
Emma W. Gachomo
Institute for Plant Diseases, University of Bonn, Nussallee 9, D-53115 Bonn, Germany
Simeon O. Kotchoni
Department of Agronomy, Purdue University, Lilly Hall, 915 West State Street, West Lafayette, IN 47907, USA
Biotechnology
Year: 2008 | Volume: 7 | Issue: 2 | Page No.: 273-279







ABSTRACT
This study was conducted with the aim of assessing the effect of plant extract derived from drought stressed rose leaves in controlling black spot disease of roses caused by Diplocarpon rosae infection. The plant extract is shown here to contain elevated levels of various reactive molecules such as H2O2, O2-, nitric oxide (NO), malondialdehyde (MDA) and expression of pathogenesis related proteins mediating defence responses in plants. Treatment of rose plants with the extract promotes black spot disease resistance through enhanced expression of pathogenesis related proteins, peroxidase and accumulation of defence response elicitors. These data suggest that plant extract from drought-stress leaves can be used as a promising alternative to control plant diseases in an environmentally friendly way.
PDF Abstract XML References Citation
How to cite this article
Emma W. Gachomo and Simeon O. Kotchoni, 2008. Extract from Drought-Stress Leaves Enhances Disease Resistance Through Induction of Pathogenesis Related Proteins and Accumulation of Reactive Molecules. Biotechnology, 7: 273-279.
DOI: 10.3923/biotech.2008.273.279
URL: https://scialert.net/abstract/?doi=biotech.2008.273.279
DOI: 10.3923/biotech.2008.273.279
URL: https://scialert.net/abstract/?doi=biotech.2008.273.279
INTRODUCTION
The rose has established itself as the most important commercial cut flower grown under glass (Horst, 1983). It is generally used in gardens and landscaping for its aesthetic value. However, the cultivation of roses is associated with various diseases. The most common rose diseases are: powdery mildew caused by Sphaerotheca pannosa, black spot disease by Diplocarpon rosae and bacterial wilt caused by Agrobacterium tumefaciens (Horst, 1983). Black spot of roses is a devastating and widespread fungal disease. It is caused by ascomycetous fungus Marssonina rosae with the sexual state known as Diplocarpon rosae Wolf (Wolf, 1912; Gachomo and Kotchoni, 2007). The disease is characterized by black spots, yellowing and premature defoliation of the leaves (Gachomo, 2005; Gachomo et al., 2006). Although, the development of resistant cultivars against black spot disease is underway (Debener, 2003), the control of black spot still requires the use of systemic fungicides (Walker et al., 1995; Reddy et al., 1992), which is not desirable due to environmental concerns and the high cost of fungicides. It is therefore imperative to develop other efficient and safe strategies to control black spot disease.
Plants in their natural environment are often challenged by both abiotic and biotic stresses. They respond to these environmental stresses through various defense mechanisms including accumulation of Reactive Oxygen Species (ROS), nitric oxide (NO), induction of secondary metabolites (phytoalexins) and production of plant cell wall derived metabolites such as oligogalacturonides (Dixon and Harrison, 1990; Reymond et al., 1995; Harborne, 1999). Recent investigations have proved the involvement of H2O2 mediated alteration of phytohormones in plant defense mechanisms (Dat et al., 2000; Kotchoni and Gachomo, 2006). Exogenous application of H2O2 to roses has been shown to promote black spot disease resistance (Gachomo and Kotchoni, 2006). Oligogalacturonidases, a product of cell wall degradation, stimulate in vitro phosphorylation of leaf plasma membrane-associated proteins required in tomato and potato defense responses (Farmer et al., 1989, 1991; Reymond et al., 1995). This indicates that components of cell wall derived metabolites might promote disease resistance acquisition during plant-pathogen interactions. Linolenic acid hydroperoxidation (a product of lipid peroxidation) is another intermediate found to induce the expression of proteinase inhibitors involved in defense responses in tomato leaves (Farmer et al., 1994).
