which have a Cd concentration ranging from 0.32 to 1.00 mM, can be regarded as polluted to a moderate level (Sanita di Toppi and Garbrielli, 1999). Cd toxicity is highest in acidic environment and decrease as the soil pH is increased (Przybulewska, 2004). Availability of Cd to plants is regulated by pH, redox potential and other physicochemical parameters. The effect of Cd salt on the growth of seedlings was weaker in loamy soil and stronger in sandy soil. The root system was more sensitive to Cd ions than the cotyledons.
Transgenic development: Transgenic development is an alternative approach to improve cadmium (Cd) tolerance and its accumulating capacity in crop plants. There are few successful reports on genetically engineered crop plants tolerant to Cd-toxicity (Zhu et al., 1999; Creissen et al., 1999). Over expression of Escherichia coli gshI gene in Indian mustard (Brassica juncia) increased γ-glutamylcysteine synthetase (γ-ECS) activity leading increased Cd tolerance and accumulation. Transgenic tobacco (Nicotiana tabaccum, var Wisconsin 38) bearing the transgene coding for the polyhistidine cluster, combined with yeast metallothione (HiCUP) showed increasing resistance to the stress response induced by cadmium (Pavlicova et al., 2004). The Cd content of the aerial biomass of transgenic tobacco increased by 45-75% compared with control. Another transgenic tobacco developed by Antosiewicz and Henning (2004) carrying cDNA LTC1, a nonspecific transporter for Ca2+, Cd2+, Na+ and K+, which displayed substantially higher level of tolerance to Cd and accumulated less Cd in roots. Plants tolerate to heavy metals through sequestration with cysteine-rich peptides, phytochelatins. Cysteine is synthesized by cysteine synthase (CS, E.C. 4.2.99.8) from hydrogen sulfide and O-acetylsrine. Transgenic Nicotiana tabaccum var. Xanthi transformed with RCS1, a cytosolic cysteine synthase gene of rice showed higher cyteine synthase activity and Cd toxicity tolerance than the wild type (Harada et al., 2001). Their result suggested that introduction of a cysteine synthase gene into tobacco plants resulted in high level production of sulfur-containing compounds that detoxify the Cd.
Saccharomyces cerevisiae protein YCF1, a member of the ATP-binding cassette pumps Cd2+ conjugated to glutathione into vacuoles. Song et al. (2003) found improved Cd (II) resistance, enhanced Cd accumulation into vacuoles and elevated Cd content in transgenic plants, which are desirable characteristics for phytoremedition.
In nutshell, the load of abiotic stresses on crop production is being incremented gradually by the directed demands of human beings for their food and luxuries. Thus, abiotic stresses have become an integral part of crop production. The ground water is depleting fast due to both intensive and extensive cultivation during off-monsoon periods as well as supplementation of water through irrigation during monsoon. Poor quality irrigation water and depletion of ground water increased salinity in arid and semi-arid zones. Intensive cultivation and heavy feeder crops lead deficiency of some nutrients and toxicity of others. The industrial growth and increase in vehicles created heavy metal toxicity in many industrial areas. In combination of industrial growth and improvement of transportation systems facilitate aggressive air pollution, which subsequently forcing climate change around the globe. All of these exert greater influence on plant growth and crop productivity.
The productivity of major staple food crops have reached to a plateau. There is very little scope to increase crop production area too. Therefore, more emphasis is required to use problem soils effectively. The soil reclamation is a costly affair and it is temporary. Development of crop genotypes tolerant/resistant to the adverse conditions is the only solution of such problem. To develop tolerant/resistant plant genotypes, the plant breeder and plant biotechnologist should work together and they should have knowledge regarding the injury and tolerance mechanisms in plants for specific stress and plant symptoms to know the nature of abiotic stress, breeding methods and biotechnological approaches. Development of transgenic plants tolerant to specific abiotic stresses is the straight-forward solution to improve crop productivity under unfavourable environments. The promising transgenic can be maintained in vitro through repeated subculture (Roy and Mandal, 2006). Those transgenic also can be mass-multiplied using tissue culture technique (Roy and mandal, 2011). Many countries grow transgenic crops under unfavourable biotic and abiotic environments. Transgenic crops yet to be popularize worldwide for cultivation.
REFERENCES
- Abdeen, A., J. Schnell and B. Miki, 2010. Transcriptome analysis reveals absence of unintended effects in drought-tolerant transgenic plants overexpressing the transcription factor ABF3. BMC Genomics, 11: 69-89.
