
Ishrak K. Khafagi
Suez Canal University, Egypt
Biotechnology
Year: 2007 | Volume: 6 | Issue: 2 | Page No.: 193-201







ABSTRACT
Seed germination, seedling growth and callus induction was compared between two varieties of Ricinus communis L. Callus cultures were initiated from roots, hypocotyls and cotyledonary leaves of the aseptically germinated seedlings on MS medium supplemented with 0.8% agar, 3% sucrose and various combinations of auxins and cytokinins. Seedling growth development and callus growth rate of large-seeded variety (var. major) was higher than that of small-seeded variety (var. minor). However, ricinine content was higher (6.59-7.97 μg g-1 FW) in callus cultures initiated from var. minor than that accumulated in var. major (2.44-4.35 μg g-1 FW). Broad spectrum antibacterial activity was recorded from ethanol and ether extracts of callus cultures of both varieties of R. communis L. against gram-positive and Gram-negative bacteria. Callus cultures subcultured for 3-6 subcultures diffused their antibacterial compounds into agar media during growth development. None of the callus extracts showed activity against Candida albicans. Bioautographic TLC analysis revealed that the antibacterial activity of var. major was due to quercetin and ricinine, while activity of var. minor was due to quercetin, kaempferol and ricinine. In vitro anthelmintic activity against Mesocestoides corti and Taenia crassiceps was recorded from callus cultures initiated from R. communis L. var. major only. Percent mortality of both organisms was high (93-95%) after incubation for 48 h in the presence of 20 μL root extracts of variety major.
PDF Abstract XML References
How to cite this article
Ishrak K. Khafagi, 2007. Variation of Callus Induction and Active Metabolite Accumulation in Callus Cultures of Two Varieties of Ricinus communis L. Biotechnology, 6: 193-201.
DOI: 10.3923/biotech.2007.193.201
URL: https://scialert.net/abstract/?doi=biotech.2007.193.201
DOI: 10.3923/biotech.2007.193.201
URL: https://scialert.net/abstract/?doi=biotech.2007.193.201
INTRODUCTION
Plant-derived antibiotics often originate from plant defense mechanisms. Secondary metabolites mediate defense mechanisms by providing chemical barriers mostly against microbial predators. This chemical warfare between plants and their pathogens consistently provides new natural product leads (Wedge and Camper, 2000). Opportunities of finding novel anti-infective drugs from plants may be increased by subjecting more plant extracts to broad array of bioassays and including extracts derived from plant cell, tissue and organ cultures too (Khafagi et al., 2003).
Production of secondary metabolites by plant tissue culture may allow the discovery of interesting compounds produced by natural plant populations in very small quantities to permit isolation and identification, or not produced at all in adult plants (Borris, 1996). Assessment of the antibacterial (Khafagi, 1998), antifungal (Salvador et al., 2003; Shariff et al., 2006), antiviral and cytotoxicity (Sökmen, 2001) produced by in vitro cultures in contrast with their corresponding adult plants were documented in the literature. However, literature concerning the biological activity of most medicinal plants lack information related to the variety used. Comparing metabolites accumulated in vitro in some plant varieties of the monotypic genus Ricinus L. may be valuable for future biotechnological application aiming at raising its in vitro secondary metabolite production.
Ricinus communis L. commonly known as castor bean belongs to family Euphorbiaceae. It is probably native to North East tropical Africa, but now widely cultivated throughout the tropics, subtropics and warm temperate regions (Meikle, 1985). In Egypt, it is mainly localized in the irrigated lands of the Nile valley and Delta with few populations in Egyptian deserts including Gabel Elba and Sinai (Shaheen, 2002). R. communis L. plants exhibit much variation regarding the size and ornamentation of the fruit and color patterning of the seeds, fruits, leaves and pollens. Worldwide, 20 varieties of R. communis L. have been recognized (Meikle, 1985). However, it is represented in Egypt by two varieties only, including the common large-seeded variety and another small-seeded variety. Based on the size of the seeds, R. communis L. has been classified into three varieties namely, major, intermedia and minor (Onwuliri and Anekwe, 2001).
Seeds of R. communis L. dating back to 4000 B.C. have been found in ancient Egyptian tombs (Hepper, 1990). Castor seeds contain a high percentage of oil that causes the purgative action, a toxic alkaloid, ricinine and a very toxic albumin called ricin (Dejey, 1975; Zohary, 1987). Castor oil is prescribed for infestation of intestinal worms. Infusion of the leaves was used as a remedy for rash, itch and eye inflammation. The decoction of roots is used for skin diseases, diarrhea and kidney and bladder troubles (Boulos, 1983). Ethanol-water extracts (1:1) of either roots or stems were reported to have in vitro antiamebic activity and cytotoxic effects against the 9KB carcinoma. In vitro antiviral activity and hypoglycemic activity were reported from leaf extracts (Ayensu, 1978).
