
Siti Nurmadihah Ismail
Department of Industrial Biotechnology, Faculty of Bioscience and Bioengineering, University Technology Malaysia, 81300 Skudai, Johor, Malaysia
Aishah Mohd Taha
Department of Industrial Biotechnology, Faculty of Bioscience and Bioengineering, University Technology Malaysia, 81300 Skudai, Johor, Malaysia
Ng Hong Jing
Department of Industrial Biotechnology, Faculty of Bioscience and Bioengineering, University Technology Malaysia, 81300 Skudai, Johor, Malaysia
Roswanira Ab. Wahab
Department of Chemistry, Faculty of Science, University Technology Malaysia,81300 Skudai, Johor, Malaysia
Aidil Abdul Hamid
Pusat Pengajian Biosains and Bioteknologi, Fakulti Sains and Teknologi, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
Rolando V. Pakingking Jr.
Southeast Asian Fisheries Development Center, Department of Aquaculture, Tigbauan, Iloilo, 5021 Philippines
Fahrul Huyop
Department of Industrial Biotechnology, Faculty of Bioscience and Bioengineering, University Technology Malaysia, 81300 Skudai, Johor, Malaysia
Biotechnology
Year: 2008 | Volume: 7 | Issue: 3 | Page No.: 481-486







ABSTRACT
A bacterial strain tentatively identified as Pseudomonas sp. R1 was isolated from a paddy (rice) field that could degrade monochloroacetic acid (MCA) for concentrations ranging from 5 to 40 mM. Quantitative agreement between the amount of MCA introduced and chloride released was also found. MCA dehalogenase activity in this strain was found to be inducible. Cell-free extracts displayed dehalogenating activity with specific halogenated organic compound with no activity on dichloropropionic acid or monochloropropionic acid. The estimated Km values for MCA was 0.14 mM. The optimal pH range for MCA dehalogenase activity (between pH 6.5 and 8.0), whereas the thermal stability profile stable up to 50 °C. The results of our current study demonstrated the potential use of Pseudomonas sp. R1 as suitable biological agent for biodegradation of MCA in contaminated agricultural area.
PDF Abstract XML References Citation
How to cite this article
Siti Nurmadihah Ismail, Aishah Mohd Taha, Ng Hong Jing, Roswanira Ab. Wahab, Aidil Abdul Hamid, Rolando V. Pakingking Jr. and Fahrul Huyop, 2008. Biodegradation of Monochloroacetic Acid by a Presumptive Pseudomonas sp. Strain R1 Bacterium Isolated from Malaysian Paddy (Rice) Field. Biotechnology, 7: 481-486.
DOI: 10.3923/biotech.2008.481.486
URL: https://scialert.net/abstract/?doi=biotech.2008.481.486
DOI: 10.3923/biotech.2008.481.486
URL: https://scialert.net/abstract/?doi=biotech.2008.481.486
INTRODUCTION
Most halogenated compounds are major environmental pollutants. Numerous studies have demonstrated that monochloroacetic acid (MCA) is toxic to aquatic life such as fishes and in particular to algae. Mineralisation of chlorinated propionic acid for example MCA, monobromoacetic acid (MBA), D,L-2-chloropropionic acid (D,L2CP) and 2,2-dichloropropionic acid (2,2DCP) has been reported earlier by Kwok et al. (2007), Tsang et al. (1988), Allison et al. (1983) and Olaniran et al. (2004, 2001). Many soil microorganisms are capable of utilizing halogenated-substituted organic acids as their sole carbon sources for growth and this has been known since the early 1960s (Jensen, 1960). The hydrolytic cleavage of the carbon-halogen bond enables microorganisms to utilise halosubstituted alkanoic acids and these enzyme posses by these bacteria species are known as dehalogenases (Fetzner and Lingens, 1994). These enzymes are inducible releasing intermediates which can then be metabolized via central metabolic pathways. The dehalogenases to date demonstrated relatively broad substrate specificities. The isolation of microorganisms degrading halogenated compounds is the first step in understanding the biochemistry and genetics of bacterial dehalogenases. In future, this information can be used to improve and engineer bacterial strains for biodegradation of persisting halogenated compounds.