Direct Link - Ahn, Y.J. and L. Zimmerman, 2006. Introduction of carrot HSP17.7 into potato (Solanum tuberosum L.) enhances cellular membrane stability and tuberization in vitro. Plant Cell Environ., 29: 95-104.
PubMed - Almeida, A.M., S.S. de Araujo, L.A. Cardoso, M.P. Fevereiro, J.M. Tarne and D.M. dos Santos, 2004. Transformation of tobacco with an Arabidosis thaliana gene involved in trehalose biosynthesis as a model to increase drought resistance in crop plants. Procedings of the 17th EUCARPIA General Congress on Genetic Variation for Plant Breeding, Sep. 8-11, Tullen, Austria, pp: 285-289.
- Apse, M.P., G.S. Aharon, W.A. Snedden and E. Blumwald, 1999. Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science, 285: 1256-1258.
PubMed - Artus, N.N., M. Uemura, P.L. Steponkus, S.J. Gilmour, C.T. Lin and M.F. Thomashow, 1996. Constitutive expression the cold-regulated Arabidopsis thaliana COR15a gene effects both chloroplast freezing tolerance. Proc. Nat. Acad. Sci., 93: 13404-13409.
Direct Link - Barthakur, S., V. Babu and K.C. Bansal, 2001. Over-expression of osmotin induces proline accumulation Over-expression of osmotin induces in transgenic tobacco. J. Plant Biochem. Biotechnol., 10: 31-37.
Direct Link - Bhattacharya, R.C., M. Maheswari, V. Dineshkumar, P.B. Kirti, S.R. Bhat and V.L. Chopra, 2004. Transformation of Brassica oleracea var. capitata with bacterial betA gene enhances tolerance to salt stress. Sci. Hortic., 100: 215-227.
CrossRef - Capell, T., C. Escobar, H. Liu, D. Burtin, O. Lepri and P. Chistou, 1998. Overxpression of oat arginine decaboxylase cDNA in transgenic rice (Oryza sativa L.) affects normal development pattern in vitro and result in putrecine accumulation in transgenic plants. Theor. Applied Genet., 97: 246-254.
Direct Link - Chen, W., N.J. Provart, J. Glazebrook, F. Katagiri and H.S. Chang et al., 2002. Expression profile matrix of Arabidopsis transcription factor genes suggests their putative functions in response to environmental stress. Plant Cell, 14: 559-574.
PubMed - Creissen, G., J. Firmin, M. Fryer, B. Kular and N. Leyland et al., 1999. Elevated glutathione biosynthetic capacity in the chloroplasts of transgenic tobacco plants paradoxically causes increased oxidative stress. Plant Cell, 11: 1277-1292.
Direct Link - Datta, K., Z. Kouklikova-Nicola, N. Baisakh, N. Oliva and S.K. Datta, 2000. Agrobacterium-mediated engineering for sheath blight resistance of indica rice cultivar from different ecosystem. Theor. Applied Genet., 100: 832-839.
CrossRefDirect Link - Datta, K., R. Velazhahan, N. Oliva, I. Ona and T. Mew et al., 1999. Over expression of cloned rice thaumatin like protein (PR-5) in transgenic rice plant enhance environmental friendly resistance to Rhizoctonia solani causing sheath blight disease. Theor. Applied Genet., 98: 1138-1145.
Direct Link - De Ronde, J.A., W.A. Cress, G.H.J. Kruger, R.J. Strasser and J. van Staden, 2004. Photosynthetic response of transgenic soybean plants, containing an Arabidopsis P5CR gene, during heat and drought stress. J. Plant Physiol., 161: 1211-1224.
CrossRef - Deak, M., G.V. Horvath, S. Davletova, K. Torok and L. Sass et al., 1999. Plants ectopically expressing the iron-binding protein ferratin are tolerant to oxidative damage and pathogen. Nature Biotechnol., 17: 192-196.
Direct Link - Delauney, A.J. and D.P.S. Verma, 1993. Proline biosynthesis and osmoregulation in plants. Plant J., 4: 215-223.
CrossRef - Delhaize, E., B.D. Gruber and P.R. Ryan, 2007. The roles of organic anion permeases in aluminum resistance and mineral nutrition. FEBS Lett., 581: 2255-2262.
CrossRef - Delhaize, E., P.R. Ryan, D.M. Hebb, Y. Yamamoto, T. Sasaki and H. Matsumoto, 2004. Engineering high-level aluminum tolerance in barley with the ALMT1 gene. Proc. Natl. Acad. Sci., 101: 15249-15254.