Ethno-pharmacological reputation of R. communis L. is remarkable. It is valued for its oil, abandoned for its toxicity and treasured for its biological activity. Despeyroux et al. (2000) found that a large extent of ricin heterogeneity was originated for various R. communis varieties. Do different varieties of R. communis L. have similar characteristics?
The present study aims at comparing seedling growth and callus induction of R. communis L. var. major and var. minor, determining their ricinine content, investigating in vitro anthelmintic, antibacterial and anti-candidal activities in callus culture extracts of both varieties and testing the contribution of the alkaloid ricinine or the flavonols quercetin and kaempferol in the antibacterial action of callus culture extracts.
MATERIALS AND METHODS
Plant material: Seeds of Ricinus communis L. were collected from plants grown in Abo-Attwa, Ismailia, (var. major) and St. Catherine, South Sinai, Egypt (var. minor) in June 2003. Morphological characters of both varieties are listed in Table 1. A voucher specimen of each plant collection was deposited in the herbarium of the Botany Department, Faculty of Science, Suez Canal University, Ismailia, Egypt.
Seed germination: Seeds were surface sterilized by shaken for 20 min with a 50% (v/v) solution of a commercial bleach (20% sodium hypochlorite) followed by three washes in sterile distilled water. Ten replicates of 10 seeds each from two varieties of R. communis L. were placed on agar-solidified media or in jars lined with cotton and moisten with Hogland’s solution or distilled water. The seeds were germinated at 25°C in subdued light. The percentage of germination was determined after 5 days. Seedling growth was determined by measuring length (cm) of primary roots and hypocotyls during a period of 15 days.
| Table 1: | Comparison of morphological characters of two varieties of R. communis L. |
 | |
Callus induction and culture conditions: Fifteen day old aseptically germinated seedlings of both varieties were cut into root, hypocotyls and cotyledonary leaves and inoculated on MS medium (Murashige and Skoog, 1962) supplemented with 2 mg L-1 2,4-dichloropheoxyacetic acid (2,4-D) (MS1), 5 mg L-1 2,4-D (MS2), 2 mg L-1 naphtaleneacetic acid (NAA) + 0.2 mg L-1 kinetin (K) (MS3), 5 mg L-1 indole-3-acetic acid (IAA) + 0.1 mg L-1 K (MS4) and 1 mg L-1 IAA + 200 mg L-1 casein hydrolysate (MS5). Callus cultures appeared within 3-6 weeks after incubation at 25°C in the dark. The obtained calli were further subcultured on the same media every 4-6 weeks.
Media used in all experiments were solidified with 0.8% LAB M Plant Tissue Culture Agar MC29, Amersham and supplemented with 3% sucrose. Media were autoclaved for 15 min at 121°C. The pH of the media was adjusted to 5.8 before autoclaving. Growth regulators were co-autoclaved with other ingredients except IAA, which was sterilized with 0.22 μm sterile filter and added after autoclaving.
Ricinine determination: Ricinine was extracted from callus cultures initiated from two varieties of R. communis L. by homogenizing calli with 10% acetic acid in ethyl alcohol for 2 min. The homogenate was then filtered and the filtrate was concentrated in a rotary evaporator at 40°C until reduced volume. Alkaloids was precipitated with 25% ammonium and then centrifuged at 5000 g for 5 min. The precipitate containing ricinine was dissolved in 3 mL methanol, then diluted with phosphate buffer (pH = 5.0) and measured with a spectrofluorimetric technique modified from that developed by Sasse et al. (1980) for the determination of harmane alkaloids of Peganum harmala L. Ricinine was determined using an excitation wavelength 322 nm and an emission wavelength 410 nm. Ricinine content of crude extracts was calculated from a calibration curve prepared using a standard ricinine compound (Sigma Co.).
Phytochemical analyses: Phytochemical analyses were carried out as described by Trease and Evans (1987) and Evans (1996). Ethanol extracts of callus cultures initiated from R. communis L. var. major and var. minor were examined for the main biologically active secondary compounds including tannins, unsaturated sterols and terpenes, flavonoids, alkaloids, glycosides and saponins (Harborne, 1984).
