
Nadine Adel El-Sebay
Veterinary Serum and Vaccine Research Institute (VSVRI), Cairo, Egypt
LiveDNA: 20.15401
Hala Mohamed Abu Shady
Department of Microbiology, Faculty of Science, Ain Shams University, Cairo, Egypt
Seham Abd El-Rashed El-Zeedy
Veterinary Serum and Vaccine Research Institute (VSVRI), Cairo, Egypt
A.A. Samy
Department of Microbiology and Immunology, Veterinary Division, National Research Center (NRC), Dokki, Egypt
Asian Journal of Scientific Research
Year: 2017 | Volume: 10 | Issue: 3 | Page No.: 194-202







ABSTRACT
Background and Objective: Salmonella typhimurium is one of the most important zoonotic pathogenic microorganisms affecting man and animals, the traditional phenotypical identification of Salmonella involving microbiological enrichment and subsequent identification, usually cannot indicate the genetic determinants of the isolates causing disease. Therefore, the aim of this study was to sequence the invA gene of local isolate which is a very important tool for periodical evaluation of mutagenicity compared with the published sequences on GenBank. Materials and Methods: The collected poultry samples were screened for Salmonella typhimurium using different biochemical and serological studies. The positive isolates were examined by PCR for detection of invA gene at specific molecular size (2058 bp) using specific sensitized primers (forward and reverse). Sequencing of the invA gene was performed and compared with the published sequences on GenBank. Results: The homology percentage of nucleotide sequence (99.2-99.4%) and amino acid sequence (99.6-99.9%) showed high similarity between the local isolates and the other published sequences on GenBank. Conclusion: Sequence analysis of the Egyptian S. typhimurium revealed that it is mandatory to monitor any changes in the genomic structure, similarity and dissimilarity with other strains in the region and all over the world.
PDF Abstract XML References Citation
Received: March 17, 2017;
Accepted: May 16, 2017;
Published: February 23, 2019
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Nadine Adel El-Sebay, Hala Mohamed Abu Shady, Seham Abd El-Rashed El-Zeedy and A.A. Samy, 2017. InvA Gene Sequencing of Salmonella typhimurium Isolated from Egyptian Poultry. Asian Journal of Scientific Research, 10: 194-202.
DOI: 10.3923/ajsr.2017.194.202
URL: https://scialert.net/abstract/?doi=ajsr.2017.194.202
DOI: 10.3923/ajsr.2017.194.202
URL: https://scialert.net/abstract/?doi=ajsr.2017.194.202
INTRODUCTION
Salmonellae are widely distributed worldwide affecting man, animals and birds, it is a direct zoonotic disease of great economic and public health concern, poultry and their products usually considered as one of the principal foodborne sources of Salmonella species that affect humans. S. typhimurium has the most prevalent serotypes isolated from poultry worldwide1,2. Salmonellosis is one of an emerging infectious disease in the USA; it causes more than 450 deaths and 1.2 million infections every year3.
For the effective prevention and control of this foodborne zoonosis, rapid and sensitive detection methods are required, many different conventional media and enrichment regimes have been proposed for isolating Salmonella species from the food and environmental samples2. Although, isolation of Salmonella by growth culture medium followed by serotyping is considered as the gold standard for confirmation of Salmonella, it is also time consuming and labor intensive. Therefore techniques like PCR are increasingly being used for rapid detection and confirmation of Salmonella and considered as a very important diagnostic tool for detection of Salmonella invA targeting gene4. Most widely used DNA based technique is PCR, utilizing genus specific primers targeting various genes. For instance, invA gene (marker gene) has been introduced for the effective, rapid and accurate detection of Salmonella in foods of animal origin5.