Direct Link - Dennis, E.S., R. Dolferus, M. Ellis, M. Rahaman and Y. Wu et al., 2000. Molecular strategies for improving water logging tolerance in plants. J. Exp. Bot., 51: 89-97.
Direct Link - Dhankher, O.P., N.A. Shasti, B.P. Rosen, M. Fuhrmann and R.B. Meagher, 2003. Increased cadmium tolerance and accumulation by plants expressing bacterial arsenate reductase. New Phytol., 159: 431-441.
Direct Link - Diamant, S., N. Eliahu, D. Rosenthal and P. Goloubinoff, 2001. Chemical chaperones regulate molecular chaperones in vitro and in cells under combined salt and heat stresses. J. Biol. Chem., 276: 39586-39591.
CrossRef - Ezaki, B., R.C. Gardner, Y. Ezaki and H. Matsumoto, 2000. Expression of aluminium-induce gene in transgenicArabidopsis plants can ameliorate aluminium stress and/or oxidative stress. Plant Physiol., 122: 657-665.
Direct Link - Fan, Y., B. Liu, H. Wang, S. Wan and J. Wang, 2002. Cloning of an antifreeze protein gene from carrot and its influence on cold tolerance in transgenic tobacco plants. Plant Cell Rep., 21: 296-301.
Direct Link - Fowler, S. and M.E. Thomashow, 2002. Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway. Plant Cell, 14: 1675-1690.
Direct Link - Fukao, K. and J. Bailey-Serres, 2008. Submergence tolerance conferred by Sub1A is mediated by SLR1 and SLRL1 restriction of gibberellin responses in rice. Proc. Natl. Acad. Sci., 105: 16814-16819.
Direct Link - Fukushima, E., Y. Arata, T. Endo, U. Sonnewald and F. Sato, 2001. Improved salt tolerance of transgenic tobacco expressing apoplastic yeast-derived invertase. Plant Cell Physiol., 42: 245-249.
Direct Link - Gao, F., Q. Gao, X.G. Duan, G.D. Yue, A.F. Yang and J.R. Zhang, 2006. Cloning of an H+-Ppase gene from Thellungiella halophila and its heterologous expression to improve tobacco salt tolerance. J. Exp. Bot., 57: 3259-3270.
Direct Link - Gisbert, C., A.M. Rus, M.C. Bolarin, J.M. Lopez-Coronado and I. Arrillaga et al., 2000. The yeast HAL1 gene improves salt tolerance of transgenic tomato. Plant Physiol., 123: 393-402.
Direct Link - Glenn, E.P., J.J. Brown and E. Blumwald, 1999. Salt tolerant mechanisms and crop potential of halophytes. Crit. Rev. Plant Sci., 18: 227-255.
Direct Link - Grumet, R. and A.D. Hanson, 1986. Genetic evidence for an osmoregulatory function of glycinebetaine accumulation in barley. Austr. J. Plant Physiol., 13: 353-364.
Direct Link - Harada, E, Y.E. Choi, A. Tsuchisaka, H. Obata and H. Sano, 2001. Transgenic tobacco plants expressing a rice synthase gene are tolerant to toxic levels of cadmium. J. Plant Physiol., 158: 655-661.
Direct Link - Hoekenga, O.A., L.G. Maron, M.A. Pineros, G.M.A. Cancado and J. Shaff et al., 2006. AtALMT1, which encodes a malate transporter, is identified as one of several genes critical for aluminum tolerance in Arabidopsis. Proc. Natl. Acad. Sci., 103: 9738-9743.
Direct Link - Hirano, H.Y. and Y. Sano, 1998. Enhancement of Wx gene expression and the accumulation of amylose in response to cool temperatures during seed development in rice. Plant Cell Physiol., 39: 807-812.
CrossRefDirect Link - Holmstrom, K.O., 1998. Engineering plant adaptation to water stress. Acta Univ. Agric. Sueciae Agaria, 84: 1401-6249.
Direct Link - Hong, S.W. and E. Vierling, 2001. Hsp101 is necessary for heat tolerance but dispensible for development and germination in the absence of stress. Plant J., 27: 25-35.
PubMed - Hong, Z., K. Lakkineni, Z. Zhang and D.P.S. Verma, 2000. Removal of feedback inhibition of Δ1-pyrroline-5-carboxylate synthetase results in increased proline accumulation and protection of plants from osmotic stress. Plant Physiol., 122: 1129-1136.
