
G.H. Shahidi Bonjar
College of Agriculture Sciences, Bahonar University of Kerman, Iran
Sonia Aghighi
College of Agriculture Sciences, Bahonar University of Kerman, Iran
Biotechnology
Year: 2005 | Volume: 4 | Issue: 2 | Page No.: 108-113







ABSTRACT
Among soil-borne fungi, cosmopolitan phytopathogen, Verticillium dahliae Klebahn is responsible for high yield losses in many plant species. Except for solarization in mediterranean countries and except in the few cases where disease-resistant cultivars are available, control of Verticillium wilt in commercial crops has been highly dependent on the application of preplant soil fumigants. Research to develop alternative control measurements should focus on biological approaches aimed at shifting the composition of soil microbial communities to suppress Verticillium. The merits of role of actinomycetes in biological control of soil-borne fungal-pathogens are known, however actinomycetes microflora of the Iranian soils has not been very well explored in searching for biofungicide agents. At the present research, in vitro studies of some biological effects of two Iranian strains of actinomycetes, Streptomyces plicatus strain 101 and Frankia sp. strain 103, are presented. Both strains revealed enzymatic activity and inhibited production of microsclerotia in V. dahliae. Treating the crude extract with chloroform, denaturized enzymatic activity of both strains. Thermal inactivation point of active phases of S. plicatus was 70 and 90°C and in Frankia sp. was determined as 60°C. Antifungal active phases of S. plicatus tolerate wide range of pH (5-13) but in Frankia sp. active phase tolerates pH 7-9. These two strains may be useful candidates for involving in integrated control programs of Verticillium vascular wilting.
PDF Abstract XML References
How to cite this article
G.H. Shahidi Bonjar and Sonia Aghighi, 2005. Chitinolytic and Microsclerostatic Activity of Iranian Strains of Streptomyces
plicatus and Frankia sp. on Olive Isolate of Verticillium
dahliae. Biotechnology, 4: 108-113.
DOI: 10.3923/biotech.2005.108.113
URL: https://scialert.net/abstract/?doi=biotech.2005.108.113
DOI: 10.3923/biotech.2005.108.113
URL: https://scialert.net/abstract/?doi=biotech.2005.108.113
INTRODUCTION
The genus Verticillium Nees (1817) represents one of the world's major pathogens, affecting crop plants mostly in the cool and warm temperate regions, but has also been reported from subtropical and tropical areas. V. dahliae is capable of surviving in field soil in several different ways. Various types of mycelia, clusters of hyaline cells and melanized microsclerotia have all been observed to germinate on assay plates used to determine the concentration of V. dahliae in naturally infested soils. These various forms of the ftmgus may be fonnd in soil essentially free from host tissue or embedded in small-to-large fragments of tissue[1]. Wide host range, long persistence in soil, propagation on rhizosphere of non- host plants and lack of effective control measurements highlights the need for research in evaluation of new methods for control of this pathoged [3]. In future, biological control of soil-borne frmgal or bacterial pathogens will be of increasing importance for a more sustainable agriculture. Furthermore, fnngicides (biocides) such as methyl bromides will be phased out and thus potential alternatives are needed to control the soil-borne pathogen such as V. dahliae. This has prompted the search for reliable antagonists which show a high degree of competitiveness and are active in the rhizospheres of different crops and in different soil types[4-5]. Some workers have reported biological control of Verticillium by Streptomyces spp[6]. Chi and Hanson[7] reported in vitro antiftmgal activity, ftmgistatic and frmgicidal effects of S. rimosus against V. dahliae and V. albo-atrum. Ezrukh[8] showed metabolites of actinomycetes from a cotton rhizosphere to interact with V. dahliae. Today, a commercial biocontrol agent Rhizovit® basis on spores of Streptomyces rimosus HR071 (DSM 12424) is formulated in Germany. It is a phytoprotectant biofnngicide and can be used to control several seed-borne and soil-borne ftmgal pathogens such as Verticillium, Pythium and Rhizoctonia both in vitro and in vivd [9]. With extended environmental diversity, however, the actinomycetes microflora of the Iranian soils has not been very well explored with the goal of exploring new means of biocontrols. With the merits of their role in biological control of soil-borne ftmgal-pathogens, at the present research, in vitro studies of some biological effects of two Iranian strains of actinomycetes such as enzymatic activity, effect on melanin biosynthesis and microsclerotia formation on are presented.
MATERIALS AND METHODS
Preparation of V. dahliae isolate: A registered phytopathogenic olive isolate of V. dahliae was used. It was obtained from Prof Banihashemi, Mycology Laboratory of the Department of Plant Pathology, College of Agriculture, Shiraz University, Shiraz, Iran. The fnngus was isolated from diseased olive trees with vascular wilting in Gorgan Province, Iran.
