
M. Baserisalehi
Department of Microbiology, Islamic Azad University, Kazeroun Branch, Kazeroun, Iran
N. Bahador
Department of Microbiology, Islamic Azad University, Fars Science Research Branch, Shiraz, Iran
Biotechnology
Year: 2008 | Volume: 7 | Issue: 4 | Page No.: 813-817







ABSTRACT
Susceptibility of thermophilic Campylobacter isolates from environmental samples to antibiotics was studied to investigate relation between occurrence of plasmid and antibiotic resistant character in Campylobacter jejuni. Antimicrobial susceptibility of environmental isolates of Camp. jejuni, Camp. coli and Camp. lari to antibiotics was assessed by disc diffusion method afterward, three strains of Camp. jejuni F44, P41 and W21 isolates were subjected to plasmid isolation and curing. The results on antibiotic susceptibility of Campylobacter isolates by disc diffusion method indicated that, all the isolates of Campylobacter were sensitive to ciprofloxacin and resistant to cefotaxime, cephalexin and ampicillin. In addition, more than 50% of the Camp. jejuni isolates were resistance to chloramphenicol and erythromycin. Plasmids were detected from 60, 50 and 80% of the Camp. jejuni, Camp. coli and Camp. lari isolates, respectively. The cured strain of Camp. jejuni F44 was sensitive to chloramphenicol and resistant to erythromycin. Hence, the most of Campylobacter isolates tested harbored plasmid and probably chloramphenicol resistant marker is plasmid mediated while, erythromycin resistant marker is chromosomally mediated. Therefore, probably gene resistant markers present in the plasmids can be transmitted among campylobacters in the environment and reach the human population by direct contact and via food products of animal origin.
PDF Abstract XML References Citation
How to cite this article
M. Baserisalehi and N. Bahador, 2008. A Study on Relationship of Plasmid with Antibiotic Resistance in Thermophilic Campylobacter spp. Isolates from Environmental Samples. Biotechnology, 7: 813-817.
DOI: 10.3923/biotech.2008.813.817
URL: https://scialert.net/abstract/?doi=biotech.2008.813.817
DOI: 10.3923/biotech.2008.813.817
URL: https://scialert.net/abstract/?doi=biotech.2008.813.817
INTRODUCTION
Members of the Campylobacter genus are gram-negative, curved and S-shaped microaerophilic bacteria. They are responsible for human gastroenteritis throughout the world and enteritis caused by them in some places is more important than that of Salmonella and Shigella (Scotter et al., 1993). Antimicrobial chemotherapy of patients with acute Campylobacter enteritis involves treatment with erythromycin, tetracyclines and fluoroquinolones (Alfredson et al., 2003). However, frequency of occurrence of antibiotics resistance Campylobacter increasing in developed and developing countries (Taylor and Courvalin, 1988), but antibiotics therapy still is final remedy in case of patient with acute Campylobacter enteritis. Resistance of Campylobacter spp. to a number of antibiotics, such as tetracycline, erythromycin, ciprofloxacin, kanamycin, nalidixic acid and chloramphenicol has been reported by Piddock et al. (2000). Furthermore, the rate of fluoroquinolone-resistant campylobacters in Thailand, from 0-84% increased during 1990-1995 (Isenbarger et al., 2002). Resistance of Emergence and dissemination of antibiotic resistance among Campylobacter spp. have been linked to the use of antibiotics in veterinary medicine and use as prophylactics and growth promoters in animal husbandry (Piddock et al., 2000). The increasing rate of human infections caused by antimicrobial-resistant strains of Camp. jejuni makes clinical management of cases of campylobacteriosis more difficult, prolonging illness and compromising treatment of patients with bacteremia (Piddock, 1995).
Despite, deficiency of data in case of the molecular pathogenesis of campylobacters, many reports opined that antibiotic resistance markers in campylobacters could be chromosomally or plasmid-mediated (Taylor and Courvalin, 1988). For instance, tetracycline resistance in strains of Campylobacter jejuni and Campylobacter coli was mediated by plasmids (Taylor and Chau, 1996). Objectives of this study were assessed to determined susceptibility of thermophilic Campylobacter isolates to antibiotics. Afterward, plasmid isolation from pathogenic campylobacters was carried out to achieved information concerning to the presence of plasmid DNA in Campylobacter isolates and finally plasmid curing was done to seek correlation between resistance to antibiotics and plasmid occurrence in Camp. jejuni.