Direct Link - Hoshida, H., Y. Tanaka, T. Hibino, Y. Hayashi, A. Tanaka and T. Takabe, 2000. Enhance tolerance to salt stress in transgenic rice over expression chloroplast glutamine synthetase. Plant Mol. Biol., 43: 103-111.
Direct Link - Hu, H., M. Dai, J. Yao, B. Xiao, X. Li, Q. Zhang and L. Xiong, 2006. Overexpressing a NAM, ATAF and CUC (NAC) transcription factor enhancing drought resistance and salt tolerance in rice. Proc. Nat. Acad. Sci., 103: 12987-12992.
Direct Link - Hur, J., K.H. Jung, C.H. Lee and G. An, 2004. Stress-inducible OsP5CS2 gene is essential for salt and cold tolerance in rice. Plant Sci., 167: 417-426.
Direct Link - Jaglo-Ottosen, K.R., S.J. Gilmour, D.G. Zarka, O. Schabenberger and M.F. Thomashow, 1998. Arabidopsis CBF1 overexpression induces COR genes and enhances freezing tolerance. Science, 280: 104-106.
Direct Link - Jain, R.K., S. Jain and R. Wu, 1996. Stimulatory effect of water stress on plant regeneration in aromatic Indica rice varieties. Plant Cell Rep., 15: 449-454.
CrossRef - Kasuga, M., Q. Liu, S. Miura, K. Yamaguchi-Shinozaki and K. Shinozaki, 1999. Improving plant drought, salt and freezing tolerance by gene transfer of signal stress-inducible transcription factor. Natl. Biotechnol., 17: 287-291.
PubMed - Kishor, P.B.K., Z. Hong, G.H. Miao, C.A.A. Hu and D.P.S. Verma, 1995. Overexpression of delta]-Pyrroline-5-Carboxylate Synthetas increases proline production and confers osmotolerance ion transgenic plants. Plant Physiol., 108: 1387-1394.
Direct Link - Kazuaka, T. and K. Oeda, 1994. Purification and characterization of Cor85-oligomeric complex from cold-acclimated spinach. Plant Cell Physiol., 35: 601-611.
Direct Link - Khalekuzzaman, M, K. Datta, N. Oliva, M.F. Alam, O.I. Jorder and S.K. Datta, 2006. Stable integration, expression and inheritance of ferritin gene in transgenic elite indica rice cultivar BR29 with enhanced iron level in the endosperm. Ind. J. Biotechnol., 5: 26-31.
Direct Link - Kim, J.C., S.H. Lee, Y.H. Cheng, C.M. Yoo and S.I. Lee et al., 2001. A novel cold-inducible zinc finger protein from soybean. SCOF-1, enhances cold tolerance in transgenic plants. Plant J., 25: 247-259.
PubMed - Kim, Y.O. and K. Hunseung, 2006. The role of a zinc finger-containing glycine-rich rna-binding protein during the cold adaptation process in Arabidopsis thaliana. Plant Cell Physiol., 47: 793-798.
Direct Link - Kinraide, T.B., D.R. Parker and R.W. Zobel, 2005. Organic acid secretion as a mechanism of aluminum resistance: A model incorporating the root cortex, epidermis and the external unstirred layer. J. Exp. Bot., 56: 1853-1865.
Direct Link - Kreps, J.A., Y. Wu, H.S. Chang, T. Zhu, X. Wang and J.F. Harper, 2002. Transcriptome changes for Arabidopsis in response to salt, osmotic and cold stress. Plant Physiol., 130: 2129-2141.
CrossRefDirect Link - Larsen, P.B., J.M. Geisler, C.A. Jones, K.M. Williams and J.D. Cance, 2005. ALS3 encodes a phloem-localized ABC transporter-like protein that is required for aluminum tolerance in Arabidopsis. Plant J., 41: 353-363.
PubMed - Lee, J.H. and F. Schoff, 1996. An Hsp70 antisense gene affects the expression of HSP70/HSc70, the regulation of HSF and the acquisition of thermotolerance in Arabidopsis thaliana. Mol. Gen. Genet., 252: 11-19.
PubMed - Lee, J.Y. and D.H. Lee, 2003. Use of serial analysis of gene expression technology to reveal changes in gene expression in Arabidopsis pollen undergoing cold stress. Plant Pysiol., 132: 517-529.
CrossRefDirect Link - Lee, S.B., H.B. Kwon, S.J. Kwon, S.C. Park, M.J. Jeong, S.E. Han, M.O. Byun and H. Daniell, 2003. Accumulation of trehalose within transgenic chloroplast confer drought tolerance. Mol. Breed., 11: 1-13.