The invA gene usually codes for protein in the inner bacterial membrane that is responsible for invasion of intestinal cells of the host6. The invA gene contains unique sequences specific to the genus Salmonella and has been proved as a specific PCR target with important diagnostic applications5,7. The invA target gene of Salmonella is located on the pathogenicity island 1 (SPI-1), it is important for the invasion of host epithelial cells. This gene is highly specific in most Salmonella serotypes and has been used as an important target for detection of Salmonella8,9.This study helps the researcher for periodical monitoring of any changes in the genomic structure of S. typhimurium in Egypt.
The study was planned to fulfill the following:
| • | Detection of invA gene in the local isolates |
| • | Sequencing of invA gene of the most virulence local isolate to detect the homology percentage of the nucleotide and amino acid sequences between local isolate and the published sequences of reference strains and isolates all over the world on GenBank |
MATERIALS AND METHODS
Salmonella isolates: Twelve strains of S. typhimurium were isolated from 7 chickens, 3 chicken eggs and 2 ducks from different geographical areas.
Colonial morphology: A loop full of the obtained samples were inoculated into selenite-F broth and incubated at 37°C for 16 h then streaked on to the surface of MacConkey’s agar, Salmonella Shigella agar, XLD and Hektoen enteric agar then incubated at 37°C for 24 h10.
Microscopical examination: The suspected Salmonella colonies were picked up, a film was made on glass slide and stained with Gram’s stain to examined under ordinary light microscope (National model 138, China)11.
Motility test: Motility was assured by growing the bacteria into semi-solid agar12.
Biochemical identification: The purified isolates of Salmonellae were examined by different biochemical reactions13.
Serological identification of Salmonellae: The isolates preliminary identified biochemically as Salmonella, were subjected to serological identification using diagnostic polyvalent (O, H) and monovalent Salmonella antisera (Difco) for serological identification of Salmonella14.
Polymerase Chain Reaction (PCR) of invA gene: Extraction of Salmonella genome was carried out as recommended by the manufacturer of the Gene JET genomic DNA purification kit (Thermo)15, the PCR was carried out by using the following specific sensitized primers by Primer 3 software (InvA/F and InvA/R) as shown in Table 1.
The amplified products were separated on agarose gel (1.5%) and examined by electrophoresis after staining with ethidium bromide (0.5 mg mL‾1) (Sigma). A 100 bp DNA ladder (Stratagene, USA) was used as a molecular size marker.
| Table 1: | Specific InvA gene primers |
 | |
| F: Forward primer, R: Reverse primer | |
| Table 2: | List of isolates and strain with the corresponding identification and accession numbers on GenBank for sequence analysis |
 | |
Purification of invA fragment of local Salmonella strain: Purification of PCR product for sequence analysis was performed according to the manufacturer instruction using QIA quick gel extraction kit (Qiagen) for extraction of DNA fragment from standard or low-melting agarose gels in TAE buffer (Tris-acetate/EDTA). The kit contains QIA quick spin columns, QG, PE and EB buffers.
Sequence analysis of the DNA fragment: All materials needed for sequence reaction were supplied by the GATC Company, Germany by using (ABI 3730xI DNA) sequencer (Applied Biosystem, USA) and the nucleotide sequencing was done by Sanger sequencing technique16.
Identification of homologies between nucleotide and amino acid sequences of the S. typhimurium isolates published on GenBank was done using BLAST 2.0 and PSI-BLAST search programs, National Center for Biotechnology Information (NCBI). The scores designated in the BLAST search have a well-defined statistical interpretation, making matches easier to distinguish from random background hits. Sequence homology percent and divergence was calculated by MegAlign (DNA STAR, Laser gene, Version 7, USA).
The sequence of the invA gene of local Egyptian isolate was compared with other 10 invA of S. typhimurium strains published sequences on GenBank as shown in Table 2.