Extracts preparations: Both callus tissues and stale agar initiated from the two varieties of R. communis were extracted with ethanol: diethyle ether (1: 1) for 5 min in a homogenizer. The proportion of callus/ solvent was 1:2 (w/v). The homogenate was left overnight at room temperature and then filtered. The filtrate was evaporated to dryness in a rotary evaporator at 40°C and the residue was dissolved in 2 mL ether and the residue left was dissolved again in 2 mL ethanol.
Test strains: Antimicrobial susceptibility tests were performed against Bacillus subtilis NRS-744, Staphylococcus aurea NRRL B-767, Klebsilla pneumonia NRRL B-3521, Escreichia coli NRRL B-3704, Proteus vulgaris NRRL B-123, Pseudomonas aeroginosa NRRL B-23 and Candida albicans NRRL Y-477. Microorganisms were obtained from United States Department of Agriculture, Northern Regional Research Laboratory (NRRL), Peoria, Illinois, USA.
Determination of antimicrobial activity: Sensitivity tests were performed by agar diffusion method (Ericsson and Sherris, 1971). An inoculum of each bacterial strain was suspended in 5 mL of Nutrient Broth and incubated overnight at 37°C for 18 h. Yeast cultures were suspended in 5 mL Sabouraud Dextrose Broth and incubated for 48-72 h at 30°C. The cultures were diluted with 1/10 with Broth before use.
Paper discs (6 mm) were impregnated with 20 μL of each extract and the solvent were evaporating at room temperature. After holding the plates at room temperature for 2 h to allow diffusion of extract into the agar, they were incubated for 24 and 48 h at 37°C for bacteria and yeast strains, respectively. The inhibition zone (mm) produced was measured after incubation.
Bioautgraphic TLC analysis: Ascending separation of callus and stale agar extracts prepared from two varieties of R. communis L. were carried out on pre-coated silica gel G 60 plates developed with chloroform: methanol (10: 4). The developed chromatograms were dried at room temperature to remove solvents. UV-active compounds were detected under ultraviolet light at 254 and 365 nm and marked on the TLC plate. A sample (5 mL) of bacterial suspension was dispersed evenly over the TLC plate using a roller covered with chromatography paper sheet. The TLC plates were incubated overnight at 30°C in a covered tray lined with wet chromatography paper (to keep humidity high and permit growth of the bacteria). The plates were then sprayed with 5 mL of an aqueous solution of p-iodonitrotetrazolium (5), then incubated again at 30°C for 4 h and examined for colorless inhibition zones against a violet background (Hamburger and Cordell, 1987). Standard ricinine, quercetin and kaempferol (Sigma, Co.) were subjected to the same separating conditions for identifying active metabolites.
In vitro anthelmintic bioassay: Fresh callus tissues initiated from roots, hypocotyls and cotyledonary leaves of two varieties of R. communis L. were extracted with ethanol for 5 min. in a homogenizer, then filtered. The filtrate was evaporated in a rotary evaporator at 40°C. The residue was dissolved in 2 mL sterile distilled water. In vitro anthelminthic activity of aqueous callus extracts was tested in pre-sterilized 24-well plates against sporocysts of the tap worms Mesocestoides corti and Taenia crassiceps. Liquid RPMI 1640 medium (Sigma, Co.) supplemented with L-glutamine and sodium bicarbonate was dispensed into the wells (1.5 mL/well). Each well was inoculated with 1 mL of the test organism (1 mL contains 15-18 sporocysts) and 20 μL crude callus extract. Distilled water was used as control. Viability of the sporocysts was observed under the light microscope after incubated at 25°C in the dark for 48 h period.
Statistical analysis: The final data were reported as the mean of 6 replicates for each treatment. Experiments were repeated three times and means and standard error of the means were calculated. Data of alkaloid determination were subjected to an analysis of variance (ANOVA) followed by HSD Tukey’s test (Zar, 1984, Lentner and Bishop, 1986) in order to test the significance (p = 0.05) between the two varieties.
RESULTS
Germination percentage, seedling growth, callus induction and ricinine formation in callus cultures were compared between two varieties of R. communis L. Accumulation of antimicrobial and anthelmintic metabolites in callus culture extracts initiated from roots, hypocotyls and cotyledonary leaves of both varieties on media supplemented with various combinations of plant growth regulators was tested.