In this study we report the isolation and identification of a soil bacterial strain from a local paddy (rice) field that is able to use MCA as sole source of carbon and energy. The properties of the dehalogenating activity found in the cell-free extracts are described in terms of stability, kinetics and substrate specificity. Our investigations explore the possibility of the use of dehalogenase for treatment of common environmental pollutants particularly in agricultural soils.
MATERIALS AND METHODS
Chemicals: Various halogenated compounds of analytical grade were purchased from Sigma-Aldrich or Fluka (Germany). Bacterial agar from Difco and the rest of chemicals were of the highest purity commercially available.
Purification of haloalkanoic acids: A 1 M solution was adjusted to pH 1 by addition of concentrated nitric acid. The solution was extracted three times with an equal volume of ethyl acetate and the combined extracts dried over anhydrous sodium sulphate. The solution was evaporated to dryness under vacuo at 45 °C in a rotary evaporator. The residues was dissolved in distilled water and neutralized with 10 M NaOH, before being made to the original volume.
Isolation and identification of organism: The bacterial isolate used in this study was originally isolated from Paddy (rice) agricultural soil, where extensive used of herbicide and pesticide. About 5 g of soil sample were added into 250 mL shake flask liquid culture containing PJC minimal salts medium (pH 7.5) with 20 mM MCA as the sole carbon source. After several subculture in solid minimal media with increasing concentrations of MCA (5, 20, 40 and 50 mM) at pH 7.5, one pure isolate was obtained by repeated streaking on solid medium with 40 mM MCA as the sole carbon source. Strains were characterized by morphological and biochemical tests as described in Bergey`s Manual of Systematic Bacteriology (Holt et al., 1994).
PCR amplification of 16S rRNA gene for bacteria identification: The primers used to amplify 16S rRNA were forward primer, FD1 (5`-aga gtt tga tcc tgg ctc ag-3`) and reverse primer, rP1 (5`-acg gtc ata cct tgt tac gac tt-3`). The program used for amplification of 16S rRNA gene were: initial denaturation 94 °C (5 min), denaturation 94 °C (1 min), annealing 55 °C (1 min) and extension 74 °C (4 min).
Growth conditions: The culture was grown at 30 °C on a rotary shaker in 250 mL flasks containing 100 mL medium. The liquid PJC minimal media was prepared as 10x concentrated basal salts containing K2HPO4. 3H2O (42.5 g L-1), NaH2PO4. 2H2O (10.0 g L-1) and (NH4)2SO4 (25.0 g L-1). The trace metal salts solution was a 10x concentrate that contained nitriloacetic acid (NTA) (1.0 g L-1), MgSO4 (2.0 g L-1), FeSO4. 7H2O (120.0 mg L-1), MnSO4. 4H2O (30.0 mg L-1), ZnSO4. H2O (30 mg L-1) and CoCl2 (10.0 mg L-1) in distilled water (Hareland et al., 1975). Minimal media for growing bacteria contained 10 mL of 10x basal salts and 10 mL of 10x trace metal salts per 100 mL of distilled water and were autoclaved (121 °C, for 15 min).
The carbon source MCA was neutralized with NaOH and sterilized by filtration and added to the autoclaved salts medium to an appropriate final concentration. The extent of growth determined by measuring the absorbance at A680 nm and the release of chloride. The bacteria were harvested by centrifugation during the late-logarithmic phase and the cells were washed three times in 0.1 M Tris-acetate buffer pH 7.5.
Preparation of cell-free extracts: Cell-free extracts were prepared from bacterial cells in late-exponential phase of growth. Cells from 100 mL culture were harvested by centrifugation at 10,000 g for 10 min at 4 °C. The cell pellets were resuspended in 20 mL of 0.1 M Tris-acetate buffer pH 7.5 and centrifuged at 10,000 g for 10 min at 4 °C. The cells were then resuspended in 4 mL of 0.1 M Tris-acetate buffer pH 7.5 and maintained at 0 °C for ultrasonication in an MSE Soniprep 150W ultrasonic disintegrator at a peak amplitude λ = 10 microns for 30 sec. Unbroken cells and cell wall material were removed by centrifugation at 20,000 g for 15 min at 4 °C. Cell-free extracts were prepared daily for investigation. Protein concentration was determined using a standard method (Bradford, 1976).