RESULTS AND DISCUSSION
Salmonella typhimurium is a leading cause of enteric diseases in humans and animals with millions of illness reported worldwide, therefore, periodical monitoring of any changes in the genomic structure of local S. typhimurium is mandatory. Non-typhoid Salmonella serovars are predominantly associated with animal origin such as eggs, milk, poultry and beef responsible for zoonotic transmission17. Salmonella enteric serovar typhimurium and S. enterica serovar enteritidis are the most frequently encountered species from foods like poultry, pork and beef products18. The incidence of salmonellosis has been reported in many developing countries including India, Egypt, Brazil and Zimbabwe19.
Twelve strains of S. typhimurium isolated from 7 chickens, 3 chicken eggs and 2 ducks from different geographical areas, all examined Salmonella strains were Gram negative, non-spore forming straight rods and highly motile, the isolates were appeared as non-lactose fermenting colonies on MacConkey agar medium. On S.S agar, a colorless colony with black center was formed. On Hekton enteric agar, blue-green colonies with black center was described onto the medium. On Xylose-Lysine Deoxycholate (XLD) agar medium, red colonies with black center were formed. All strains were oxidase negative and H2S positive. Salmonella colonies were tested for serotyping and all 12 isolates are Salmonella typhimurium. These results agree with El-Zeedy et al.20 who isolated 12 isolates from egg samples and 67 isolates from poultry and serotyped as S. rubislaw, S. virginia, S. poona, S. typhimurium, S. montevideo, S. enteritidis, S. infantis, S. cerro, S. agona, S. derby and S. kentucky. The same results obtained by Abdellah et al.21 who recovered S. typhimurium as most predominant Salmonella isolated from contaminated chicken samples. While Abd El-Aziz19 found that the prevalence of S. typhimurium in raw chicken meat, heart and liver was 44, 48 and 40%, respectively. Also, Andino and Hanning22 mentioned that the data from food born outbreak in USA related to human illness showed that serovar enteritidis was the most frequently isolated followed by typhimurium, newport, heidelberg and montevideo by percentage 27, 14, 7, 10 and 3%, respectively. While in the study of Parvej et al.23 the S. enteric serovar typhimurium was isolated and characterized from commercial poultry farms in Bangladesh.
The usefulness of PCR for rapid detection of Salmonella typhimurium from poultry carcasses was proved by many recent studies24,25. Therefore, the polymerase chain reaction (PCR) was performed on genomic DNA extracted from 12 local Salmonella typhimurium isolates for amplification of 2058 bp fragment of invA gene which contains unique sequences to this genus and has been proved as a specific PCR target5,7.
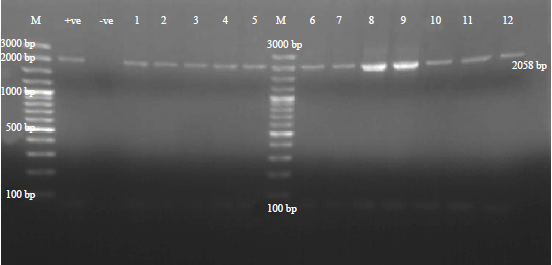 | |
| Fig. 1: | PCR products of 2058 bp fragment of InvA gene |
| Lane M: Gene ruler 100 bp plus DNA ladder marker (Thermo scientific), Lane +ve: ST.LT2 reference positive control, Lane -ve: Negative control, Lane 1-12: Salmonella typhimurium isolates | |
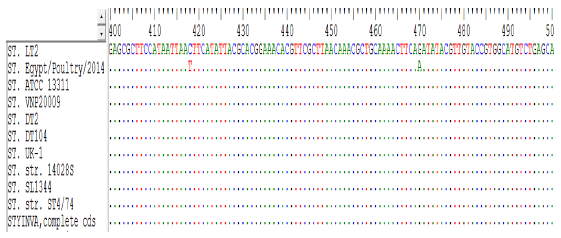 | |
| Fig. 2: | Multiple alignment of nucleotide sequence of invA in Egyptian Salmonella typhimurium strain compared with other 10 invA in Salmonella typhimurium strains published sequences on GenBank |
Electrophoresis of the amplified products on 1.5% agarose gel revealed the presence of specific product at the correct size (2058 bp) according to the standard DNA ladder as shown in Fig. 1.