MATERIALS AND METHODS
Organisms: Seventy isolates belonging to Camp. jejuni, Camp. coli and Camp. lari were isolated from environmental sources viz., animal feces (cow and horse) and sewage in Kazeroun, Iran, 2007. The method for isolation of the bacteria was prêt KB method (Baserisalehi et al., 2004). All these isolates were maintained in Burcella broth with 15% glycerol at -15°C.
Antibiotic susceptibility by disc diffusion method: The antimicrobial susceptibility pattern of the strains under study was studied by the disc diffusion method (Bauer et al., 1966). To perform the disc diffusion test, each culture was grown in 5 mL of Muller-Hinton broth until the turbidity corresponded to 0.5 MacFarland standard tube (1.5x108 cells mL-1). The suspension was spread inoculated using sterile cotton swab onto Muller-Hinton agar plate and various antibiotic discs were placed on it. After incubating the plates at 37°C under microaerophilic conditions for 48 h the inhibition zones were recorded.
The antibiotic discs included: chloramphenicol (30 μg), norfloxacin (10 μg), kanamycin (30 μg), co-trimoxazole (25 μg), cefotaxime (30 μg), ampicillin (10 μg), ciprofloxacin (5 μg), tetracycline (30 μg), erythromycin (15 μg), gentamycin (10 μg), cephalexin (30 μg) (Hi Media Laboratories Limited, Mumbai).
Plasmid isolation: Plasmid was isolated using standard method recommended by Birnboim and Doly (1979). The quantitation and purity of DNA was done by determining 260/280 nm absorbance ratio spectra by spectrophotometer (UV-1601, Shimadzu).
Medium: Luria Bertani broth (LB) contained (g 100 mL-1) Yeast Extract 0.5, NaCl 0.5 and Tryptone 1. The pH was adjusted to 7.2 and sterilized at 121°C for 20 min.
Reagents:
| Solution I | : | :50 mM Glucose, 10 mM Na2EDTA, pH 8 and 25 mM Tris-Cl, pH 8 |
| Solution II | : | 0.2N NaOH and 1% SDS |
| Solution III | : | 3M Sodium acetate, pH 4.8-5.3 |
| TE buffer | : | 10 mM Tris-Cl, pH 8 and 1mM Na2EDTA, pH 8 |
Single colony of each isolate was inoculated into 2 mL LB medium separately and incubated overnight at 37°C under microaerophilic conditions. The cells were harvested by centrifugation at 5000 rpm for 5 min at 4°C. The cell pellet was resuspended in 100 μL of solution I. Then 200 μL of solution II was added and the suspension was mixed gently and incubated on ice for 10 min. The solution III (150 μL) was added to the mixture and incubated on ice for 10 min. The mixture was centrifuged at 11500 rpm, 15 min at 4°C and the supernatant from each tube was transferred to fresh Eppendroff tubes. The mixture of phenol and chloroform in 1:1 proportion (500 μL) was added in the tubes and mixed properly. In each case the phases were separated by centrifugation at 11500 rpm, for 15 min at 4°C. Aqueous layer from each tube was transferred to fresh set of Eppendroff tubes and 1000 μL of 70% ethanol was added into each tube and incubated at room temperature for 10 min. The suspensions were centrifuged at 11500 rpm for 15 min at 4°C. The supernatant was removed by aspiration and discarded. The precipitated plasmid DNA was dried at room temperature for 30 min and dissolved in 50 μL TE buffer.
Confirmation of plasmid DNA by agarose gel electrophoresis: Electrophoresis of plasmid DNA was performed using horizontal gel electrophoresis. Agarose gel (0.8%) was prepared in TAE buffer (40 mM tris-HCl, 50 mM Sodium acetate, 1 mM EDTA; pH 8). Gel was run for two and half hours at 50 volts, stained for 30 min with ethidim bromide (0.5 μg mL-1). The plasmids were visualized using UV light in Alpha imager gel documentation system (Alpha Innotech Corp., USA).