Dehalogenase enzyme assay and activity: Activity of the enzyme was measured by determining the release of chloride indicated by a colorimetric method employing mercuric thiocyanate as previously reported by Bergman and Sanik (1957). For the enzyme assay, dehalogenase activity was determined as total chloride released at 30 °C in an incubation mixture containing 0.1 M Tris-acetate buffer (pH 7.5) (4700 μL), 0.1 M halogenated aliphatic acid (50 μL) and distilled water and enzyme to a final volume of 5 mL. After 5 min equilibration at 30 °C, the reaction was initiated by adding cell-free extract. Samples (1.0 mL) were removed at appropriate intervals and assayed for halide ions. Each assay was carried out in triplicates. The absorbance of the mixture was measured at A460 nm and was proportional to the chloride ion concentration. Control experiments that the enzyme was stable in the incubation buffer for the time required to perform the assays. Boiled cell-free extracts did not dehalogenate MCA under the same assay conditions. Specific activity is defined as the μmole of chloride liberated per milligram protein in 10 min under the stated conditions. For kinetic studies a colorimetric method was used through out the investigation (Bergman and Sanik, 1957).
Determination of the temperature and pH optimum: For the determination of the temperature optimum cell-free extracts were incubated for 15 min at various temperature (15, 30 50 and 60 °C) and 1 mL sample were removed at 5 min interval for the enzyme assay at 30 °C as described earlier. To determine pH optimum, the cell-free extracts were mixed with equal amounts of 0.1 M Tris-acetate buffer (pH 5.0, 5.5, 6.0, 6.5 7.0, 7.5 and 8.0) of various pH. The reaction was started by addition of the substrate and after 10 min incubation chloride ions released was determined as described. Control experiment, lacking enzyme preparation was included in each set of assay to detect spontaneous halogen released.
Non-denaturing polyacrylamide gel electrophoresis (PAGE): In non-denaturing PAGE, the enzyme extract remains active. Gels were prepared based on the method of (Hardman and Slater, 1981). Resolving gels containing 12% bis-acrylamide in 375 mM Tris-acetate pH 7.8 were polymerized by the addition of 0.05% ammonium persulphate and 0.05% TEMED. Stacking gels were formed from 4% bis-acrylamide in 125 mM Tris-acetate pH 6.8. Gels were left overnight at 4 °C before being used to allow the ammonium persulphate to decompose completely. Gels were run using a Mini-Protean II gel system from Biorad in 25 mM Tris, 19 mM glycine buffer (pH 8.3) at a constant voltage of 200 V and a temperature 4 °C for 1 h. Samples were prepared as for cell extracts and mixed with 0.1 volumes of sample buffer (0.1% bromophenol blue; 10% glycerol; 100 mM DTT in 50 mM Tris-acetate pH 6.8). The gel was run until the dye front reached the bottom of the gel. Gels were then stained for dehalogenase activity.
The gel was incubated in 40 mM Cl-free halogenated substrate for 15 min at 30 °C. Substrate was carefully removed and the gel was then placed in a 0.1 M AgNO3 solution and incubated in the dark until bands appeared on the gel due to the precipitation of AgCl. The gel was then washed with distilled water to remove the AgNO3 and fixed by washing in 5% acetic acid for 10 min.