Direct Link - Licausi, F., J.T. Van Dongen, B. Giuntoli, G. Novi, A. Santaniello, P. Geigenberger and P. Perata, 2010. HRE1 and HRE2, two hypoxia-inducible ethylene response factors, affect anaerobic responses in Arabidopsis thaliana. Plant J., 62: 302-315.
Direct Link - Lin, W., C.S. Anuratha, K. Datta, I. Potrykus, S. Muthukrishnan and S.K. Datta, 1995. Genetic engineering of rice for resistance to sheath blight. Nat. Biotechnol., 13: 686-691.
CrossRefDirect Link - Liu, J., J.V. Magalhaes, J. Shaff and L.V. Kochian, 2009. Aluminum-activated citrate and malate transporter from the MATE and ALMT families function independently to confer Arabidopsis aluminum tolerance. Plant J., 57: 389-399.
PubMed - Liu, Q., M. Kasuga, Y. Sakuma, H. Abe, S. Miura, K. Yamaguchi-Shinozaki and K. Shinozaki, 1998. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell, 10: 1391-1406.
Direct Link - Magalhaes, J.V., J. Liu, C. T. Guimaraes, U.G.P. Lana and V.M.C. Alves et al., 2007. A gene in the multidrug and toxic compound extrusion (MATE) familu confers aluminum tolerance in sorghum. Nature Genet., 39: 1156-1161.
Direct Link - Malik, M.K., J.P. Solvin, C.H. Hwang and J.L. Zimmerman, 1999. Modified expression of a carrot small heat shock protein gene, Hsp17.7, results in increased or decreased thermotolerance. Plant J., 20: 89-99.
PubMed - McKersie, B.D., S.R. Bowley, E. Harjanto and O. Leprince, 1996. Water-deficit tolerance and field performance of transgenic alfalfa over expressing superoxide dismutase. Plant Physiol., 111: 1177-1181.
PubMed - McKersie, B.D., S.R. Bowley and K.S. Jones, 1999. Winter survival of transgenic alfalfa overexpression superoxide dismutase. Plant Physiol., 199: 839-849.
Direct Link - McKersie, B.D., Y. Chen, M. de Bens, S.R. Bowley and C. Bowler et al., 1993. Superoxide dismutase enhances tolerance for freezing stress in transgenic alfalfa (Medicago sativa L.). Plant Physiol., 103: 1155-1163.
Direct Link - McNeil, S.D., D. Rhodes, B.L. Russel, M.L. Nuccio, Y. Shachar-Hill and A.D. Hanson, 2000. Metabolic modeling identifies key constraints on an engineered glycine betaine synthesis pathway in tobacco. Plant Physiol., 124: 153-162.
Direct Link - Mittler, R. and E. Blumwald, 2010. Genetic engineering for modern agriculture: Challenges and perspectives. Annu. Rev. Plant Biol., 61: 443-462.
CrossRefDirect Link - Mukhoopadhyay, A., S. Vij and A.K. Tyagi, 2004. Overexpression of a zinc-finger protein gene from rice confers tolerance to cold, and salt stress in transgenic tobacco. Proc. Nat. Acad. Sci., 101: 6309-6314.
Direct Link - Nanjo, T., M. Kobayashi, Y. Yoshida, Y. Kakubari, K. Yamaguchi-Shinozaki and K. Shinozaki, 1999. Antisense suppression of proline degradation improves tolerance to freezing and salinity in Arabidopsis thaliana. Febs Lett., 461: 205-210.
Direct Link - Oh, S.J., C.W. Kwon, D.W. Choi, S.I. Song and J.K. Kim, 2007. Expression of barley HvCBF4 enhances tolerance to abiotic stress in transgenic rice. Plant Biotech. J., 5: 646-656.
PubMed - Pan, A., M. Yang, F. Tie, L. Li, Z. Chen and B. Ru, 1994. Expression of mouse metallothionein-I gene confers cadmium resistance in transgenic tobacco plants. Plant Mol. Biol., 24: 341-351.
PubMed - Park, B.J., Z. Liu, A. Kanno and T. Kameya, 2005. Genetic improvement of Chinese cabbage for salt and drought tolerance by constitutive expression of a B. napus LEA gene. Plant Sci., 169: 553-558.
CrossRef - Park, J.M., C.J. Park, S.B. Lee, B.K. Ham, R. Shin and K.H. Paek, 2001. Overexpression of the tobacco Tsi1 gene encoding an EREBP/AP2-type transcription factor enhance resistance against pathogen attach and osmotic stress in tobacco. Plant Cell, 13: 1035-1046.