Germination percentage and seedling growth: Prominent germination rates were produced for R. communis L. seeds germinated on cotton moisten with distilled water than those moisten with Hogland’s solution or seeds grown on agar-solidified media. Germination efficiency of the two varieties of R. communis L. was high (81.5-92%). Germination percentage of var. major was higher than var. minor. Seed germination of var. major started earlier than that of var. minor as seed coats of large seeds were cracked open two days in advance of small seeds (Fig. 1). Moreover, seedling growth of variety major produced longer seedlings than variety minor (Fig. 1).
Callus induction: Friable cream to yellow heterotrophic calli were produced from roots, hypocotyls and cotyledonary leaves of both varieties of R. communis L. Callus formation started within one week of inoculation on MS media supplemented with various combinations of auxins and cytokinins. Callus cultures subcultured for several passages retain their friability and healthy appearance after several subcultures on the same culture media. Generally, increase in biomass was higher in variety major than variety minor in all tested combinations of growth regulators (Fig. 2). Calli derived from both varieties and grown on media supplemented with 2,4-D (such as MS1 and MS2) grow faster than calli grow on media supplemented with auxin and cytokinin combinations (such as MS3 and MS4) (Fig. 2). However, 5 mg L-1 2,4-D was slightly inhibitory to callus growth of both varieties. Media supplemented with IAA and casein hydrolyzate was good for both varieties and certainly resulted in the best growth of var. minor.
Phytochemical composition: Callus cultures derived from hypocotyls of R. communis L. var. major and var. minor and grown on MS media with 2 mg mL-1 2,4-D were tested for the accumulation of main biologically active secondary metabolites. Ethanol extracts of calli derived from both varieties accumulated the same phytochemicals including alkaloids, flavonoids, anthocyanin, terpenes, tannins and glycosides (Table 2). Nevertheless, saponin was not present in callus culture extracts of both varieties (Table 2).
Ricinine content: Heterotrophic callus cultures of R. communis L. var. major and var. minor initiated from roots, hypocotyls and cotyledonary leaves accumulated the pyridine alkaloid ricinine in their tissues in the stationary growth phase after 28 days development in culture media. Generally, various auxin-cytokinin combinations sustain alkaloid formation (data not shown). Ricinine accumulation in calli grown on MS media supplemented with 2 mg mL-1 2,4-D was compared. It was higher (6.59-7.97 μg g-1 FW) in callus cultures derived from all explants of variety minor than that accumulated in calli derived from corresponding explants of variety major (2.44-4.35 μg g-1 FW) (Fig. 3).
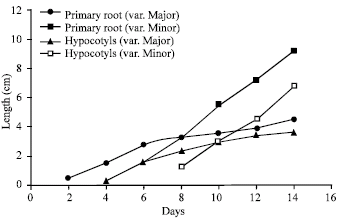 | |
| Fig. 1: | Comparison of seedling growth of Ricinus communis L. var. major and var. minor. Primary roots and hypocotyls (cm) of seedlings were measured for a period of 15 days. Six seeds were used for each determination. The experiment was repeated three times |
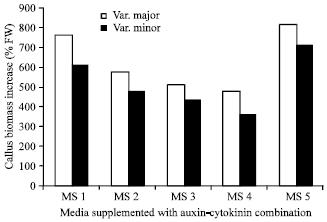 | |
| Fig. 2: | Effect of various combinations of auxins and cytokinins on callus biomass increase (% FW) of two varieties of Ricinus communis L. Fresh weight of calli derived from hypocotyls of both varieties were compared using Murashig-Skoog medium supplemented with 2 mg L-1 2,4-D (MS1), 5 mg L-1 2,4-D (MS2), 2 mg L-1 NAA + 0.2 mg L-1 K (MS3), 5 mg L-1 IAA + 0.1 K (MS4) or 1 mg L-1 IAA + 200 mg L-1 casein hydrolyzate (MS5) |
Significantly higher accumulation level of ricinine at p = 0.05 was confirmed for R. communis L. calli derived from var. minor over that produced in calli derived from var. minor (Fig. 3).