RESULTS
Isolation of MCA degrading bacteria: From paddy field agricultural soil sample, a bacteria strain R1, which degraded MCA as sole source of carbon was isolated. A single colony was observed on solid PJC minimal media supplemented with 40 mM MCA. The morphological, cultural and some physiological characteristics of the strain R1 are shown in Table 1. According to Bergey`s
| Table 1: | Some morphological and physiological properties of strain R1 |
 | |
| + : Positive, - : Negative | |
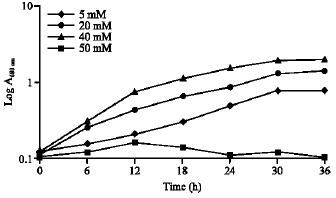 |
| Fig. 1: | Measurement of growth of Pseudomonas sp. strain R1 at various MCA concentrations. Flasks contain PJC minimal media supplemented with various MCA concentrations |
Manual of Systematic Bacteriology, the strain R1 was found to belong to the genus Pseudomonas sp. The 16S rRNA gene fragment was also sequenced using FD1 and rP1 primers. The partial nucleotide sequence of 16S rRNA gene from the isolated bacterium suggested that the species belongs to the genus Pseudomonas. This sequence was submitted to the GeneBank with accession number EU272817.
This bacterium was able to grow in the liquid PJC minimal media in the presence of MCA for concentrations ranging from 5 to 40 mM with a doubling time of 13-14 h (Fig. 1). Raising the MCA concentration above 40 mM caused a reduction in the organism`s growth rate and no further increase in cell yield suggesting that MCA in the concentration below 40 mM was non-toxic to the cell.
Figure 2 shows differential plot of the growth measured at A680 nm of Pseudomonas sp. on 40 mM MCA in a 100 mL liquid PJC minimal medium. During growth the chloride ion concentration in the medium increased until all covalently bound chloride had been released by the organism. The plot was linear during
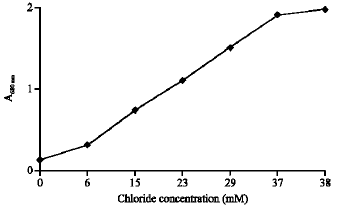 | |
| Fig. 2: | Differential plot of culture at A680 nm against chloride ions concentration for Pseudomonas sp. R1 |
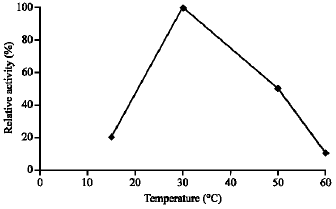 | |
| Fig. 3: | Effect of temperature on the enzyme activity |
exponential growth, indicating that the rate of MCA dehalogenation was proportional to the growth rate during this period. Since MCA was the sole carbon source in this experiment, the results also indicate that the dehalogenation product was being utilized by the organisms for metabolism and growth.
Effect of temperature and pH: Dehalogenase cell-free extracts prepare from cultures of Pseudomonas sp. grown on MCA was assayed to determine the optimum temperature and pH. The initial velocities for the first 10 min were measured with MCA at different temperature (at pH 7.5). The highest reaction rate was observed at 30 °C (Fig. 3). Dehalogenase exposed to higher temperature above 30 °C showed decreasing value of dehalogenase activity. Another important feature of dehalogenase reported here was its stability against thermal denaturation and it retained 50% of the MCA dehalogenase activity after 15 min incubation at 50 °C. Using 30 °C temperature setting, optimum pH of the dehalogenase was found to be about pH 7.5 although other pH between 6.5 to 8 showed some activity (Fig. 4).
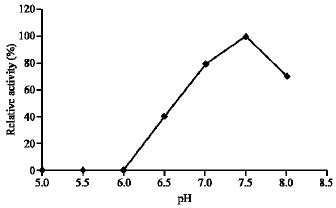 | |
| Fig. 4: | Effect of pH on the enzyme activity |
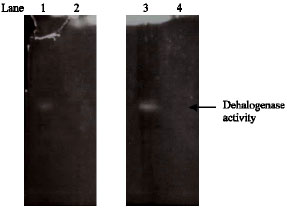 |
| Fig. 5: | Non-denaturing polyacrylamide gel electrophoresis analysis of dehalogenase activity, Lane 1: Dehalogenase activity towards MCA, Lane 2: Control: distilled water towards MCA, Lane 3: Dehalogenase activity towards MBA, Lane 4: Control: distilled water towards MBA |
Dehalogenase activity in cell-free extracts: Cell-free extracts from bacteria grown on MCA or MBA as sole source of carbon was analysed for activity towards all the growth substrates. Halide liberation was detected by extracts grown in MCA or MBA. However, no dehalogenase activity was detected in 20 h by extracts of bacteria grown in medium containing 5 mM glucose supplemented with 10 mM dichloropropionic acid or monochloropropionic acid or bi and tri-chlorinated acetic acid of dichloroacetic acid (DCA) and trichloroacetic acid (TCA), respectively. Glucose, pyruvate or glycollate alone as the carbon source did not indicate any dehalogenase activity. This experiment suggested that the dehalogenase(s) of Pseudomonas sp. R1 is inducible.