Abiotic stresses such as drought, UV-light and chemical treatment are found to induce elevated accumulation of ROS (Kotchoni et al., 2006), phytoalexins (Gachomo et al., 2003), lignin, phenolic compounds, cellulose, callose and cell wall proteins (Harborne, 1999; Kotchoni et al., 2006). We therefore hypothesized that chemical component of plant extracts derived from drought stressed plants might contain readily available and well-balanced metabolites that can efficiently control plant disease in an environmental friendly manner. Using rose leaf extract derived from drought stressed plants we here examined the possibility of efficiently controlling black spot disease by spraying rose plants before and after pathogen infection. The induction of reactive substances and pathogenesis related proteins and their role in conferring disease resistance in plant host cells are here discussed.
MATERIALS AND METHODS
Plant material, growth conditions and fungal inoculum: Floribunda rose plants cv. Frensham of parentage [Floribunda seedling x crimson Glory] obtained from Belle Epoque Rozenwekerij breeders (Netherlands) were used in this study. Pure culture of Diplocarpon rosae used for in vitro infection was generously provided by Dr. Gachomo (fungal stock cultures; Gachomo, 2005). The inoculum concentration was adjusted to 1x105 conidia mL–1. Potted plants infected with the pathogen were maintained in greenhouse at 20±5°C, 300 μExm–2, with a day/night cycle of 14/10 h and a relative humidity of 48±5% and leaf tissue samples were taken at pre-determined time intervals post inoculation. The untreated control plants were inoculated with sterile distilled water. The disease severity on infected plants was scored at 10, 15 and 20 days post inoculation (dpi) based on the diameter of the black spots, yellowing and defoliation according to the following scale: 0%-no disease symptom; 1-20%-disease spots less than 5 mm, no yellowing and no defoliation; 21-40% - spots less than 5 mm, yellowing and/or defoliation; 41-60% - spots of 5-10 mm, no yellowing and no defoliation; 61-80% - spots of 5-10 mm, yellowing and/or defoliation; 81-100% - spots of a diameter greater than 10 mm, yellowing of leaves and/or defoliation (Gachomo, 2005; Gachomo et al., 2006).
Plant extract derived from dehydrated rose plants: Healthy, non-infected rose plants were subjected to drought stress in the greenhouse by withholding watering for two weeks. Leaf tissues of the stressed plants were collected, ground in liquid nitrogen and resuspended in sodium phosphate buffer pH 7.0. The suspension was filtered through a double-layered cheesecloth. Tween 20 [0.1% (v/v)] was added to the suspension before use. The suspension was sprayed on the test plants (both infected and non infected plants) not later than one hour after extraction throughout this study to avoid alteration of reactive molecules of the plant extract.
In order to assess the involvement of ion channel pathways in the plant defense mechanism, another set of plants were treated with drought stressed plant extract containing 0.5 mM EDTA (ion chelator) and compared to the EDTA-free extract treated plants at 15 dpi.
Protein extraction and detection of peroxidases (PODs): Total protein was extracted from 200 mg of fresh weight plant material by boiling ground plant material in 1 x SDS loading buffer [62.5 mM Tris-HCl, pH 6.8, 10% (v/v) glycerol, 2% (w/v) SDS, 5% β-mercaptoethanol and 0.1% (w/v) bromophenolblue] (Laemmli, 1970). Equal amounts of protein (20 μg) were separated in 12% (w/v) SDS-PAGE (Zehr et al., 1989) and stained with 20 mM Guiacol + 0.1% (v/v) H2O2 (Sigma, Steinheim, Germany) to detect the accumulation of peroxidase proteins (PODs) according to Fecht-Christoffers et al. (2003). Pre-stained protein molecular weight markers (Sigma-Saint Louis, USA) were used as molecular weight standard for the protein analysis.
Histochemical staining and quantification of H2O2 levels: To visualize H2O2 in the plants, excised leaves were placed in 1 mg mL–1 3,3`-diaminobenzidine (DAB) solution (Sigma, Steinheim, Germany) and incubated at 25°C in dark, for 8 h at 250 rpm. The chlorophyll of the leaf samples was then cleared at 80°C in 80% (v/v) ethanol for 2 h and the cleared leaves embedded in 10% (v/v) glycerol for observations (Thordal-Christensen et al., 1997). DAB was dissolved at pH 5.4 and adjusted to pH 7.5. The quantification of H2O2 was performed according to Tiedemann (1997). H2O2 concentrations were obtained via standard calibration with solutions containing 5, 10, 25, 50, 75, 100, 150 μM H2O2 (Sigma, Steinheim, Germany) as described by Kotchoni et al. (2006).