In vitro accumulation of antimicrobials: Results presenting the antimicrobial potential of ethanol and ether extracts of callus cultures derived from hypocotyls of two varieties of R. communis L. grown on MS medium supplemented with 2 mg L-1 2,4-D are illustrated in Fig. 4.
| Table 2: | Phytochemical composition of callus cultures initiated from the hypocotyls of two varieties of Ricinus communis L. |
 | |
| + = Positive, - = Negative | |
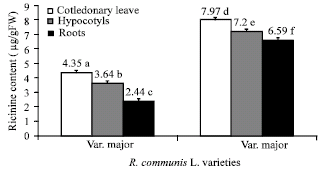 | |
| Fig. 3: | Ricinine content (μg g-1 FW) of callus cultures derived from roots, hypocotyls and cotyledonary leaves of two varieties of Ricinus communis L. Callus cultures were grown on media supplemented with 2 mg mL-1 2,4-dichlorophenoxyacetic acid. Mean values followed by different letters are significantly different at p≤0.05 |
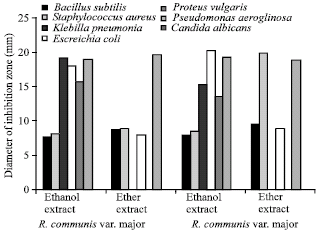 | |
| Fig. 4: | Antimicrobial activity of ether and ethanol extracts of callus cultures initiated from hypocotyls of two varieties of Ricinus communis L. on media supplemented with 2 mg L-1 2,4-D. Activity was recorded against seven test strains including two Gram-positive bacteria, four Gram-negative bacteria and a yeast. Antimicrobial activity is expressed as diameter of inhibition zones (mm) |
Callus culture extracts of both varieties exhibited broad spectrum antibacterial activity against most test strains. Active compounds, against Gram-positive as well as Gram-negative bacteria mutually exist in ethanol and ether extracts of callus cultures (Fig. 4). None of the callus extracts showed activity against Candida albicans. Various plant growth regulators added to growth media supported the accumulation of antimicrobials in callus tissues of both varieties. Callus derived from roots and cotyledonary leaves of both varieties contained the same spectrum of antibacterial activity as shown from hypocotyls activity (Data not shown).
Subsequent subcultures of callus tissues had a tendency to diffuse their active metabolites to agar during their in vitro growth course (Fig. 5). Callus cultures subcultured for 3-6 subcultures diffused their antibacterial compounds into agar media. Ether extracts of stale agar of both varieties were active against all test bacterial strains with higher activity from variety minor (Fig. 5). On the other hand, both ethanol and ether extracts of callus tissues lack antibacterial activity. None of the agar cultures extracts showed activity against Candida albicans (Fig. 5).
Bioautographic TLC analysis revealed that the antibacterial activity of R. communis L. variety major was due to the presence of quercetin and ricinine compounds, while activity of variety minor due to quercetin, kaempferol and ricinine (Table 3). Inhibition zone around ricinine spot was the largest followed by that of quercetin. Spots corresponding to ricinine, quercetin and kaempferol were eluted from TLC chromatogram and tested in combinations. It was found that quercetin and kaempferol have additive antibacterial effect. On the other hand, ricinine with either quercetin or kaempferol produced synergetic effect against all test bacterial strains.
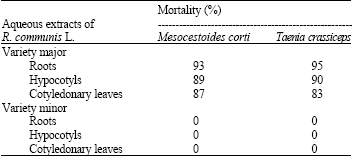
In vitro anthelmintic action: In vitro anthelmintic activity was recorded from callus cultures derived from roots, hypocotyls and cotyledonary leaves of aseptically-germinated seedlings of R. communis L. var. major against Mesocestoides corti and Taenia crassiceps. Percent mortality of both organisms was high (93-95%) after incubation for 48 h in the presence of 20 μL root extracts of variety major (Table 4). Aqueous extracts of calli derived from cotyledonary leaves of variety major produced less anthelmintic action (83-87%) compared with extracts of calli derived hypocotyls (89-90%). R. communis L. var. minor did not show any in vitro anthelmintic activity against either test organisms (Table 4).
