
Monika Garg
Faculty of Agriculture, Tottori University, 4-101 Koyama cho Minami, Tottori, 680-0945, Japan
Y.S. Rao
Department of Plant Breeding, Genetics and Biotechnology, Punjab Agricultural University, Ludhiana, Punjab, India
Ajay Goyal
Faculty of Agriculture, Tottori University, 4-101 Koyama cho Minami, Tottori, 680-0945, Japan
Baldev Singh
Department of Plant Breeding, Genetics and Biotechnology, Punjab Agricultural University, Ludhiana, Punjab, India
Biotechnology
Year: 2007 | Volume: 6 | Issue: 3 | Page No.: 444-446







ABSTRACT
Twenty one accessions from different diploid species, 10 high yielding varieties each of Triticum durum and Triticum aestivum were analysed by SDS-PAGE, to study polymorphism among the triticin protein. Also tricin gene fragment, of 21 accessions of different diploid species was amplified by PCR. No polymorphism was found in triticin protein subunits of T. durum and T. aestivum varieties but enough polymorphism was observed in wild wheats. The A-genome species -T. urartu and T. boeticum accessions having molecular weight higher than 1AS coded triticin subunits of Chinese spring can be used for further analysis. Expression of triticin subunits, in S-genome, Ae. longissima and Ae. speltoides and absence of the same in hexaploid wheats (B-gemone coded subunits) suggested that later can be transferred to cultivated wheats for increasing triticin/lysine content.
PDF Abstract XML References Citation
How to cite this article
Monika Garg, Y.S. Rao, Ajay Goyal and Baldev Singh, 2007. Variations in Seed Storage Protein-Triticin among Diploid Triticum and Aegilops Species. Biotechnology, 6: 444-446.
DOI: 10.3923/biotech.2007.444.446
URL: https://scialert.net/abstract/?doi=biotech.2007.444.446
DOI: 10.3923/biotech.2007.444.446
URL: https://scialert.net/abstract/?doi=biotech.2007.444.446
INTRODUCTION
Wheat (Triticum aestivum) is most widely consumed food crop in the world. Wheat proteins are considered to be poorly balanced in terms of all essential amino acids because of the limiting amino acids lysine and threonine (Payne, 1983). Increasing the proportion of lysine in wheat protein would substantially increase its nutritional value, as monogastric animals do not have the ability to synthesize lysine, not even from its α-keto acid precursor.
Triticin, a minor storage protein of wheat, the gene of which is located on short arm of chrosomes 1A, 1B and 1D (Singh et al., 1993) is rich in lysine content. The cDNA sequence of triticin has shown lysine-rich decapeptide motif in the hypervariable region (HVR) of the protein.
Thus it may be possible to increase the lysine content of wheat seed by development of transgenic plants, by using more efficient promoter and/or further increasing the lysine content of triticin in its HVR region through genetic engineering. However, the acceptability of transgenic food is still in question, therefore searching the germplasm of wheat and its wild relatives, that could express enhanced level of lysine content may prove to be better alternative. The present experiment was designed to investigate the polymorphism of triticin protein and its gene in cultivated and wild Triticum and Aegilops species.
MATERIALS AND METHODS
The plant material used for this study was 21 accessions from different diploid species (Table 1), 10 high yielding varieties each of Triticum durum (AB) and Triticum aestivum (ABD). All the material was obtained from Biotechnology center, Punjab Agricultural University, Ludhiana, Punjab-India.
Triticin extraction and Sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) was carried out according to Shailaja et al. (2002).
DNA was extracted by SDS-potassium acetate method (Dellaporta et al., 1983). PCR reactions were set using C2/T13 primer (Shailaja et al., 2002). This primer amplifies 816bp fragment including HVR region and a segment between 2 conserved cysteine residues involved in interchain disulphide linkage.
| Table 1: | Accessions of various diploid Triticum and Aegilops species used for polymorphism of triticin using SDS-PAGE |
 | |
RESULTS AND DISCUSSION
The genes of triticin are located on the short arm of 1A, 1B and 1D, however the expression of 1B encoded protein subunits have not been detected so far (Singh et al., 1993). Banding pattern of unreduced triticin extracts from Chinese spring on SDS-PAGE shows triplet band, the slowest moving triticin band is coded by gene on the short arm of chromosome 1D, the fastest moving on the short arm of chromosome 1A, whereas the middle is a hybrid product of the two proteins.
The variations in triticin patterns of different diploid Triticum and Aegilops species are shown in Fig. 1. It showed that there was enough polymorphism for triticin protein among the accessions of T. beoticum, T. urartu, Ae. longissima and Ae. speltoides, but no polymorphism among the accessions of Ae. Squarrosa. The triticin subunits of T. beoticum were distinct in having higher molecular weight, as compared to those encoded by T. urartu, the A genome donar to polyploid wheats.
Apart from diploid species we subjected 10 high yielding varieties each from Triticum durum (AB) and Triticum aestivum (ABD) to SDS-PAGE to study the polymorphism of triticin subunits. A single band at the same level as that of the lowest band of Chinese spring was seen in case of all the 10 durum varieties. The banding pattern in all the ten-hexaploid wheats was similar to Chinese spring. Thus no polymorphism was seen among the 20 lines of cultivated wheat. Therefore we subjected only diploid Triticum and Aegilops species to PCR amplification, using C2/T13 primer to study, the polymorphism at the protein level, weather it could be due to differences at the genomic level, or some other post transcriptional modification.
The variations in the PCR amplification products of triticin gene are shown in Fig. 2. It showed that there was enough polymorphism at genomic level for different accession. The polymorphism was also found for different accessions of Ae. Squarosa, for which no polymorphism was found by SDS-PAGE of protein extracts, but no correlation, was found between amplification products of genomic DNA and triticin protein subunits.
 | |
| Fig. 1: | SDS-PAGE patterns of triticin subunits from seed extracts of different diploid Triticum and Aegilops species. CS-Chinese Spring |
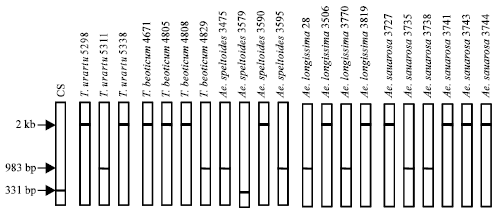 | |
| Fig. 2: | PCR amplification of triticin gene segment of different diploid Triticum and Aegilops species using C2/T13 primer |
The polymorphism observed at the DNA level may be due to the differences in the non-coding regions of the DNA such as introns, which are excised and not translated into proteins. Accessions of certain progenitors and related species like T. beoticum and Ae. longissima with high molecular weight triticin subunits, can be used for studying the sequence and increasing the triticin content through interspecific hybridization.
It has been reported that triticin gene on chromosome 1BS in Chinese spring and other T. aestivum and T. durum is not expressed (Singh et al., 1991). In this study the expression of triticin, in S-genome Ae. speltoides- the progenitor of B-genome and another S-genome species Ae. longissima, was reported. The accessions of S-genome species can be useful source for expression of B-genome or S-genome encoded triticin subunits, in hexaploid wheats for increasing their triticin/lysine content.
REFERENCES
- Dellaporta, S.L., J. Wood and J.B. Hicks, 1983. A plant DNA minipreparation: Version II. Plant Mol. Biol. Rep., 1: 19-21.
CrossRefDirect Link - Shailja, K., M. Rathore, N. Puri, D. Yadav and N.K. Singh, 2002. PCR amplification of hypervariable region of wheat triticin gene. J. Cereal Sci., 35: 129-134.
CrossRef