Culture media: Casein glycerol (or starch) agar (CGA) was used for supporting actinomycetes strains which composed of glycerollO g, casein 0.3 g, KNO3, 2 g, NaCl, 2 g, K2HPO4, 2 g, MgS04,.7H2O 0.05 g, CaCO3, 0.02 g, FeS04.7H2O 0.01 g and agar 18 g in 1 L of distilled H2O (pH 7. 2)[10]. In submerged cultures, agar and glycerol was excluded and soluble starch was used (CG medium). Actinomycetes colonies were selected and transferred to CGA slants for fwiher studies. All V. dohlioe isolates were maintained on potato dextrose agar (FDA) (Difco).
Submerged culture of active strains and preparation of crude extract: The two active strains were grown in submerged cultures of CG medium on rotary shakers rmder 130 rpm at 30°C. To monitor the activity, aseptically small aliquots of culture media were taken every 24 h for 20 days and the activity was evaluated by well diffusion-method[10,11]. When the activity reached maximum, the cultures were hanrested; spores and mycelia were excluded by filtration through two layers of cheese cloth. The clarified sap was then dried to dark crude rmder reduced air at 30°C and kept refrigerated before use.
Classification of actinomycete: Actinomycete colonies were characterized morphologically and physiologically following the direction mentioned in the methods manual of international cooperative project for description and deposition of cultures of Streptomyces (ISP)l 12 Identification procedures were done by pH of the reaction mixtures was varied using the buffers Saadmm et al[14-15] Department of Biological Sciences, described by Covington and Davisod[13,14] The pH stability University of Science and Technology, Irbid, Jordan[15]
In vitro antifungal bioassays Agar disk-method: The two active Iranian actinomycete isolates reported previously[16.18l were smeared on CGA medium as a single streak and after incubation at 28°C for 4-6 days, from well-grown streaks 6 mm agar disks of actinomycete colony masses were prepared by using sterile cork borers. Disks were then aseptically transferred to FDA plates having fresh lawn cultures of V. dohlioe isolates. Controls included using plain disks from CGA medium. Plates were incubated at 26°C for 4-6 days and frmgicidal property indicative of chitinolytic activity[19-20] and/or microsclerostatic activity were evaluated by measuring the diameter of inhibition (mm).
Well diffusion method: For evaluation of antifnngal activity of aqueous samples, by use of sterile cork borer, wells (6x4 mm, 2 em apart) were pnnctured in fresh lawn cultures or at 30 mm distance from plugs of V. dahliae isolates. Respective concentrations in dimethyl sulfoxide: methanol (1/1: v /v) solvent (DM solvent) were then administered to fullness in each well. Plates were incubated at 26°C for 4-6 days for lawn cultures and 14 days for dual culture disk-plugs. Plates were incubated at 26°C for 4-6 days and fnngicidal property indicative of chitinolytic activityP 9· 21l and/or microsclerostatic activity were evaluated by measuring the diameter of inhibition zones (rnrn). Each experiment was repeated three times and the mean of inhibitory zones recorded. Controls included use of blank wells and use of DM solvent without test componnds.
Effect of heat and chloroform on bioactivity: To monitor the effect of temperature on bioactivity, small aliquots (10 mg ml-1) of soluble crude were exposed to each of 30, 40, 50, 60, 70, 80 and 90°C for 10 min and cooled on ice afterwards[22] Bioactivity of treated samples was evaluated using well diffusion method. Control included incubation of an rmtreated sample at 26°C. For determination of sensitivity to chloroform, small aliquots of aqueous solutions from 7 day old submerged cultures were vigorously mixed with equal volumes of chloroform for 10 min, centrifuged at 4000 rpm for 10 min, phases were then separated and dried at 40°C rmder reduced air and placed in a desiccator overnight. All samples were tested by well diffusion method as described earlier.
Effect of pH on antifungal bioactivity: Effect of pH on activity and stability of activity was measured at different pH values by the general standard assay methods. The pH of the reaction mixtures was varied using the buffers described by Covington and Davisod[23]. The pH stability of the active crude sap was evaluated by incubating it for 0.5 hat various pH values at 30°C and evaluating them by agar well diffusion method.