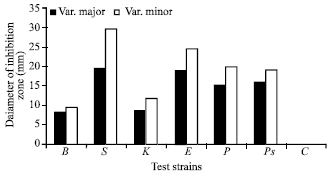 | |
| Fig. 5: | Antimicrobial activity of ether extracts of diffused active compounds from stale agar of the third subculture of callus initiated from hypocotyls of two varieties of Ricinus communis L. Antimicrobial activity was determined as diameter of inhibition zone (mm) against Bacillus subtilis NRS-744 (B), Staphylococcus aureus NRRL B-767 (S), Klebsilla pneumonia NRRL B-3521 (K), Escreichia coli NRRL B-3704 (E), Proteus vulgaris NRRL B-123 (P), Pseudomonas aeroginosa NRRL B-23 (Ps) and Candida albicans NRRL Y-477 (C) |
| Table 3: | Presence (+) or absence (-) of active metabolites in ether extracts of callus cultures initiated from hypocotyls of Ricinus communis L. var. major and var. minor as determined by bioautographic TLC analysis against Staphyllococcus aureus |
 | |
| The TLC chromatogram was developed in chloroform: methanol (10:4). The flavonols quercetin and kaempferol were exposed to ammonia fumes and detected by yellow fluorescence under UV light. The alkaloid ricinine was detected by Dragendorf’s reagent and fluorescence under UV light. +++ = Highly active | |
| Table 4: | In vitro antihelmenthic activity of two varieties of Ricinus communis L. |
 | |
DISCUSSION
Plant tissue culture approach has offered potent techniques, which may significantly contribute to the scheme of increasing levels of medicinally useful secondary metabolites (Ferreira and Duke, 1997; Duke et al., 1999; Khafagi et al., 2003). The present study compared growth characteristics of seedlings and callus cultures derived from seedling explants of two varieties of Ricinus communis L. grown on defined media supplemented with some auxin and cytokinin combinations. In vitro accumulation of secondary metabolites in callus tissues of both varieties was compared as well.
Results obtained in the present study demonstrated that R. communis L. large-seeded variety major had higher seedling and callus growth rate compared with the small-seeded variety minor. However, ricinine accumulation was significantly higher in callus cultures derived from roots, hypocotyls and cotyledonary leaves of R. communis L. variety minor than that accumulated in callus tissues derived from corresponding explants of variety major. It was commonly established that slow growing callus cultures tend to accumulate higher levels of secondary metabolites compared with fast growing callus cultures (Ferreira and Duke, 1997).
Application of biotechnological systems to produce some compounds produced from such medicinally important R. communis L. plant should be more rational after thoroughly exploring various plant varieties for in vitro growth and major metabolite production. Despeyroux et al. (2000) highlights the existence of ricin heterogeneity that is originated for various R. communis varieties, which may necessitate the importance of exploring various R. communis L. varieties for active metabolites. This is simply to relate unusual behavior to the particular variety used instead of extend that to include all R. communis varieties.
Major phytochemicals such as tannins, alkaloids, flavonoids and terpenoids that are known to be present in ethanol extracts of adult R. communis L. plant (Kang et al., 1985) was found in ethanol extracts of callus cultures initiated from seedling explants of both varieties. However, differences in the amount and/or spectrum of alkaloids and flavonoids were noticed for the two studied varieties. Significantly higher levels of ricinine (6.59-7.97 μg g-1 FW) was accumulated in callus cultures initiated from various explants of variety minor than that accumulated in callus of variety major (2.44-4.35 μg g-1 FW). On the other hand, the additive antibacterial effect of quercetin and kaempferol as well as the synergetic action of ricinine with either quercetin or kaempferol against test microorganisms in the susceptibility test, may demonstrate the higher antibacterial activity of extracts of variety minor over that of variety major that lack the flavonol kaempferol and consequently lack its direct and interactive antimicrobial effect, which may have implication for the activity of callus of various varieties. Comparable results were reported by Adbel-Wahab (2001), who found variations in the electrophoretic protein patterns in roots and leaves of R. communis L. populations of deserts and riverine, which are also corresponding to large-seeded and small-seeded varieties. Such varieties were represented by the appearance of some newly synthesized polypeptides and disappearance of some others. Also, differences in the accumulation of potassium, calcium, magnesium, soluble carbohydrates, soluble protein, praline and amino acids content of the two populations was reported (Adbel-Wahab, 2001). Additionally, Shaheen (2002) found clear morphological differentiation within R. communis L. South-Eastern desert populations and those of riverine, Deltatic and Sinai populations, which may lead to assurance of having two varieties in Egypt. He suggests that morphological data should be tested against chemical data.
Diffusing out active metabolites produced by callus cultures of R. communis L. was an observation confirmed from some other comparable research studies. It was noticed that ribosome-inactivating proteins and agglutinins synthesized from callus and cell suspension cultures initiated from seedling explants of R. communis L. were secreted into the medium (D’Silva et al., 1993). Also, cell suspension cultures of R. communis L. diffused out kaurene synthetase inhibitor intercellularly and to the growth media (Gafni and Shechter, 1981). Although, it is usually advantage to prove accumulation of secondary metabolites in vitro cultures, it is particularly favorable if cultured cells diffuse out their toxic metabolites to the surrounding media, rendering cultured cells or tissues without toxicity. Furthermore, the observation that none of in vitro culture extracts showed activity against Candida albicans, though; their adult and seedling extracts have moderate anti-candidal activity was also noticed for ten plant tissue culture extracts (Khafagi, 1998). Activity of callus extracts against prokaryotic cells and not eukaryotic one may reveal the lack of toxicity against eukaryotes, which may help developing useful antibacterial drugs for animals and human, since R. communis L. adult plant is one of the highly toxic plants that cause both DNA damage and chromosomal aberrations (Fennell et al., 2004).