Plasmid curing
Plasmid curing by chemical agents: Three Camp. jejuni isolates (F44, P41 and W21) randomly were selected and subjected to plasmid curing by chemical agents and elevated temperature. Chemical agents used for plasmid curing were, Acridine orange, Acriflavine, Ethidium bromide (Intercalating dyes) and Rifampin. To perform plasmid curing, stock solutions of curing agents were prepared in distilled water. The curing agents were serially diluted in LB broth (1600 to 1.6 μg mL-1). Overnight growth (0.1 mL) (adjusted to No. 0.5 McFarland tube 1.5x108 cfu mL-1) of each isolate was inoculated into each of the tubes containing 1 mL LB with increasing concentrations of curing agent and incubated at 37°C for 48 h. The Minimal Inhibitory Concentration (MIC) and the Minimal Bactericidal Concentration (MBC) were determined by observing, absence of growth in brain heart infusion broth and on nutrient agar at the lowest concentration of curing agent, respectively. The curing agents diluted to Subinhibitory Concentration (SIC) and were incorporated into the LB agar at SIC values. The overnight growth of resistant cultures was spread on LB agar with SIC of curing agent and incubated at 37°C for 2 days. The isolated colonies were picked up by sterile toothpicks and inoculated on another LB agar plate (50 colonies/plate). The plates were incubated at 37°C for 48 h and used as master plates. The colonies from each master plate were replicated on Luria agar with 30 μg mL-1 chloramphenicol and on Luria agar with 15 μg mL-1 Erythromycin and incubated at 37°C for 48 h. Those colonies, which failed to grow on selective medium, were regarded as cured colonies.
At the same time, a control for curing of each marker was maintained by inoculating non-cured culture on selective media to have a check on spontaneous loss of resistant markers.
Characterization of cured and non-cured Camp. jejuni isolates: Morphology of the cured and non-cured Camp. jejuni was evaluated using phase contrast microscope (Nikon, Japan). They were subjected to phenotypic identification tests recommended by Atabay and Corry (1997). These tests included, H2S lead acetate strip, nitrate reduction, growth in 1% glycine and 3.5% NaCl, growth at different temperatures, viz., 25, 37 and 42°C and resistance to nalidixic acid (30 μg disc) and cephalothin (30 μg disc). Additional tests were hippurate hydrolysis, indoxyl acetate hydrolysis, urease production, alkaline phosphotase production and Glucose fermentation.
RESULTS
Antibiotic susceptibility of Campylobacter isolates: The results on antibiotic susceptibility of thermophilic Campylobacter isolates by disc diffusion method indicated that all the isolates of Campylobacter were sensitive to ciprofloxacin and resistant to cefotaxime, cephalexin and ampicillin. Besides, all the Camp. lari isolates were resistant to co-trimoxazole. All isolates of Camp. coli were sensitive to tetracycline. Amongst Camp. jejuni isolates, 74 and 70% of them were sensitive to gentamicin and kanamycin while 59 and 55% of them were sensitive to erythromycin and norfloxacin respectively. Less than 50% of the Camp. jejuni isolates were sensitive to chloramphenicol, tetracycline and co-trimoxazole and less than 50% isolates of Camp. coli were sensitive to rest of the antibiotics except co-trimoxazole and chloramphenicol. The number of Camp. coli isolates sensitive to antibiotics was relatively less than that of Camp. jejuni. Besides, less than 50% of Camp. lari isolates were sensitive to chloramphenicol, gentamicin, norfloxacin, kanamycin and erythromycin except tetracycline (Table 1).
Plasmid isolation: A set of forty isolates of Campylobacter was randomly selected and subjected to detection of plasmid by alkali lysis method. As shown in Table 2, 60% Camp. jejuni, 50% Camp. coli and 80% Camp. lari isolates harboured plasmids with ≥21 kb in size. The results indicated that the frequency of occurrence of the plasmid in Camp. lari isolates was relatively high while in Camp. coli was relatively low. Based on these observations, no correlation has been found between sources of isolates and presence of plasmids in Campylobacter isolates. Purity of plasmid DNA was found to be between 1.4-1.9 corresponding to 74.2-121 μg DNA mL-1.
Plasmid curing: Curing is to confirm whether the genes for resistance are encoded by genomic DNA or plasmid DNA. Here, attempt was made to cure antibiotic resistant marker from Camp. jejuni isolates using chemical agents and physical agent (elevated temperature).
Camp. jejuni isolates F44, P41 and W21 were subjected to plasmid curing by chemical agents. The results obtained indicated that MIC, SIC and MBC values of rifampicin were relatively high while, that of Ethidium bromide were relatively low. The frequency of plasmid curing induced by rifampicin was 4%. Plasmid curing was observed only in Camp. jejuni F44. Acridine orange, Acriflavine and Ethidium bromide could not cure Camp. jejuni (Table 3).
