
B. Ghosh
Microbiology Research Laboratory, Department of Zoology, Molecular Biology and Genetics, Presidency College, 86/1, College Street, Kolkata 700 073, India
R.R. Ray
Microbiology Research Laboratory, Department of Zoology, Molecular Biology and Genetics, Presidency College, 86/1, College Street, Kolkata 700 073, India
Biotechnology
Year: 2010 | Volume: 9 | Issue: 2 | Page No.: 224-228







ABSTRACT
Extra cellular isoamylase from Rhizopus oryzae PR7 MTCC 9642 was found to saccharify soluble potato starch and various native raw starches collected from domestic effluents, of which arrow root, tamarind kernel, tapioca and oat were noteworthy. In the present study, the effects of incubation time, pH, temperature, substrate concentration, gelatinization and chemical pretreatment of starches on saccharification were quantified in detail. Highest extent of saccharification was found at 55°C. Except tamarind kernel powder, the best pH for bioconversion of all other substrates was found to be at 5.0. The maximum amount of bioconversion was accomplished within 60 min of incubation with a substrate concentration ranging from 0.5 to 1% (w/v). About 85-200% increase in sugar production was found after thermal pretreatment of the starches. Amongst the chemicals, Mn2+ brought about a uniform increase in sugar production for all the substrates used, whereas thiol compounds could also increase the extent of saccharification.
PDF Abstract XML References Citation
How to cite this article
B. Ghosh and R.R. Ray, 2010. Saccharification of Raw Native Starches by Extracellular Isoamylase of Rhizopus oryzae. Biotechnology, 9: 224-228.
DOI: 10.3923/biotech.2010.224.228
URL: https://scialert.net/abstract/?doi=biotech.2010.224.228
DOI: 10.3923/biotech.2010.224.228
URL: https://scialert.net/abstract/?doi=biotech.2010.224.228
INTRODUCTION
Starch occurs as highly organized structures and has unique thermal properties and functionality that have permitted its wide use in food products and industrial application (Ratnayake and Jackson, 2009). Bioconversion of starch may be accomplished by acid treatment or enzymatic hydrolysis, of which the latter is preferred as it offers a number of advantages including improved yields and favorable economics (Satyanarayana et al., 2004). Moreover, recent discoveries on the use of micro organisms as sources of industrially relevant enzymes have led to an increased interest in the application of microbial enzymes in various industrial processes (Alva et al., 2007).
Today, food brewing and pharmaceutical industries depend solely on various extra cellular microbial enzymes (Abu et al., 2005) and enzymatic hydrolysis of starch can be used to obtain various valuable hydrolyzates with different compositions (Baks et al., 2008).
Starch is degraded by amylases, of which isoamylase (glycogen-6-glucanohydrolase, E.C.3.2.1.68) hydrolyses 1, 6- α-D-glycosidic linkages of glycogen, amylopectin and α and β limit dextrins, producing linear malto oligosaccharides (Fang et al., 1994). Isoamylase is used primarily in the production of food ingredients from starch like glucose, maltose, trehalose and cyclodextrins (Olemposka-Beer, 2007).
Conventionally, conversion of starch to glucose uses cooked or pre gelatinized substrate and thus is energy intensive and expensive process. With the view of reducing the energy consumption, currently there is considerable research on raw starch degrading enzymes (Sun et al., 2008), which eventually curtail the cost of sugar production in food industries. Only few micro organisms have been reported to possess the ability to produce raw starch degrading amylase (Abu et al., 2005) and almost no published report is available on raw starch debranching fungal isoamylase.
In the present study, saccharification of various native starches, in their uncooked form, by the extra cellular isoamylase synthesized by Rhizopus oryzae was reported with elucidation of various factors influencing the extent of saccharification.
MATERIALS AND METHODS
Enzyme source: The present study was carried out during early summer to early winter (March to December, 2009) of this year. A strain of Rhizopus oryzae PR7 MTCC 9642, isolated from Eastern India was grown in basal medium composed of (g L-1): peptone 0.9; (NH4)2HPO4 0.4; KCl 0.1; MgSO4.7H2O 0.1 and starch 0.25 (pH: 8) at 28°C for 72 h. The culture broth was filtered, the filtrate was centrifuged at 5,000 g for 5 min and the supernatant was used as enzyme source.