RESULTS AND DISCUSSION
Cultures of active strains: Pure cultures of Streptomyces plicatus and Frankia sp. in slant CGA medium are indicated in Fig. 1. Streak cultures are indicated in Fig. 2. In situ morphology of aerial mycelia in slide cultures of S. plicatus nnder light microscope is indicated in Fig. 3.
| Table I: | Bioactivity of Streptomyces and Frankia sp. on Verticillium dah/iae determined by agar disk method in mm Actinomycetes |
| CZ: Clear zone of complete inhibition of fungal growth adjacent to tested agar disks of actinomycetes; MsiZ: Microsclerostatic or microsclerotial inhibition zone, representing inhibitory e±Iect on microsclerotia formation and development; TI: Total inhibition zone of fungal growth | |
| Table 2: | Identification criteria of Frankia sp. based on morphological and physiological characterizations |
 | |
| W: White; 1: Positive; 0: Negative;-: No utilization;+: Utilization. Low sporulation on Oatmeal agar medium, diameter of the hyphae is considerably larger than of Streptomyces, the myceliwn (sporophore) ends to large club shaped sporangium | |
| Table 3: | Effect of chloroform on bioactivity of 7-days old submerged cultures and crude extracts of Streptomyces plicatus and Frankia sp. indicated by diameter of inhibition zones in mm |
| A: Control, 7-days old aqueous submerged cultures; B: chlorofonn phase and aqueous phase© after treated with chloroform; D: Dried crude prepared from 7-days old aqueous submerged cultures. CZ: Clear zone of complete fungal growth inhibition; MsS: Microsclerostatic zone or microsclerotial inhibition zone | |
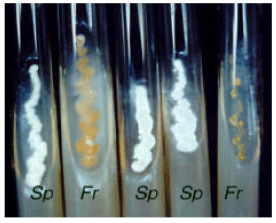 | |
| Fig. 1: | Pure cultures of Streptomyces plicatus (Sp) and Frankia sp. (Fr) in slant CGA medium |
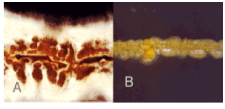 | |
| Fig. 2: | Streak cultures of Streptomyces plicatus (A) and Frankia sp. (B) on CGA medium |
 | |
| Fig. 3: | In situ morphology of aerial mycelia of Streptomyces plicatus in slide cultures under light microscope (100 X) |
 | |
| Fig. 4: | Clockwise from top, well method bioassay results of Streptomyces plicatus, Frankia sp. against Verticillium dahliae and blank well (control) |
 | |
| Fig. 5: | Shake flasks of submerged cultures of Streptomyces plicatus (A) and Frankia sp. (B) in CG liquid culture media 7 days after inoculation |
Bioassays: Well method bioassay results of S. plicatus and Frankia sp. are indicated in Fig. 4 and Table I.
Submerged cultures and crude extract: Shake flasks containing submerged cultures of S. plicatus and Frankia sp. are indicated in Fig. 5.
 | |
| Fig. 6: | Colonies of streptomyces plicatus from 7 days old aqueous submerged cultures of shake flasks under binocular microscope (40 X) |
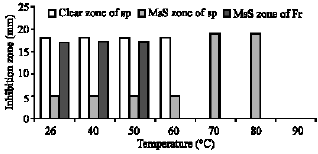 | |
| Fig. 7: | Temperature effect on antifungal bioactivity of crude extract of ::lreptomyces plicatus and Frankia sp. Sp.: streptomyces plicatus, Fr: Frankia sp., MsS: Microsclerostatic zone or microsclerotial inhibition zone |
Colonies from submerged culture under binocular microscope are indicated in Fig. 6. The resulted crude extract composed of dark brown residue which was divided to small portions and preserved under refrigeration conditions at 4°C before use.
Classification of actinomycete: streptomyces plicatus strain 101 was identified and reported as a new record from Iran previously[16] Table 2 shows the identification criteria of Frankia sp. based on morphological and physiological characterization.
Effect of heat and chlorofonn on activity:Temperature effect on antifungal activity of S. plicatus and Frankia sp. are indicated in Fig. 7. Effect of chloroform on bioactivity of both strains is indicated in Table 3.
Effect of pH on activity: Effect of pH on antifungal bioactivity of 7 days old aqueous submerged cultures of streptomyces plicatus and Frankia sp. is indicated in Fig. 8.