However, erythromycin and chloramphenicol were considered as resistant markers for plasmid curing. But all cured Camp. jejuni showed loss of resistance to chloramphenicol (Table 4).
Characterization of cured and original Camp. jejuni isolates: The results obtained from characterization of cured and original Camp. jejuni indicated that cured and original Camp. jejuni exhibited similar behavior regarding all tests as well as morphology.
| Table 1: | Susceptibility of environmental campylobacters by disc diffusion method |
 | |
| *Ch: Chloramphenicol, Ce: Cephalexin, No: Norfloxacin, Ka: Kanamycin, Co: Co-trimoxazole, Cf: Cefotaxime, Am: Ampicillin, Ci: Ciprofloxacin, Te: Tetracycline, Er: Erythromycin, Ge: Gentamicin | |
| Table 2: | Plasmid isolation from Campylobacter isolates |
 | |
| Plasmids isolated by Birnboim and Doly (1979) method | |
| Table 3: | Susceptibility of Campylobacter jejuni isolates to chemical curing agents |
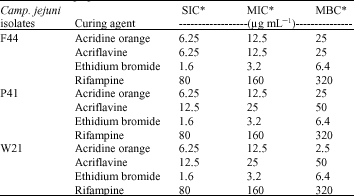 | |
| *MIC: Minimal Inhibitory Concentration, *MBC: Minimal Bactericidal Concentration, *SIC: Subinhibitory Concentration | |
| Table 4: | Curing of plasmid in Campylobacter jejuni |
 | |
| *chr: Chloramphenicol resistant marker, -: No strain was cured | |
DISCUSSION
Antimicrobial resistance property can come up through acquisition of genetic material encoding enzymes that inactivate a particular antibiotic (Hoffman, 1999). Gene resistance markers in campylobacters can be presented in plasmid, chromosome or both. For instance, erythromycin resistance has been reported previously as by chromosomal genes, whereas chloramphenicol resistance is plasmid encoded. It has been reported that resistance to chloramphenicol, kanamycin and tetracycline in Camp. jejuni is plasmid-mediated, while resistance to rest of the antibiotics is chromosomally mediated (Taylor and Courvalin, 1988; Taylor and Chau, 1996). On the other hand, several reports illustrated that some virulence factors of Campylobacter jejuni are associated with existence of the plasmid in the bacterium (Bacon et al., 2000; Tracz et al., 2005).
Present findings from this study indicated that out of all Campylobacter tested; plasmids were detected in 60% Camp. jejuni, 50% Camp. coli and 80% Camp. lari isolates with ≥21 kb in size. It means frequency of plasmid occurrence in Camp. lari is relatively high while, in Camp. coli was relatively low. Hence, it could be interpreted that occurrence of the plasmid in the Camp. lari isolates with high frequency might induce antibiotic resistant property to the bacterium.
Although Campylobacter jejuni is most important causes of bacterial diarrhea worldwide (Taylor, 1992), the details of its molecular pathogenesis are not well understood (Bacon et al., 2000). Thus, the present study was undertaken to carry out plasmid curing in order to achieve maximum information concerning to location of resistant markers in this bacterium.
Plasmid curing defined as a loss of plasmid from cell, which leads to loss of specific phenotypes such as drug resistance (Bouanchaud et al., 1969). To perform plasmid curing three strains of Camp. jejuni (F44, P41 and W21) isolates were subjected to plasmid curing to find out location of resistant markers of chloramphenicol and erythromycin. The result obtained indicated that the frequency of plasmid curing induced by elevated temperature and rifampicin was 8 and 4%, respectively while, Acridine orange, Acriflavine and Ethidium bromide could not cure Camp. jejuni. According to the data, only Camp. jejuni F44 was cured and the cured strain were sensitive to chloramphenicol and resistant to erythromycin. Therefore, it can be concluded that probably erythromycin resistant marker in Camp. jejuni is chromosomally mediated, while chloramphenicol resistant marker is plasmid mediated. This finding is supported by Wang and Taylor (1990). Although, according to present data chloramphenicol and erythromycin resistant markers in Camp. jejuni is plasmid and chromosomally mediated respectively, Dasti et al. (2007) reported, tetracycline resistance in Camp. jejuni, could be plasmid or chromosomally mediated.