Detection of superoxide (O2–) using NBT: The superoxide ion was assayed using a modified nitroblue tetrazolium (NBT) reduction test (Doke, 1983). Leaves were harvested from plants at various times following drought stress. Leaves from none-stressed plants were used in the control. The leaves were placed in the reaction solution (10 mM potassium phosphate buffer [pH 7.8], 0.05% (w/v) NBT and 10 mM NaN3) for 1 h at room temperature, 20±5°C. The reaction was stopped by heating the tubes at 85°C for 15 min and then cooling immediately in ice. The reduction of NBT in presence of O2– to an insoluble blue product, formazan, was quantified using AIDA Image Analyzer version 2.11 (Raytest GmbH, Straubenstadt, Germany).
To confirm the role of superoxide anions in the reduction of NBT, 100 units of active superoxide dismutase (SOD; Sigma, Steinheim, Germany) in 10 mM potassium phosphate buffer (pH 7.5) were added to a set of tubes containing leaves harvested at similar intervals from drought stress. Inactive SOD (prepared by boiling for 10 min) was used for another set of tubes as control.
Lipid peroxidation assay: The levels of lipid peroxidation in the plants were assayed using the thiobarbituric acid (TBA) test, which determines the amounts of malondialdehyde (MDA) as end product of the lipid peroxidation according to Kotchoni et al. (2006).
Nitric oxide (NO) detection using the DAF2-DA fluorescence: Plant tissues were incubated in the dark in 10 μM of 4,5-diaminofluoresceindiacetate (DAF2-DA) (Sigma, Steinheim, Germany) dissolved in 10 mM Tris-KCl, pH 7.2 for 10 h. Leaf areas were then analysed microscopically using an Axioplan Universal fluorescence microscope equipped with a confocal laser scanner (Zeiss, Goettingen, Germany). DAF2-DA fluorescence was recorded using a channel with a 505 to 530 nm band-pass filter and the detected NO was quantified using AIDA Image Analyzer version 2.11 (Raytest GmbH, Straubenstadt, Germany).
Statistical analysis: Data were expressed as mean values (±SD) of at least three independent experiments. p-values were determined by Student`s t-test analysis.
RESULTS AND DISCUSSION
Plant extract treatment inhibits fungal infection and confers resistance to D. rosae: The ultimate goal of this study was to evaluate the potential of drought-stressed plant extract in controlling black spot disease of roses. We took the advantage of our previous study in which H2O2 treatment conferred resistance to black spot disease in roses (Gachomo and Kotchoni, 2006) to hypothesize that multiple reactive molecules in drought-stressed plant extract might be a reservoir of a well-balanced regulatory component network to confer disease resistance. The disease symptoms and severity on the extract-treated plants was compared to the control samples sprayed with non-stressed plant extract or water. Results of non-stressed extract sprayed samples are similar to the water sprayed samples. As shown in Fig. 1a, the disease symptoms were highly reduced in the samples treated Disease symptoms of leaves treated with non stressed (NS) and drought stressed (DS) extract at 15 dpi are shown (a); disease severity of plants treated with non stressed (NS) and drought stressed (DS) extract at different days post inoculation was shown (b). Data represent mean values (±SD) of three independent experiments with drought-stressed extract compared to the control. The disease symptoms established and spread faster in the control plants compared to those treated with the drought-stressed extract (Fig. 1a). Likewise, the disease severity examined at different time post inoculation was significantly reduced in the plants sprayed with drought stressed extract compared to the control (Fig. 1b). This suggests that components of drought stressed plant extract positively regulate defense responses and protection of rose plants against D. rosae infection. It is known that ion transport pathways are involved in the processes of plant defense responses (Stamler, 1994; Leshem, 1996; Noritake et al., 1996; Xu et al., 1998). We therefore investigated whether the plant extract treatment mediates defense responses through ion transport pathways. To do so we chelated the ions in the stressed plant extract by adding EDTA to the extract. EDTA, an ion chelator, compromises the function of ion channel flux pathways (Kotchoni and Shonukan, 2002). Addition of 0.5 mM EDTA led to reduced-disease resistance in plants treated with the extract (Table 1), demonstrating therefore the functional implication of ions, Ca2+, in mediating defense response in plants sprayed with the stressed extract. The addition of Ca2+ (CaCl2, 150 mM end concentration) reversed the effect of EDTA and restored the ability of plant stressed extract to trigger disease resistance in infected leaves (Table 1).