Kinetics: Apparent Km value was derived from computed Michaeles-Menten plots. The dehalogenase activity followed Michaeles-Menten kinetics over the substrate range 1 and 0.05 mM for MCA with apparent Km value of 0.14 mM.
Polyacrylamide gel electrophoresis (PAGE) of cell-free extracts: Activity stain PAGE utilizing Ag to precipitate free halide ions in regions of dehalogenase activity, has been employed to identify electrophoretically distinguishable dehalogenases. Using this technique, one single dehalogenase was identified in cell-free extracts (Fig. 5). Similar electrophoretic mobility band pattern was located when gel incubated with MBA.
DISCUSSION
A bacterial strain was isolated from soil in minimal medium supplemented with 10 mM up to 40 mM MCA. Maximum growth was obtained at 40 mM and growth above 40 mM was inhibited possibly due to toxicity of MCA. This strain with MCA dehalogenase activity was identified according to the morphological and biochemical tests shown in Table 1. The strain was designated as Pseudomonas sp. R1. Most of the previously reported dehalogenase bacterial strains were belonging to the genera Pseudomonas, Arthrobacter, Alcaligenes, Rhizobium, Rhodococcus and Methylobacterium (Hardman and Slater, 1981; Allison et al., 1983; Hardman et al., 1986; Scholtz et al., 1987; Liu et al., 1994; Ng and Huyop, 2007, 2008).
In current finding, dehalogenase activity found to be inducible. Most of the reported dehalogenating bacteria were inducible dehalogenase enzymes (Slater et al., 1979, 1997; Ng and Huyop, 2007) although constitutive dehalogenases have been reported in Moraxella sp. (Kawasaki et al., 1981). The optimal pH was also investigated in the crude cell-free extracts. However, some reported dehalogenase activities were within pH 6.5 and 8.0 with the activity decreasing markedly on either side of these values. pH profiles of dehalogenase activities in several strains of Pseudomonas (Smith et al., 1990), Arthrobacter (Scholtz et al., 1987) and Moraxella (Kawasaki et al., 1981) described pH optima in the same range as the values reported here. No acidic dehalogenases activity has so far been reported. In biotechnology, a potential enzyme in industry requires stability at higher temperature. However, thermal stability of our enzyme remained 50% active when exposed to 50 °C.
The affinity of dehalogenases for their substrates is of biotechnological interest especially degradation of halogenated compound at lower substrate concentration. Biodegradation of very low concentrations of xenobiotic compounds has been neglected. The kinetic study obtained for the MCA dehalogenase activity at their optimal pH was 0.14 mM. Km range for different halogenated compound has been reported for other Pseudomonas isolate (Tsang et al., 1988).
Activity staining of native PAGE has been used in order to identify whether a single or more dehalogenases were present. Multiple dehalogenases are separable as a result of relative electrophoretic mobilities (Allison et al., 1983; Keuning et al., 1985). The visualization of dehalogenase activity involved a single chloro and bromo atom of MCA and MBA as the substrates. Other compounds such as DCA and TCA was not tested. On non-denaturing gel electrophoresis the activity appears to be due to a single protein. Using the nomenclature based on electrophoretic mobility, Pseudomonas cepacia MBA4 possesses more than one dehalogenases with dehalogenase IVa which is expressed in batch and continuous culture and dehalogenase III, which is only observed in cell-free extracts prepared from cells grown in continuous culture (Tsang et al., 1988). Rhizobium sp. was also reported to contain multiple dehalogenases based on similar investigation (Leigh, 1986).