The results obtained from characterization of cured and original strain of Camp. jejuni F44 using phenotypic identification tests indicated that no significant difference has been found between cured and original Camp. jejuni F44. Therefore, it can be concluded that the most of phenotypic characters of Camp. jejuni are associated with primary metabolism and its metabolites is chromosomally mediated.
Furthermore, present finding indicated that plasmid mediated of some antibiotic resistance markers in campylobacters cause transmission of the resistance markers among these bacteria and finally reach the human population by direct contact and via food products of animal origin.
REFERENCES
- Alfredson, D.A., R.J. Akhurst and V. Korolik, 2003. Antimicrobial resistance and genomic screening of clinical isolates of thermophilic Campylobacter spp. from South-East Queensland, Australia. J. Applied Microbiol., 94: 495-500.
Direct Link - Atabay, H.I. and J.E.L. Corry, 1997. The prevalence of campylobacters and arcobacters in broiler chickens. J. Applied Microbiol., 83: 619-626.
CrossRefDirect Link - Bacon, D.J., R.A. Alm, D.H. Burr, L. Hu and D.J. Kopecko et al., 2000. Involvement of a plasmid in virulence of Campylobacter jejuni 81-176. Infect. Immunity, 68: 4384-4390.
CrossRefDirect Link - Baserisalehi, M., N. Bahador and B.P. Kapadnis, 2004. A novel method for isolation of Campylobacter spp. from environmental samples, involving sample processing and blood and antibiotic free medium. J. Applied Microbiol., 97: 853-860.
Direct Link - Bauer, A.W., W.M.M. Kirby, J.C. Sherris and M. Turck, 1966. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol., 45: 493-496.
CrossRefPubMedDirect Link - Birnboim, H.C. and J. Doly, 1979. A rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucleic Acids Res., 7: 1513-1523.
CrossRefPubMedDirect Link - Bouanchaud, D.H., M.R. Scauizzi and Y.A. Chabbert, 1968. Elimination by ethidium bromide of antibiotic resistance in Enterobacteria and Staphylococci. J. Gen. Microbiol., 54: 417-425.
CrossRef - Dasti, J.I., U. Grob, S. Pohl, R. Lugert, M. Weig and R.S. Ott, 2007. Role of the plasmid-encoded tet(O) gene in tetracycline-resistant clinical isolates of Campylobacter jejuni and Campylobacter col. J. Med. Microbiol., 56: 833-837.
CrossRef - Hoffman, P.S., 1999. Antibiotic resistance mechanisms of Helicobacter pylori. Can. J. Gastroenterol., 13: 243-249.
PubMedDirect Link - Piddock, I.J.N., V. Ricci, K. Stanley and K. Jones, 2000. Activity of antibiotics used in human and medicine for Campylobacter jejuni isolated from farm animals and their environment in Lancashire, UK. J. Antimicrob. Chemother., 46: 303-306.
Direct Link - Piddock, L., 1995. Quinolone resistance and Campylobacter spp. Antimicrob. Agents Chemother., 36: 891-898.
Direct Link - Scotter, S.L., T.J. Humphrey and A. Henely, 1993. Methods for detection of thermotolerant campylobacters in food: Results of an interlaboratory study. J. Bacteriol., 74: 155-163.
Direct Link - Taylor, D.E. and P. Courvalin, 1988. Mechanisms of antibiotic resistance in Campylobacter species. Antimicrob. Agent Chemother., 32: 1107-1112.
PubMedDirect Link - Taylor, D.E., 1992. Genetics of Campylobacter and Helicobacter. Annu. Rev. Microbiol., 46: 35-64.
PubMedDirect Link - Taylor, D.E. and A. Chau, 1996. Tetracycline resistance mediated by ribosomal protection. Antimicrob. Agents Chemother., 40: 1-5.
Direct Link - Tracz, D.M., M. Keelan, J. Ahmed-Bentley, A. Gibreel, K. Kowalewska-Grochowska and D.E. Taylor, 2005. pVir and bloody diarrhea in Campylobacter jejuni enteritis. Emerg. Infect. Dis., 11: 838-843.
CrossRefDirect Link - Wang, Y. and D.E. Taylor, 1990. Chloramphinicol resistance in Campylobacter coli: Nucleotide sequence, expression and cloning vector construction. Gene, 94: 23-28.
PubMedDirect Link