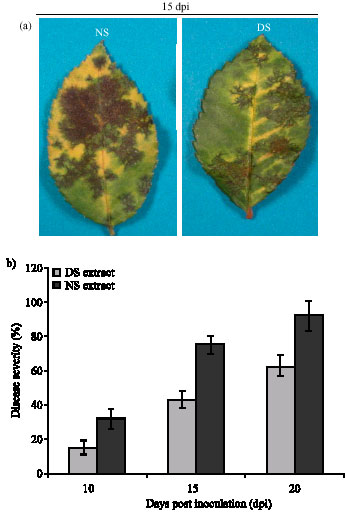 | |
| Fig. 1: | Induction of disease resistance in rose leaves treated with drought stressed extract. |
| Table 1: | Disease severity in infected plant following drought-stress plant extract treatment |
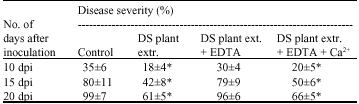 | |
| Data represent the mean percentage values (±SD) of three replicate experiments. The mean differences (±SD) marked with * are statistically significant at p<0.05, (n = 15) according to Student`s t-test. DS plant extr. = Drought-stress plant extract | |
| Table 2: | Drought stress promotes accumulation of different reactive molecules in plant extract |
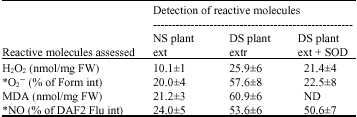 | |
| *: The intensity of detected substances was quantified using AIDA image analyzer version 2.11 and expressed in % of background intensity referred to as 1%. NS plant ext = non-stress plant extract, DS plant ext = drought-stress plant extract, Form Int = formazan intensity, Flu Int = fluorescence intensity, FW = Fresh Weight. ND = Not Determined, SOD = Superoxide Dismutase. Data represent the mean percentage values (±SD) of three replicate experiments | |
Metabolite composition of the plant extract: Various metabolites and reactive molecules such as H2O2, O2– and NO were detected in rose leaves subjected to drought stress (Fig. 2a-e, Table 2). Addition of active SOD significantly inhibited reduction of NBT by drought stressed plants, while inactive SOD had no effect. SOD is an O2– scavenger and its presence significantly reduced the amount of O2– in the plant extract (Table 2) thereby having significant inhibitory effects on the reduction of NBT compared to the control. Results of previous investigations indicated that exogenous application of H2O2 significantly inhibited degradation of plant host proteins by the pathogen, improving disease resistance of infected rose leaves (Gachomo and Kotchoni, 2006). Reactive Oxygen Species (ROS) such as O2– or H2O2 have been shown by virtue of their chemical properties to mimic the oxidative burst generated during the early stages of the Hypersensitivity Responses (HR), which mediate the induction of defense responses in plants (Runeckles and Vaartnou, 1997; Rao and Davis, 1999; Malolepsza, 2005). In addition, the level of MDA derived from lipid peroxidation as result of drought stress was higher in drought extract compared to control (Table 2). Putting these observations together, ROS molecules together with other reactive molecules are believed to trigger signal transduction through ion channel pathways to promote improved defense responses in extract treated rose plants against D. rosae infection.
 | |
| Fig. 2: | Induction of reactive molecules in rose leaves by drought stress. Pictures of leaf before (a) and after two weeks of drought stress (b) are here shown. The accumulation of H2O2 in the leaves before (c) and after (d) drought stress was detected by DAB staining. The content of O2– (e) was examined in the leaf extract at different exposure times to drought stress using nitroblue tetrazolium (NBT) buffer reagent. In presence of O2–, NBT is reduced to a blue coloured product, which positively correlates with the amount of O2– present in the solution |
The typical preformed and constitutive defense barriers are generally structural barriers (waxes, cutin, suberin, lignin, phenolics, cellulose, callose and cell-wall proteins), which are subsequently reinforced upon the infection. Defense reinforcement mechanisms include oxidative burst (ROS), nitric oxide (NO) accumulation, rapid and localized cell death (hypersensitive response), accumulation of phytoalexins and synthesis of Pathogenesis-Related (PR) proteins. These defense mechanisms are not only activated upon infection by pathogenic microorganisms, but also induced by abiotic stresses such as UV-light, chemical treatment, drought and salt stress (Harborne, 1999; Kotchoni et al., 2006). Therefore, we hypothesized that the extract from drought stressed plants might contain defense related metabolic compounds i.e. phytohormones, reactive molecules and readily active proteins/enzymes to fight off pathogen infections.