The present study reveals the in vitro production of secondary metabolites from callus cultures of the medicinally important plant R. communis L., which has broad medicinal uses that are documented in the ethnobotanical literature (Ayensu, 1978; Boulos, 1983; Rizk, 1986). Recent ethnopharmacological research studies had regularly confirmed most of the therapeutic values reputed for such well-known medicinal plant. For instance, methanol extracts of R. communis L. root were reported to possess significant anti-inflammatory activity in acute and chronic inflammatory models in rats (Ilavarasan et al., 2006), stimulant effects of ricinine on the central nervous system of mice, which cause memory-improving effect and seizure-eliciting properties was reported and may post ricinine for the treatment of human amnesias (Ferraz et al., 1999), relation of the insecticidal properties of R. communis foliage extracts to the flavoniods quercetin and kaempferol (Upasani et al., 2003). Fennell et al. (2004) confirmed the anthelmintic activity of R. communis L. extracts. Immanuel et al. (2004) reported a potent antimicrobial action of n-butanol extract of R. communis L. against shrimp pathogen Vibrio parahaemolyticus. Sitton and West (1975) descried the production of the anti-fungal diterpene casbene in cell-free extracts of R. communis L. seedlings. Antifungal activity of fatty acids against the symbiotic fungus Leucoagaricus gongylophorus and insecticidal activity of ricinine against the leaf-cutting ant Atta sexdens rubropilosa were established (Bigi et al., 2004). Also, antifertility effects of 50% ethanol extract was revealed, which produced alteration in the motility, mode of movement and morphology of the sperms of male rats (Sandhyakumary et al., 2003).
Bedouins usually utilize castor oil for the infestation of intestinal worms (Zohary, 1987; Rizk, 1986). However, in vitro anthelmintic activity with high mortality 92-95% against M. corti and T. crassiceps was reported from the large-seeded variety major only. This result may raise the question about the variety used in folk medicine of Sinai Bedouins. Fennell et al. (2004) reported that R. communis L. possess antihelmintic activity, without mentioning the variety used for the bioassay.
In conclusion, apparent variation in seedling growth and callus induction rates between Ricinus communis L. var. major and var. minor was confirmed. Earlier and higher seedling growth of variety major accompanied by higher increase in callus biomass compared by slow growth produced from either seedlings or callus cultures of variety minor demonstrated such variation. This variation in growth was accompanied with opposite variation in secondary metabolite accumulation. Ricinine content of callus cultures of variety minor was higher than that of variety major. Also, quercetin and kaempferol was accumulated in callus of variety minor, while callus of variety major accumulated quercetin only. Confirmation of the presence of in vitro anthelmintic activity in callus extracts of R. communis L. variety major may verify its reputation in folk medicine. However, lack of in vitro anthelmintic activity from variety minor signifies the variation between the two varieties.
REFERENCES
- Bigi, M.F., V.L. Torkomian, S.T. Groote, M.J. Hebling and O.C. Bueno et al., 2004. Activity of Ricinus communis (Euphorbiaceae) and ricinine against the leaf-cutting ant Atta sexdens rubropilosa (Hymenoptera: Formicidae) and the symbiotic fungus Leucoagaricus gongylophorus. Pest Manage. Sci., 60: 933-938.
Direct Link - Borris, R.P., 1996. Natural products research: Perspectives from a major pharmaceutical company. J. Ethnopharmacol., 51: 29-38.
CrossRefDirect Link - Despeyroux, D., N. Walker, M. Pearce, M. Fisher, M. McDonnell, S.C. Bailey, G.D. Griffiths and P. Watts, 2000. Characterization of ricin heterogeneity by electrospray mass spectrometry, capillary electrophoresis and resonant mirror. Anal. Biochem., 279: 23-36.
Direct Link - Podder, K.S., I. D'Silva and C.S. Vaidyanathan, 1993. Ribosome-inactivating proteins and agglutinins from callus and suspension cultures of Ricinus communis L. and Abrus precatorius L. Plant Sci., 94: 161-172.