The identification of so many dehalogenases and the presence of multiple dehalogenases in many genera and species so far raise the question of their environmental significance. These enzymes are specifically induced by halo-acetic and propionic acids involving complex regulatory controls which respond to the growth environment. Some dehalogenases are specific and react to carbon-halogen bonds adjacent to the carboxyl group, whether at α and β-position of halogen group. Are these dehalogenases in natural environment will be expressed as dehalogenases or less specific hydrolases? The study of a complex regulatory control which responds to the growth environment may shed light on this question.
ACKNOWLEDGMENTS
This study was funded by a Ministry of Science, Technology and Innovation of Malaysia under Vot numbers 79073 (MOSTI e-Science Fund) and 74190 (RMC-IRPA RM-8 Research Grant) and both sources of funding are gratefully acknowledged by SNI, AMT and NHJ.
REFERENCES
- Allison, N., A.J. Skinner and R.A. Cooper, 1983. The dehalogenases of a 2,2 dichloropropionate degrading bacterium. J. Gen. Microbiol., 129: 1283-1293.
CrossRef - Bergman, J.G. and J. Sanik, 1957. Determination of trace amounts of chlorine in naptha. Anal. Chem., 29: 241-243.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Hardman, D.J. and J.H. Slater, 1981. Dehalogenases in soil bacteria. J. Gen. Microbiol., 123: 117-128.
Direct Link - Hareland, W.A., R.L. Crawford, P.J. Chapman and S. Dagley, 1975. Metabolic function and properties of a 4-hydroxyphenylacetic acid 1-hydroxylase from Pseudomonas acidovorans. J. Bacteriol., 121: 272-285.
Direct Link - Holt, J.G., N.R. Kreig, P.H.A. Sneath, J.T. Staley and S.T. Williams, 1994. Bergey's Manual of Determinative Bacteriology. 9th Edn., Lippincott Williams and Wilkins, Baltimore, USA., ISBN-13: 9780683006032, Pages: 787.
Direct Link - Sui-Yi, K., A.F.M. Siu, S.M. Ngai, C.M. Che and J.S.H. Tsang, 2007. Proteomic analysis Burkholderia cepacia MBA4 in the degradation of monochloroacetate. Proteomics, 7: 1107-1116.
CrossRefDirect Link - Jing, N.H. and F. Huyop, 2007. Dehalogenation of chlorinated aliphatic acid by Rhodococcus sp. Asia Pac. J. Mol. Biol. Biotechnol., 15: 147-151.
Direct Link - Jing, N.H. and F. Huyop, 2008. Enzymatic dehalogenation of 2,2-dichloropropionic acid by locally isolated Methylobacterium sp. HJ1. J. Biol. Sci., 8: 233-235.
CrossRefDirect Link - Olaniran, A.O., G.O. Babalola and A.I. Okoh, 2001. Aerobic dehalogenation potentials of four bacterial species isolated from soil and sewage sludge. Chemosphere, 45: 45-50.
CrossRef - Olaniran, A.O., D. Pillay and B. Pillay, 2004. Haloalkane and haloacid dehalogenases from aerobic bacterial isolates indigenous to contaminated sites in Africa demonstrate diverse substrate specificities. Chemosphere, 55: 27-33.
CrossRef - Scholtz, R., T. Leisinger, F. Suter and A.M. Cook, 1987. Characterization of 1-chlorohexane halidohydrolase, a dehalogenase of wide substrate range from an Arthrobacter sp. J. Bacteriol., 169: 5016-5021.
Direct Link - Slater, J.H., D. Lovatt, A.J. Weightman, E. Senior and A.T. Bull, 1979. The growth of Pseudomonas putida on chlorinated aliphatic acids and its dehalogenase activity. J. Gen. Microbiol., 114: 125-136.
CrossRef - Smith, J.M., K. Harrison and J. Colby, 1990. Purification and characterization of D-2-haloacid dehalogenase from Pseudomonas putida strain AJ1/23. J. Gen. Microbiol., 136: 881-886.
PubMed - Tsang, J.S.H., P.J. Sallis, A.T. Bull and D.J. Hardman, 1988. A monobromoacetate dehalogenase from Pseudomonas cepacia MBA4. Arch. Microbiol., 150: 441-446.
CrossRef