In summary, drought stress promoted the accumulation of various molecules including by-products of lipid peroxidation that could probably prime the stress extract-treated plants to efficiently fight off the pathogen infections. Since we detected higher accumulation of various reactive metabolites in the plant extract, we investigated whether spraying rose plants with the stressed plant extract would induce the accumulation of these metabolites in rose plants following spraying.
Induction of metabolites in roses after spraying with drought-stress extract: Since the stressed extract contained elevated levels of various defense related molecules/metabolites and was able to reduce disease severity in infected roses (Table 2, Fig. 1), we then predicted that plants treated with such extract might show alteration in physiological metabolism leading to accumulation of phytohormones or reactive molecules mediating defense responses.
Plant samples treated with the drought-stressed extract showed elevated accumulation of ROS (H2O2, O2–) (Table 3), although not too high to induce oxidative damage. Another important reactive molecule involved in plant defense responses is nitric oxide (NO). As shown in Table 3, higher accumulation of NO was detected in plants sprayed with drought-stress plant extract compared to control. The detection of O2–, H2O2 and other reactive substances in the stressed extract and the increased levels of these molecules after plant treatment indicate their crucial involvement in rose defense response pathways against D. rosae infection (Table 2). Research evidence has shown that NO often acts as a plant defense response mediator through the activation of guanylate cyclase, which produces the second messenger cyclic GMP (cGMP) and S-nitrosylation of redox-sensitive transcription factors, or through ion (Ca2+) channels (Stamler, 1994; Xu et al., 1998). The ion channel pathway is crucial for activation of defense response because the alteration of ion flow through EDTA-mediated ion chelation led to loss of disease resistance induction by the plant extract (Table 1). The ions and ion channel pathways are linked to phytoalexin production pathways (Leshem, 1996; Noritake et al., 1996) and act as a redox transmitters in the regulation of an array of diverse physiological processes, especially in host responses to pathogen infection (Stamler, 1994; Hausladen et al., 1996). Treatment of the drought-stressed extract with EDTA inhibits the induction of NO accumulation in the sprayed rose samples, while the addition of Ca2+ was able to reverse the NO inhibitory effect of EDTA (Table 1, 3). In the plants treated with the extract, the elevated levels of ROS and NO were therefore thought to additionally promote defense response through the involvement of ion flux in well-coordinated network pathways resulting in the expression of pathogenesis-related genes. To confirm this hypothesis, we checked the expression pattern of pathogenesis related proteins in infected plants treated with the stressed extract. As shown in Fig. 3, the exogenous application of the drought-stress extract induced the expression of various pathogenesis related proteins, in this case the peroxidases (POD). POD has several physiological functions in plant tissue; its activation in response to a broad range of biotic and abiotic factors plays an important role in defense responses (Greppin et al., 1986; Obinger et al., 1996). POD activity is often used as a physiological marker for plant stress response. It is here confirmed that accumulation of PODs mediated by drought-stress extract treatment are important for plant response to D. rosae infection. In total, our data suggest that spraying plants with drought-stress plant extract promotes disease resistance through alteration of metabolites leading to elevated expression of pathogenesis related proteins (PODs).