Enzyme assay: Isoamylase activity was measured by incubating the assay mixture (1 mL) containing an equal volume of enzyme and 1% (w/v) Oyster glycogen in 0.1 (M) phosphate buffer (pH-5) at 55°C for 5 min. The reducing sugar released was measured by the dinitrosalicylic acid method (Bernfeld, 1955) taking glucose as standard. Blanks were prepared with inactivated enzymes. One unit of isoamylase was defined as that amount of enzyme that liberated 1 μmole of glucose/mL/min of reaction (Ara et al., 1993).
Saccharification of native raw starches: The native starches namely arrow root (rhizome of Maranta arundinacea), oat (Avena sativa), tamarind kernel, tapioca (Manihot esculenta), millet, arum and pulse dust were collected from market dumps and domestic effluents, washed thoroughly with water, air dried, pulverized and sieved to 40 mesh particle size, before using as substrate for saccharification.
Dry starch granules (10 mg) were incubated with 1 mL of isoamylase diluted with 0.1 (M) phosphate buffer (pH: 5) in a screw capped tube for various time periods at 55°C. The resultant supernatant following centrifugation at 2000 g for 2 min was analyzed by DNSA method (Bernfeld, 1955) using glucose as standard.
The percentage of saccharification was calculated (Baig et al., 2004) as:
Effect of various factors on saccharification: To determine the effect of incubation time, saccharification was carried out for various time periods at optimum conditions. The role of pH on saccharification was determined by varying the pH of the assay mixture from 4 to 7. The effects of increasing substrate concentration were estimated by changing the substrate concentrations keeping other factors unchanged. To check the effects of chemical pretreatment, starch residues were treated with 10 mM of metal ions, thiol compounds and thiol inhibitors for 10 min before using them as substrate of saccharification mixture. The effect of cooking or gelatinization was checked by heating the starch suspension at 100°C for 10 min before enzyme treatment.
RESULTS
The enzyme was able to hydrolyse many types of starch molecules (Fig. 1) of which apart from potato starch (Merck), other types of native starches showing promising results, were starch granules extracted from arrow root, tamarind kernel, oat and tapioca.
 | |
| Fig. 1: | Saccharification of various native starches by Isoamylase of Rhizopus oryzae |
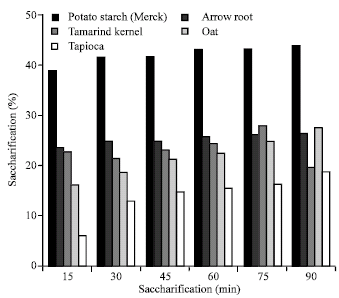 | |
| Fig. 2: | Effect of incubation time on saccharification of native starches by Isoamylase of Rhizopus oryzae |
Therefore, further experiments were carried out with the bioconversion of these starch residues only.
The maximum amount of bioconversion was accomplished within 60 min of incubation (Fig. 2) which was followed by a slow rate of increase in sugar production, but it took longer time for tamarind kernel and oat starch.
As pH of the saccharification media is one of the important determinants of bioconversion process,saccharification was tested at various pH, which showed that except tamarind kernel powder, the best pH for bioconversion was 5.0 (Fig. 3).
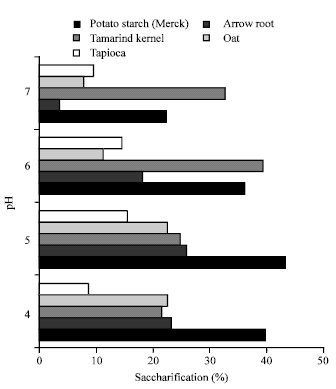 | |
| Fig. 3: | Effect of pH on saccharification of native starches by Isoamylase of Rhizopus oryzae |
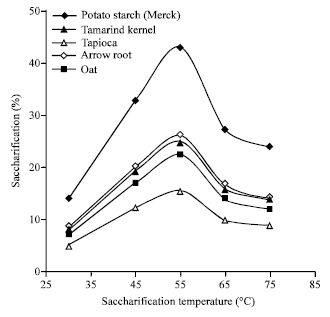 | |
| Fig. 4: | Effect of temperature on saccharification of native starches by Isoamylase of Rhizopus oryzae |
On the other hand, the highest amount of sugar production was found at a temperature of 55°C (Fig. 4) for all types of starch tested.