Direct Link - Duke, S.O., A.M. Rimando, M.V. Duke, R.N. Paul, J.F.S. Ferreira and R.J. Smeda, 1999. Sequestration of Phytotoxins by Plants: Implications for Biosynthetic Production. In: Biologically Active Natural Products: Agrochemicals, Cutler, H.G. and S.J. Cutler (Eds.). CRC Press, Boca Raton, Florida, USA., pp: 127-136.
- Fennell, C.W., K.L. Lindsey, L.J. McGaw, S.G. Sparg and G.I. Stafford et al., 2004. Assessing African medicinal plants for efficacy and safety: Pharmacological screening and toxicology. J. Ethnopharmacol., 94: 205-217.
CrossRefPubMedDirect Link - Ferraz, A.C., M.E. Angelucci, M.L. Costa, I.R. Batista, B.H. De-Oliveira and C. Da-Cunha, 1999. Pharmacological evaluation of ricinine, a central nervous system stimulant isolated from Ricinus communis. Pharmacol. Biochem. Behavior., 63: 367-375.
Direct Link - Ferreira, J.F.S. and S.O. Duke, 1997. Approaches for maximizing biosynthesis of medicinal plant secondary metabolites. AgBiotech News Inform., 9: 309-315.
Direct Link - Hamburger, M.O. and G.A. Cordell, 1987. A direct bioautographic TLC assay for compounds possessing antibacterial activity. J. Nat. Prod., 50: 19-22.
CrossRef - Ilavarasan, R., M. Mallika and S. Venkataraman, 2006. Anti-inflammatory and free radical scavenging activity of Ricinus communis root extract. J. Ethnopharmacol., 103: 478-480.
CrossRefPubMedDirect Link - Immanuel, G., V.C. Vincybai, V. Sivaram, A. Palavesam and M.P. Marian, 2004. Effect of butanolic extracts from terrestrial herbs and seaweeds on the survival, growth and pathogen (Vibrio parahaemolyticus) load on shrimp Penaeus indicus juveniles. Aquaculture, 236: 53-65.
CrossRefDirect Link - Kang, S.S., A. Cordell, D.D. Soejarto and H.H.S. Fong, 1985. Alkaloids and flavonoids from Ricinus communis. J. Nat. Prod., 48: 155-156.
CrossRefDirect Link - Khafagi, I.K., A. Dewedar and M. Amein, 2003. Opportunities of finding novel anti-infective agents from plant cell cultures. Curr. Med. Chem. Anti-infective Agents, 2: 191-211.
Direct Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Onwuliri, V.A. and G.E. Anekwe, 2001. Amino acids and other biochemical components of Ricinus communis (Variety Minor), an anti-conceptive seed. Pak. J. Biol. Sci., 4: 866-868.
CrossRefDirect Link - Salvador, M.J., P.S. Pereira, S.C. Fran�a, R.C. Candido, I.Y. Ito and D.A. Dias, 2003. Comparative study of antibacterial and antifungal activity of callus culture and adult plants extracts from Alternanthera maritima (Amaranthaceae). Brazilian J. Microbiol., 34: 131-136.
Direct Link - Sandhyakumary, K., R.G. Bobby and M. Indira, 2003. Antifertility effects of Ricinus communis (Linn) on rats. Phytother. Res., 17: 508-511.
CrossRefPubMedDirect Link - Shaheen, A.M., 2002. Morphological variation within Ricinus communis L. in Egypt: Fruit, leaf, seed and pollen. Pak. J. Biol. Sci., 5: 1202-1206.
CrossRefDirect Link - Shariff, N., M.S. Sudarshana, S. Umesha and P. Hariprasad, 2006. Antimicrobial activity of Rauvolfia tetraphylla and Physalis minima leaf and callus extracts. Afr. J. Biotechnol., 5: 946-950.
Direct Link - Sokmen, A., 2001. Antiviral and cytotoxic activities of extracts from the cell cultures and respective parts of some Turkish medicinal plants. Turk. J. Biol., 25: 343-350.
Direct Link - Upasani, S.M., H.M. Kotkar, P.S. Mendki and V.L. Maheshwari, 2003. Partial characterization and insecticidal properties of Ricinus communis L. foliage flavonoids. Pest Manage. Sci., 59: 1349-1354.
CrossRefDirect Link - Zar, J.H., 1984. Biostatistical Analysis. 2nd Edn., Prentice-Hall Inc., Englewood Cliffs, New Jersey, USA, Pages: 718.
Direct Link
A.vijender reddy Reply
sir i have read ur article i have very much intrested in ur article after reading ur i have found it also used for antihepatitis socan i use ur article for the research artice as a review
waiting for ur reply