| Table 3: | Drought-stress plant extract mediates disease resistance through ion channel network and accumulation of reactive molecules |
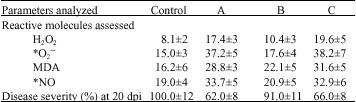 | |
| The values are expressed in the same unit as described in Table 2. * See Table 2 for detail information on O2– and NO quantification assay. Data represent the mean percentage values (±SD) of three replicate experiments. DS ext = drought-stress extract, (A) = drought-stress extract treated plant, (B) = drought-stress extract plant + EDTA. (C) = drought-stress extract treated plant + EDTA + Ca2+ | |
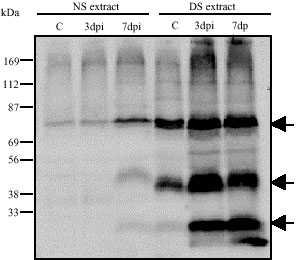 | |
| Fig. 3: | One-dimensional (1D) pathogenesis related protein analysis. Proteins extracted from control (C, non infected plant samples) and infected plants treated with non Stressed (NS) and Drought Stressed (DS) extract were separated in SDS-PAGE. Peroxidases (PODs) were detected (see arrows) by staining with 20 mM Guaiacol, 0.01% (v/v) H2O2 (cf. Material and Methods) |
The results presented here indicate that disease resistance could be obtained by spraying plants with drought-stress extract. This approach of disease management is harmless to the environment. The use of abiotic-stress plant extract in disease control may reduce or positively complement some of the fungicide factors and minimizes exposure of farmers to fungicides. Compared to fungicides, which often have only one specific active ingredient against the pathogen(s), DS-plant extract contains a versatile and highly dynamic chemical composition, which directly depends on the drought stress conditions and therefore very difficult for the pathogen(s) to develop resistance mechanisms against. Another interesting feature is the high flexibility of the approach e.g. one could decide to increase the number of sprays without any adverse effect on the environment. This approach may be used to control different pathogens. This will not only be advantageous to the farmer`s health, but also that of the consumers.
REFERENCES
- Dat, J., S. Vandenbeele, E. Vranova, M. Van Montagu, D. Inze and F. Van Breusegem, 2000. Dual action of the active oxygen species during plant stress responses. Cell. Mol. Life Sci., 57: 779-795.
CrossRef - Dixon, R.A. and M. Harrison, 1990. Activation, structure and organization of genes involved in microbial defense in plants. Adv. Genet., 28: 165-234.
PubMed - Doke, N., 1983. Involvement of superoxide anion generation in the hypersensitive response of potato tuber tissues to infection with an incompatible race of Phytophthora infestans and to the hyphal wall components. Physiol. Plant Pathol., 23: 345-357.
CrossRefDirect Link - Farmer, E.E., G. Pearce and C.A. Ryan, 1989. In vitro phosphorylation of plant plasma membrane proteins in response to the proteinase inhibitor inducing factor. Proc. Natl. Acad. Sci. USA., 86: 1539-1542.
Direct Link - Farmer, E.E., T.D. Moloshok, M.J. Saxton and C.A. Ryan, 1991. Oligosaccharide signaling in plants. Specificity of oligouronide-enhanced plasma membrane protein phosphorylation. J. Biol. Chem., 266: 3140-3145.
Direct Link - Farmer, E.E., D. Caldelari, G. Pearce, M.K. Walker-Simmons and C.A. Ryan, 1994. Diethyldithiocarbamic acid inhibits the octadecanoid signaling pathway for the wound induction of proteinase inhibitors in tomato leaves. Plant Physiol., 106: 337-342.
CrossRefDirect Link - Fecht-Christoffers, M.M., H.P. Braun, C. Lemaitre-Guillier, A. van Dorsselaer and W.J. Horst, 2003. Effect of manganese toxicity on the proteome of the leaf apoplast in cowpea. Plant Physiol., 133: 1935-1946.
Direct Link - Gachomo, E.W., O.O. Shonukan and O.S. Kotchoni, 2003. The molecular initiation and subsequent acquisition of disease resistance in plants. Afr. J. Biotechnol., 2: 26-32.
Direct Link - Gachomo, E.W., 2005. Studies of the Life Cycle of Diplocarpon rosae Wolf on Roses and the Effectiveness of Fungicides on Pathogenesis. Cuvillier Verlag, Goettingen, pp: 1-147.
Direct Link - Gachomo E.W., W.H. Dehne and U. Steiner, 2006. Microscopic evidence for the hemibiotrophic nature of Diplocarpon rosae, cause of black spot disease of rose. Physiol. Mol. Plant Pathol., 69: 86-92.