Direct Link - Parvanova, D., A. Povova, I. Zaharivena, P. Lambrev and T. Kostantinova et al., 2004. Low temperature tolerance of tobacco plants transferred to accumulate proline, fructans, or glycine betaine. Variable chlophyll fluorescence evidence. Phytosynthetica, 42: 179-185.
Direct Link - Pavlicova, D., T. Macek, M. Mackova, J. Szakova and J. Balik, 2004. Cadmium tolerance and accumulation in transgenic tobacco plants with a yeast metallothionein combined with a polyhistidin tail. Int. Biodeterioration Biodegradation, 54: 233-237.
CrossRef - Pellegrineschi, A., M. Reynolds, M. Pacheco, R.M. Bitro, R. Almeraya, K. Yamaguchi-Shinazaki and D. Hoisington, 2004. Stress-induced expression in wheat of the Arabidopsis thaliana DREB1A gene delays water stress symptoms under green house condition. Genome, 47: 493-500.
CrossRefDirect Link - Perera, I.Y., C.Y. Hung, C.D. Moore, J. Stevenson-Paulik and W.F. Boss, 2008. Transgenic Arabidopsis plants expressing the type 1 inositol 5-phosphatase exhibit increased drought tolerance and altered abscisic acid signaling. Plant Cell, 20: 2876-2893.
Direct Link - Pilon-Smits, E.A.H., M.J.M. Ebskamp, M.J. Paul, M.J.W. Jeuken, P.J. Weibeck and S.C.M. Smeekens, 1995. Improved performance of transgenic fructan-accumulating tobacco under drought stress. Plant Physiol., 107: 125-130.
Direct Link - Pilon-Smits, E.A.H., M.J.M. Ebskamp, M.J.W. Jeuken, I.M. van der Meer, P.J. Weisbeck, R.G.F. Visser and S.C.M. Smeekens, 1996. Microbial production in transgenic potato plants and tubers. Indus. Crops Prod., 5: 35-56.
CrossRef - Pinto, A.P., A.M. Mota, A. De Varennes and F.C. Pinto, 2004. Influence of organic matter on the uptake of cadmium, zinc, copper and iron by sorghum plants. Sci. Total Environ., 326: 239-247.
Direct Link - Pomponi, M., V. Censi, D.V. Girolamo, A.D. Paolis, L.D. Toppi, R. Aromolo, P. Costantino, M. Cardarelli, 2006. Overexpression of Arabidopsis phytochelatin synthase in tobacco plants enhances Cd2+ tolerance and accumulation but not translocation to the shoot. Planta, 223: 180-190.
Direct Link - Prasad, S.R., P.G. Bagali, S. Hittalmani and H.E. Shaahidhar, 2000. Molecular mapping of quantitative trait loci associated with seedling tolerance to salt stress in rice (Oryza sativa L.). Curr. Sci., 78: 162-164.
Direct Link - Queitsch, C., S.W. Hong, E. Vierling and S. Lindquist, 2000. Heat shock protein 101 plays a crucial role in thermotolerance in Arabidopsis. Plant Cell, 12: 479-492.
Direct Link - Rahaman, M., A. Grover, W.J. Peacock, E.S. Dennis and M.H. Ellis, 2001. Effects of manipulation of pyruvate decarboxylase and alcohol dehydrogenase levels on the submergence tolerance of rice. Austr. J. Plant Physiol., 28: 1231-1241.
Direct Link - Richards, R.A., 1995. Improving crop production on salt-affected soils: By breeding or management?. Exp. Agric., 31: 395-408.
Direct Link - Rohila, J.S., R.K. Jain and R. Wu, 2002. Genetic improvement of Basmati rice for salt and drought tolerance by regulated expression of a barley Hva1 cDNA. Plant Sci., 163: 525-532.
CrossRef - Roxas, V.P., R.K. Jr. Smith, E.R. Allen and R.D. Allen, 1997. Over expression of glutathione-S-transferase/glutathione peroxidase enhances the growth of transgenic tobacco seedlings during stress. Nature Biotechnol., 15: 988-991.
Direct Link - Roy, B. and A.B. Mandal, 2005. Anther culture response in indica rice and variations in major agronomic characters among the androclones of a scented cultivar, Karnal local. Afr. J. Biotechnol., 4: 235-240.