This finding agreed with Abd El-Tawwab et al.25 who reported that the invA gene was detected in all isolates (100%) from chickens and ducks samples. While Sharma and Das6 found that, the invA gene codes for protein in inner bacterial membrane, which is important for invasion of the host epithelial cells, they detected invA genes in 22 out of 40 Salmonella isolates from poultry meat samples (55%).
Due to invA that encodes a protein in the inner bacterial membrane which is responsible for invasion to the host epithelial cells, the ability of Salmonella specific primers to detect Salmonella species was rapid and accurate9,26 and now has been proved as an international standard for identification of genus Salmonella7.
The nucleotide sequence of 2058 bp PCR fragment representing the invA gene of S. typhimurium isolate was compared with other 10 S. typhimurium strains published sequences on GenBank as shown in Table 2 using Bio Edit software (ClastalW) representing 10 complete sequences of invA gene of different S. typhimurium strains. The different important part of this data is summarized in Fig. 2. After the nucleotide sequence analysis of the invA gene of Salmonella typhimurium local isolate ST. Egypt/Poultry/2014 (ST12) by using the 2058 bp InvA specific primers, the obtained sequence was submitted to GenBank by direct submission using Bankit submission tool under Accession No. (KR185982).
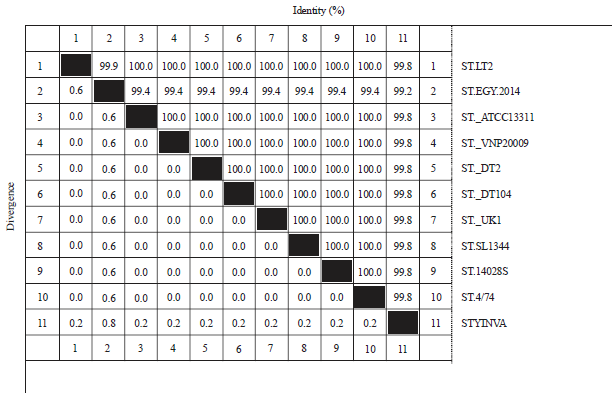 | |
| Fig. 3: | Homology percentage of nucleotide sequence of InvA in Egyptian Salmonella typhimurium strain in comparison with other published Salmonella typhimurium strains |
The obtained sequences of local isolate covered around 2058 bp that encodes 685 deduced amino acid. Multiple alignment revealed nucleotide substitution at the base pair number 470 from the start codon which lead to amino acid substitution from CAG (Serine (Ser/S)) to CAA (Phenylalanine (Phe/F)) at residue number 530, this change is very characteristic and unique for the local isolate than the rest of isolates and strains. Also some different silent nucleotides substitution, non-significant differences, not affected on amino acid substitution occurred in the sequence at the base pairs number: A 208 G, T 418 C, T 589 C, T 1156 A, C 1165 T, A 1288 G, A 1381 G, G 1408 A, G 1654 A, G 1801 A and T 1813C.
The homology percentage of nucleotide sequence results (Fig. 3) showed high homology (99.4%) between ST/Egypt/Poultry/2014 strain and ST.LT2, ST.ATCC13311, ST.VNP20009, ST.DT2, ST.DT104, ST.UK1, ST.SL1344, ST.14028S and ST.4/74 but it gives (99.2%) with STYINVA strain. These results agree with Shi et al.27 who found the high invA homology between Salmonella strains (72.9-97.2%). Also agree with Jarvik et al.28 who found the 14028s and LT2 genomes have more than 98% similarity in sequence and more closely related than any pair of Salmonella strains examined in his studies and in UK-1 strain two unique genes were predicted which are highly homologous to the genes involved in the type III secretion system(as invA gene)29.