Saccharification rate was found to increase with the increase in substrate concentration (Fig. 5) up to 1% (w/v). But, with further increase in substrate concentration to 1.5% (w/v), the sugar production dropped in cases of potato starch (Merck), tamarind kernel powder and oat starch.
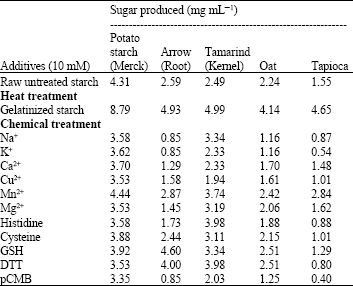
| Table 1: | Effect of substrate treatments on saccharification of starch molecules |
 | |
| Substrate: 10 mg mL -1 | |
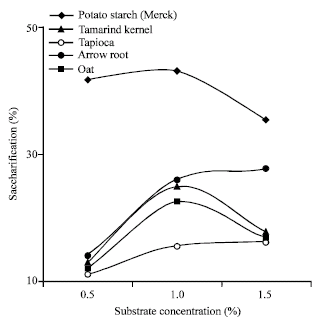 | |
| Fig. 5: | Effect of starch concentration on saccharification by Isoamylase of Rhizopus oryzae |
As pretreatment of substrate plays an important role in saccharification, the starch granules were treated both thermally and chemically, which showed a remarkable increase in sugar production after gelatinization or cooking. An 85-200% increase in sugar production (Table 1) was found after gelatinization. Amongst the chemicals, Mn2+ brought about a uniform increase in sugar production for all the substrates used, whereas thiol compounds like reduced glutathione (GSH) and dithiothreitol (DTT) enhanced the sugar production up to a certain extent.
DISCUSSION
Starches extracted from different sources showed different susceptibility towards enzymic hydrolysis. These differential rates of saccharification could be explained by their different granular morphology (Howling, 1989).
For all types of substrates, the sugar production rate was initially high which gradually reduced after certain time, probably due to substrate and/or enzyme limitation or as a result of product accumulation and consequent product inhibition (Ray et al., 1994).
The best pH for saccharification by isoamylase of R. oryzae PR7 was slightly acidic, similar to the optimum pH of bacterial isoamylases (Castro et al., 1992; Wu et al., 1994; Fang et al., 2005). The higher pH required for tamarind kernel powder suggested the existence of distinct crystalline organization (Colonna et al., 1992) of the starch granule extracted from it.
The optimum temperature of the isoamylase from the present strain for sugar production was higher than the temperature optima (30-40°C) reported from other microbial isoamylases (Ueda and Nanri, 1967; Fang et al., 1994; Ara et al., 1993; Spencer-Martins, 1982; Olemposka-Beer, 2007), which indicated that the isoamylase from the present strain may be used conveniently at moderately high temperature in sugar manufacturing industries.
Although, there was an initial positive correlation between starch concentration and sugar production, with further increase in substrate concentration there came a decline in saccharification, probably due to enzyme limitation. The difference in the level of substrate saturation amongst the native starches tested could be attributed to the nature of the starch granules, as amylolytic hydrolysis of native starch granule is governed by the specific surface area, not by the mass concentration of each granule (Kim et al., 2008).
Gelatinization brought about a remarkable increase in saccharification, as the kinetics of the reaction at lower temperature were much slower than above the gelatinization temperature where the starch exists in a molecularly dispersed sol in the presence of an excess of water (Howling, 1989) and hydrothermic action on starch molecules enhanced its rate of depolymerization that made them more accessible to enzyme action.
The enhancement of saccharification by Mn2+ might be achieved by the increased activity of the enzyme used, but thiol compounds probably increased the affinity of the substrate towards the enzyme as indicated by the differential response by different types of starch.
In view of energy cost, effective utilization of natural resources, direct hydrolysis of starch below the gelatinization temperature is desirable (Kelly et al., 1995). The sacchrification potential of the isoamylase produced by the present strain may add economy in sugar production from raw native starches.