Direct Link - Roy, B. and A.B. Mandal, 2006. Rapid and recurrent in vitro mass-multiplication of androgenic rice embryos. Indian J. Biotechnol., 5: 239-242.
Direct Link - Roy, B. and A.B. Mandal, 2011. Profuse microtillering of androgenic plantlets of elite indica rice variety IR 72. Asian J. Biotechnol., 3: 165-176.
CrossRefDirect Link - Roy, M. and R. Wu, 2001. Arginine decarboxylase transgene expressing and analysis of environmental stress tolerance in transgenic rice. Plant Sci., 160: 869-875.
CrossRef - Di Toppi, L.S. and R. Gabbrielli, 1999. Response to cadmium in higher plants. Environ. Exp. Bot., 41: 105-130.
CrossRefDirect Link - Sasaki, T., Y. Yamamoto, B. Ezaki, M. Katsuhara and S.J. Ahn et al., 2004. A wheat gene encoding an aluminum-activated malate transporter. Plant J., 37: 645-653.
PubMedDirect Link - Schellenbaum, L., N. Sprenger, H. Schuepp, A. Wiemken and T. Boller, 1999. Effects of drought, transgenic expression of a fructan synthesizing enzyme and of mycorrhizal symbiosis on growth and soluble carbohydrate pools in tobacco plants. New Phytol., 142: 67-77.
Direct Link - Seki, M., M. Narusaka, J. Ishida, T. Nanji and M. Fujita et al., 2002. Monitoring the expression profiles of 7000 Arabidopsis genes under drought, cold and high-salinity stresses using a full-length cDNA microarray. Plant J., 31: 279-292.
Direct Link - Seki, M., M. Narusaka, H. Abe, M. Kasuga and K. Yamaguchi-Shinozaki et al., 2001. Monitoring the expression pattern of 1300 Arabidopsis genes under drought and cold stresses by using a full-length cDNA microarray. Plant Cell, 13: 61-72.
CrossRefPubMedDirect Link - Gupta, A.S., J.L. Heinen, A.S. Haladay, J.J. Barke and R.D. Allen, 1993. Increased resistance to oxidative stress in transgenic plants that overexpress chloroplastic Cu/Zn superoxide dismutase. Proc. Natl. Acad. Sci., 90: 1629-1633.
Direct Link - Serrano, R., F.A. Culianz-Macia and V. Moreno, 1998. Genetic engineering of slat and drought tolerance with yeast regulatory genes. Sci. Horticulturae, 78: 261-269.
CrossRef - Blum, A., R. Munns, J.B. Passioura, N.C. Turner and R.E. Sharp et al., 1996. Genetically engineered plants resistant to dry soil and salt stress: How to interpret osmotic relations. Plant Physiol., 110: 1051-1053.
Direct Link - Shelveleva, E., W. Chamara, H.J. Bohnert and R.G. Jensen, 1997. Increased salt and drought tolerance by D-ononitol production in transgenic Nicotiana tabacum L. Plant Physiol., 115: 1211-1219.
Direct Link - Shen, Y.G., B.X. Du, W.K. Zhang, J.S. Zhang and S.Y. Chen, 2002. AhCMO, regulated by stresses in Atriplex hortensis, can improve drought tolerance in transgenic tobacco. Theoret. Applied Genet., 105: 815-821.
CrossRefDirect Link - Shou, H.X., P. Bordallo and K. Wang, 2004. Expression of Nicotiana protein kinase (NPK1) enhance drought tolerance in transgenic maize. J. Exp. Bot., 55: 1013-1019.
Direct Link - Sivamani, E., A. Bahieldin, J.M. Wraith, T. Al-Niemi, W.E. Dyer, T.D. Ho and R. Qu, 2000. Improved biomass productivity and water use efficiency under water deficit conditions in transgenic wheat constitutively expressing the barley HVA1 gene. Plant Sci., 155: 1-9.
PubMedDirect Link - Song, W.Y., E.J. Sohn, E. Martino, Y.J. Lee and Y.Y. Yang et al., 2003. Engineering tolerance accumulation of lead and cadmium in transgenic plants. Nature Biotechnol., 21: 914-919.
Direct Link - Steponkus, P.I., M. Uemura, R.A. Joseph, S. Gilmour and M.F. Thomashow, 1998. Mode of action of the COR15a gene on the freezing tolerance of Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA., 95: 14570-14575.
Direct Link - Tanaka, Y., T. Hibino, Y. Hayasi, A. Tanaka and S. Kishitani et al., 1999. Salt tolerance of transgenic rice overexpressing yeast mitrochondrial Mn-SOD in chloroplasts. Plant Sci., 148: 131-138.