Since there is high variation in the soil ecology of Iran, the population diversity of the actinomycetes in Iranian soils has to be further explored for new active isolates. Present findings represent the presence of potential antifungal metabolite(s) in both isolates against olive isolate of V. dahliae. ::lreptomyces plicatus prohibits mycelial growth and formation of melanin in the pathogen. It can be postulated that lack of melanin
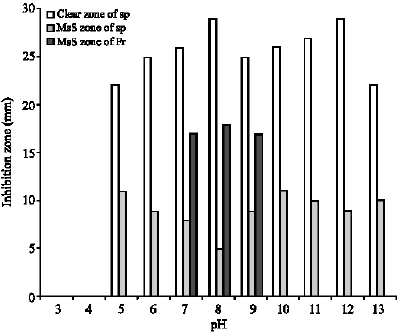 | |
| Fig. 8: | Effect of pH on antifungal bioactivity of 7 days old aqueous submerged cultures of streptomyces plicatus and Frankia sp. Sp.: streptomyces plicatus, Fr: Frankia sp., MsS: Microsclerostatic zone or microsclerotial inhibition zone |
renders the pathogen more susceptible to antagonists and promotes the conditions needed for its biological control in the rhizosphere. To enhance the tolerance and resistance of plants to fnngal attack and lower or eliminate the use of envirornnentally nnsafe ftmgicides, the genes for antiftmgal metabolites can be engineered into plants. It can be forecasted that "oncoming future" does not tolerate further pollution resulting from vast use of chemical pesticides. Hence, environmentally safe and non chemical-measures have to be developed to control plant maladies. In ideal biological control measures, proper micro organisms are those having well adaptation in soil and rhizosphere by exerting effective antagonistic activity against soil pathogens persistently. In this regard soil-driven actinomycetes do not have adverse effect or alter the biological balance of soils as chemical measures do. They should receive higher attention in research for biological controls worldwide. Conclusively, it may be assumed that the antifnngal-metabolite gene from Streptomyces plicatus may be a useful candidate for genetic engineering of olive plants for increased tolerance against V. dahliae. Findings of this and similar researches may contribute for evolvement of resistant transgenic-plants with recombinant DNA having antiftmgal genes cloned from biologically active actinomycetes.
ACKNOWLEDGMENTS
Thanks to Head of Research Affairs Office of Bahonar University of Kerman for financial support of the project and helpful information and kind gift of Verticillium dahliae isolate by Prof. Banihashemi, Dept. of Plant Protection, College of Agriculture, Shiraz, Iran. We also would like to thank helpful works of Dr. Saadonn and his coworkers for identification of actinomycetes. This research is dedicated to Mr. A. Afzalipour, the fonnder of Bahonar University in Kerman.
REFERENCES
- Smalla, K., G. Wieland, A. Buchner, A. Zock, J. Parzy, S. Kaiser, N. Roskot, H. Heuer and G. Berg, 2001. Bulk and rhizosphere soil bacterial communities studied by denaturing gradient gel electrophoresis: Plant-dependent enrichment and seasonal shifts revealed. Applied Environ. Microbiol., 67: 4742-4751.
- Shirling, E.B. and D. Gottlieb, 1966. Methods for characterization of Streptomyces species. Int. J. Syst. Evol. Microbiol., 16: 313-340.
CrossRefDirect Link - Saadoun, I., F. Al-Momani, H. Malkawi and M.J. Mohammad, 1999. Isolation, identification and analysis of antibacterial activity of soil streptomycetes isolates from North Jordan. Microbios, 100: 41-46.
Direct Link - Shahidi-Bonjar, G.H., M.H. Fooladi, M.J. Mahdavi and A. Shahghasi, 2004. Broadspectrim, a novel antibacterial from Streptomyces sp. Biotechnology, 3: 126-130.
CrossRefDirect Link - Aghighi, S., G.H.S. Bonjar and I. Saadoun, 2004. First report of antifungal properties of a new strain of Streptomyces plicatus (Strain 101) against four Iranian phytopathogenic isolates of Verticillium dahliae, a new horizon in biocontrol agents. Biotechnology, 3: 90-97.
CrossRefDirect Link - Aghighi, S., G.H.S. Bonjar, R. Rawashdeh, S. Batayneh and I. Saadoun, 2004. First report of antifungal spectra of activity of Iranian actinomycetes strains against Alternaria solani, Alternaria alternate, Fusarium solani, Phytophthora megasperma, Verticillium dahliae and Saccharomyces cerevisiae. Asian J. Plant Sci., 3: 463-471.
CrossRefDirect Link - Kong, L.D., D.D. Tzeng and C.H. Yang, 2001. Generation of PCR-based DNA fragments for specific detection of Streptomyces saraceticus N45. Proc. Natl. Sci. Council Roc., 25: 119-127.
Direct Link - Dwivedi, R.K., M.A. Khan, H.P. Singh, D.N. Singh and R.K. Tyagi, 2000. Production Dynamics and fisheries development in Naktara Reservoir, Madhya Pradesh, India. J. Inland Fish. Soc. India, 32: 81-86.
Direct Link - Ugelstad, J., A. Berge, T. Ellingsen, R. Schmid and N. Nilsen et al., 1992. Preparation and application of new monosized polymer particles. Prog. Polym. Sci., 17: 87-161.
Direct Link