The results in this study were parallel also with Edwards et al.30 who described that all of the SPI loci (which contain invA genes) from different serotypes sharing nearly the same distribution of identity as a whole genomes (varying between 97.7 and 98.6% similarities).
The multiple alignment of deduced amino acid (685 amino acid) in Fig. 4 of Egyptian isolate with other published S. typhimurium strains on GenBank showed only one amino acid difference at (F 530 S) and this significant difference led to high identity percentage between S. typhimurium strains. Amino acid substitution from CAG (Serine (Ser/S)), which is considered as a polar amino acid, to CAA (Phenylalanine (Phe/F)), which is a hydrophobic amino acid, at residue number 530. This change is very characteristic and unique for the local isolate ST. Egypt/Poultry/2014 than the rest of published isolates.
The homology percentage of deduced amino acid results (Fig. 5) showed high homology (99.9%) between our Egyptian isolate of ST/Egypt/Poultry/2014 strain and ST.LT2, ST.ATCC13311, ST.VNP20009, ST.DT2, ST.DT104, ST.UK1, ST.SL1344, ST.14028S and ST.4/74 but it gives (99.6%) with STYINVA strain.
The phylogenetic analysis of nucleotide sequence (Fig. 6) showed two major clusters or branches, one representing the Egyptian isolate ST/Egypt/Poultry/2014 and the second divided in two sub-branches, the first for STYINVA strain and the second divided in two small subgroups, the first contains ST.4/74 and ST.SL1344 strains and the second contains ST.LT2, ST.ATCC13311, ST.VNP20009, ST.DT2, ST.DT104, ST.UK1 and ST.14028S strains.
 | |
| Fig. 4: | Multiple alignment of deduced amino acid sequence of InvA in Egyptian Salmonella typhimurium strain compared with other 10 InvA in Salmonella typhimurium strains published sequences on GenBank |
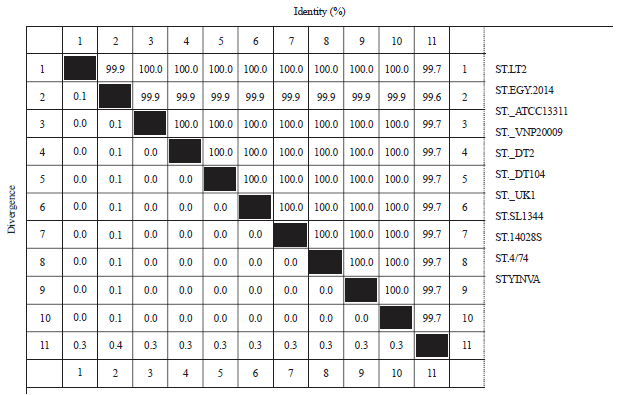 | |
| Fig. 5: | Homology percentage of deduced amino acid sequence of InvA in Egyptian Salmonella typhimurium strain in comparison with other published Salmonella typhimurium strains |
Therefore, ST. Egypt/Poultry/2014 is unique in their position with other Salmonella typhimurium strains.
The phylogenetic analysis of deduced amino acid (Fig. 7) showed two major clusters or branches on rooted tree, one representing the STYINVA (M90846.1) strain and the second one divided in two sub-branches, the first for ST/Egypt/Poultry/2014 Egyptian local strain (KR185982) and the second for remaining strains: ST.LT2 (AE006468.1), ST.ATCC13311 (CP009102.1), ST.VNP20009 (CP007804.2), ST.DT2 (HG326213.1), ST.DT104 (HF937208.1), ST.UK1 (CP002614.1), ST.SL1344 (CP001363.1), ST.14028S (FQ312003.1) and ST.4/74 (CP002487.1). Therefore, ST/Egypt/Poultry/2014 is unique in their position with other Salmonella typhimurium strains.