Direct Link - Tarezynski, M.C., R.G. Tensen and H.J. Bohnert, 1993. Stress protection of transgenic tobacco by production of the osmolyte mannitol. Science, 259: 508-510.
PubMed - Thomas, J.C., M. Sepathi, B. Arendall and H.J. Bohnert, 1995. Enhancement of seed germination in high salinity by engineering manitol expression in Arabidopsis thaliana. Plant Cell Environ., 18: 801-806.
Direct Link - Urao, T., K. Yamagnchi-Shinozaki and K. Shinozak, 2000. Two component systems in plant signal transduction. Trends Plant Sci., 5: 67-74.
PubMed - Veena Reddy, V.S. and S.K. Sopory, 1999. Glyoxalase I from Brassica juncea: Molecular cloning, regulation and its over-expression confer tolerance in transgenic tobacco under stress. Plant J., 17: 385-395.
PubMed - Wan, J.L., H.Q. Zhai, J.M. Wan and H. Ikehashi, 2003. Detection and analysis of QTLs for ferrous iron toxicity tolerance in rice (Oryza sativa L.). Euphytica, 13: 201-206.
Direct Link - Wang, W., B. Vinocur and A. Altman, 2003. Plant responses to drought salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta, 218: 1-14.
PubMed - Wang, Y., J. Ying, M. Kuzma, M. Chalifoux and A. Sample et al., 2005. Molecular tailoring of farnesylation for plant drought tolerance and yield protection. Plant J., 43: 413-424.
Direct Link - Wang, Y.C., G.Z. Qu, H.Y. Li, Y.J. Wu, C. Wang, G.F. Liu, and C.P. Yang, 2010. Enhanced salt tolerance of transgenic poplar plants expressing a manganese superoxide dismutase from Tamarix androssowii. Mol. Bio. Rep., 37: 1119-1124.
CrossRef - Winicov, I. and D.R. Bastola, 1999. Transgenic over expression of the transcription factor Alfin1 enhances expression of the endogenous MSPRP2 gene alfalfa and improve salinity tolerance of the plant. Plant Physiol., 120: 473-480.
Direct Link - Wu, L., Z. Fan, L. Guo, Y. Li, Z.L. Chen and L.J. Qu, 2005. Over-expression of the bacterial nhaA gene in rice enhances salt and drought tolerance. Plant Sci., 168: 297-302.
CrossRef - Xu, D., X. Duan, B. Wang, B. Hong, T.D. Ho and R. Wu, 1996. Expression of a late embryogenesis abundant protein gene, HVA1, from barley confers tolerance to water deficit and salt stress in transgenic rice. Plant Physiol., 110: 249-257.
Direct Link - Xu, K., X. Xu, T. Fukao, P. Canlas and R. Maghirang-Rodriguez et al., 2006. Sub1A is an ethylene-response-factor-like gene that confers submergence tolerance to rice. Nature, 442: 705-708.
CrossRefDirect Link - Zhang, H.X. and E. Blumwald, 2001. Transgenic salt tolerant tomato plants accumulate salt in the foliage but not in the fruits. Nat. Biotechnol., 19: 765-768.
PubMed - Zhang, H.X., J.N. Hodson, J.P. Williams and E. Blumwald, 2001. Engineering salt tolerant Brassica plants: Characterization of yield and seed oil quality in transgenic plants with increased vacuolar sodium accumulation. Proc. Nat. Acad. Sci. USA., 48: 12832-12836.
CrossRefDirect Link - Zhu, B., J. Sua, M. Changa, D.P.S. Verma, Y.L. Fan and R. Wu, 1998. Overexpression of a Δ1 pyrroline-5-carboxylate synthetase gene and analysis of tolerance to water-and salt-stress in transgenic rice. Plant Sci., 139: 41-48.
CrossRef - Zhu, Y.L., E.A.H. Pilon-Smits, A.S. Tarun, S.U. Weber, L. Jouanin and N. Terry, 1999. Cadmium tolerance and accumulation in Indian mustard is enhanced by overexpressing γ-glutamylcysteine synthetase. Plant Physiol., 121: 1169-1177.
Direct Link - Wang, Y.P., Q.C. Liu, A.X. Li, H. Zhai, S.S. Zhang and B.L. Liu, 2003. In vitro selection and identification of drought-tolerant mutants in sweet potato. Agric. Sci. China, 2: 1314-1320.
Direct Link