ACKNOWLEDGMENT
The authors wish to thank University Grants Commission, India for financial assistance.
REFERENCES
- Abu, E.A., S.A. Ado and D.B. James, 2005. Raw starch degrading amylase production by mixed culture of Aspergillus niger and Saccharomyces cerevisae grown on sorghum pomace. Afr. J. Biotechnol., 4: 785-790.
Direct Link - Alva, S., J. Anupama, J. Savla, Y.Y. Chiu and P. Vyshali et al., 2007. Production and characterization of fungal amylase enzyme isolated from Aspergillus sp. JGI 12 in solid state culture. Afr. J. Biotechnol., 6: 576-581.
Direct Link - Ara, K., K. Saeki and S, Ito, 1993. Purification and characterization of an alkaline isoamylase from an alkalophilic strain of Bacillus. J. General Microbiol., 139: 781-786.
Direct Link - Baig, M.M.V., M.L.B. Baig, M.A.I. Baig and M. Yasmeen, 2004. Saccharification of banana agro-waste by cellulolytic enzymes. Afr. J. Biotechnol., 3: 447-450.
Direct Link - Baks, T., M.E. Bruins, A.M. Matser, A.E.M. Janssen and R.M. Boom, 2008. Effect of gelatinization and hydrolysis conditions on the selectivity of starch hydrolysis with alpha-amylase from Bacillus licheniformis. J. Agric. Food Chem., 56: 488-495.
CrossRefDirect Link - Castro, G.R., G.F. Garcia and F. Sineriz, 1992. Extracellular isoamylase produced by Bacillus circulans MIR-137. J. Applied Microbiol., 73: 520-523.
CrossRef - Colonna, P., V. Leloup and A. Boleon, 1992. Limiting factors of starch hydrolysis. Eur. J. Clin. Nutr., 46: S17-S32.
PubMedDirect Link - Fang, T.Y., L.L. Lin and W.H. Hsu, 1994. Recovery of isoamylase from Pseudomonas amyloderamosa by adsorption-elution on raw starch. Enzyme Microbiol. Technol., 16: 247-252.
Direct Link - Fang, T.Y., W.C. Tseng, C.J. Yu and T.Y. Shih, 2005. Characterization of the thermophilic isoamylase from the thermophilic archeon Sulfobolobus solfataricus ATCC 35092. J. Mol. Catalysis B: Enzymatic, 33: 99-107.
CrossRef - Kelly ,C.T., M.A. McTigue, E.M. Doyle and W.M. Fogarty, 1995. The raw starch-degrading alkaline amylase of Bacillus sp IMD 370. J. Ind. Microbiol. Biotechnol., 15: 446-448.
CrossRef - Kim, J.C., B.W. Kong, M.J. Kim and S.H. Lee, 2008. Amylolytic hydrolysis of native starch granules affected by granule surface area. J. Food Sci., 73: C621-C624.
CrossRef - Ratnayake, W.S. and D.S. Jackson, 2008. Starch gelatinization. Adv. Food. Nutr. Res., 55: 221-268.
CrossRefDirect Link - Satyanarayana, T., S.M. Noorwez, S. Kumar, J.L.U.M. Rao, M. Ezhilvannan and P. Kaur, 2004. Development of an ideal starch saccharification process using amylolytic enzymes from thermophiles. Biochem. Soc. Trans., 32: 276-278.
PubMedDirect Link - Spencer-Martins, I., 1982. Extracellular isoamylase production by theyeast Lipomyces kononenkoae. Applied Environ. Microbiol., 44: 1253-1257.
Direct Link - Sun, H., P. Zhao and M. Peng, 2008. Application of maltitol to improve production of raw starch digesting glucoamylase by Aspergillus niger F-08. World. J. Microbiol. Biotechnol., 24: 2613-2618.
CrossRefDirect Link - Ueda, S. and N. Nanri, 1967. Production of isoamylase by Escherichia intermedia. Applied Environ. Microbiol., 15: 492-496.
Direct Link - Wu, D.H., C.Y. Wen, L.L. Lin, W.S. Chu and W.H. Hsu, 1994. Effect of pH on isoamylase production by Pseudomonas amyloderamosa WU 5315. Lett. Applied Microbiol., 19: 67-69.
CrossRef