CrossRef - Gachomo, E.W. and O.S. Kotchoni, 2007. . Detailed description of developmental growth stages of Diplocarpon rosae wolf in Rosa: A building block for an efficient disease management. Ann. Applied Biol., 151: 233-243.
Direct Link - Harborne, J.B., 1999. The comparative biochemistry of phytoalexin induction in plants. Biochem. Syst. Ecol., 27: 335-367.
CrossRefDirect Link - Hausladen, A., C.T. Privalle, T. Keng, J. DeAngelo and S.J. Stamler, 1996. Nitrosative stress: Activation of the transcription factor OxyR. Cell, 86: 719-729.
CrossRefDirect Link - Kotchoni, O.S. and O.O. Shonukan, 2002. Regulatory mutations affecting the synthesis of cellulase in Bacillus pumilus. World J. Microbiol. Biotechnol., 18: 487-491.
CrossRefDirect Link - Kotchoni, O.S. and W.E. Gachomo, 2006. The reactive species network pathways: An essential prerequisite for perception of pathogen attack and the acquired disease resistance in plants. J. Biosci., 31: 389-404.
Direct Link - Kotchoni, O.S., C. Kuhns, A. Ditzer, H.H. Kirch and Bartels, 2006. Over-expression of different aldehyde dehydrogenase genes in Arabidopsis thaliana confers tolerance to abiotic stress and protects plants against lipid peroxidation and oxidative stress. Plant Cell Environ., 29: 1033-1048.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Leshem, Y.Y., 1996. Nitric oxide in biological systems. Plant Growth Regul., 18: 155-159.
CrossRefDirect Link - Malolepsza, U., 2005. Spatial and temporal variation of reactive oxygen species and antioxidant enzymes in o-hydroxyethylorutin-treated tomato leaves inoculated with Botrytis cinerea. Plant Pathol., 54: 317-324.
Direct Link - Noritake, T., K. Kawakita and N. Doke, 1996. Nitric oxide induces phytoalexin accumulation in potato tuber tissues. Plant Cell Physiol., 37: 113-116.
CrossRefDirect Link - Rao, M.V. and K.R. Davis, 1999. Ozone-induced cell death occurs via two distinct mechanisms in Arabidopsis: The role of salicylic acid. Plant J., 17: 603-614.
PubMed - Reddy, S., J.A. Spencer and S.E. Newman, 1992. Leaflet surfaces of blackspot-resistant and susceptible roses and their reactions to fungal invasion. HortScience, 27: 133-135.
Direct Link - Reymond, P., S. Grunberger, K. Paul, M. Muller and E.E. Farmer, 1995. Oligogalacturonide defense signals in plants: Large fragments interact with the plasma membrane in vitro. Proc. Natl. Acad. Sci. USA., 92: 4145-4149.
Direct Link - Runeckles, V.C. and M. Vaartnou, 1997. EPR evidence for superoxide anion formation in leaves during exposure to low levels of ozone. Plant Cell Environ., 20: 306-314.
CrossRefDirect Link - Stamler, J.S., 1994. Redox signaling: Nitrosylation and related target interactions of nitric oxide. Cell, 78: 931-936.
PubMed - Tiedemann, A.V., 1997. Evidence for a primary role of active oxygen species in induction of host cell death during infection of bean leaves with Botrytis cinerea. Physiol. Mol. Plant Pathol., 50: 151-166.
CrossRefDirect Link - Walker, S., Z. Mandegaran and A.M. Roberts, 1995. Screening roses for resistance to Diplocarpon rosae. Acta Hort., 424: 209-213.
CrossRef - Xu, L., J.P. Eu, G. Meissner and J.S. Stamler, 1998. Activation of the cardiac calcium release channel (ryanodine receptor) by poly-S-nitrosylation. Science, 279: 234-237.
CrossRefDirect Link - Zehr, B.D., T.J. Savin and R.E. Hall, 1989. A one-step, low background coomassie staining procedure for polyacrylamide gels. Anal. Biochem., 182: 157-159.
CrossRefDirect Link - Thordal-Christensen, H., Z. Zhang, Y. Wei and D.B. Collinge, 1997. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley-powdery mildew interaction. Plant J., 11: 1187-1194.
CrossRefDirect Link