Both phylogenetic trees (Fig. 6, 7) demonstrated that the Egyptian isolate (ST/Egypt/Poultry/2014) is closely related to the other S. typhimurium isolates but in a separate cluster because the presence of single, significant amino acid substitution at position (F 530 S), due to the change of nucleotide at position (A 470 G) and non-significant changes in another nucleotides.
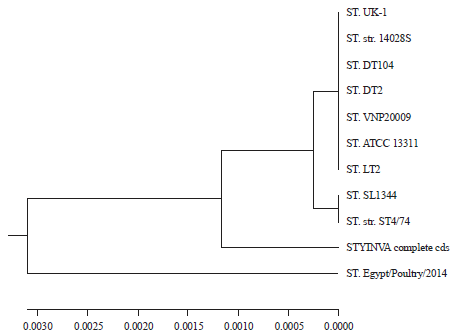 | |
| Fig. 6: | Phylogenetic tree of nucleotide sequence of InvA gene of Egyptian Salmonella typhimurium strain with other published Salmonella typhimurium strains |
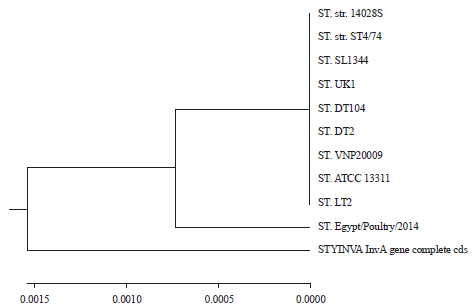 | |
| Fig. 7: | Phylogenetic tree of deduced amino acid sequence of InvA gene of Egyptian Salmonella typhimurium with other published Salmonella typhimurium strains |
These results agree with Kingsley et al.31 who found the isolates of DT2 formed a distinct phylogenetic cluster within Salmonella typhimurium serotypes. Comparative genomic analysis of DT2 94-213 and S. typhimurium SL1344, D23580 and DT104 identified few differences in genetic content with the exception of variations within prophages. Also, in this study found that the STYINVA strain was in a separate cluster in both phylogenetic trees due to some amino acid substitutions when comparing with other Salmonella strains, this differences may be due to long years lasts from isolation and sequencing of this strain science 199232, while the other strains (including Egyptian isolate) were isolated and sequenced from 2001 until 2014, which may explains the minor differences them.
All these results of invA alignment agree with Galan and Curtiss33 and Shi et al.27 who said that the invA is present and functional in most, if not all, virulent Salmonella strains and its conserved gene.
CONCLUSION
Regular monitoring of the Egyptian S. typhimurium sequence changes is an important discriminatory marker that could be used in epidemiological investigations as well as during vaccine preparation and evaluation.
ACKNOWLEDGMENT
We would like to express our great thanks to the staff members of Genetic Engineering Department, VSVRI, Agricultural Research Center, Ministry of Agriculture, Egypt for providing support for this study.
SIGNIFICANCE STATEMENT
This study discovered the changes in invA gene sequence between local Egyptian S. typhimurium strain and other published strains on GenBank. These changes should be taken in consideration during manufacturing and evaluation of new the S. typhimurium vaccine.
REFERENCES
- EFSA. and ECDPC., 2013. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2011. EFSA J., Vol. 11.
CrossRefDirect Link - Scallan, E., R.M. Hoekstra, F.J. Angulo, R.V. Tauxe and M.A. Widdowson et al., 2011. Foodborne illness acquired in the United States-major pathogens. Emerg. Infect. Dis., 17: 7-15.
CrossRefDirect Link - Li, R., Y. Wang, J. Shen and C. Wu, 2013. Development of a novel hexa-plex PCR method for identification and serotyping of Salmonella species. Foodborne Pathogens Dis., 11: 75-77.
CrossRefDirect Link - Rahn, K., S.A. De Grandis, R.C. Clarke, S.A. McEwen and J.E. Galan et al., 1992. Amplification of an invA gene sequence of Salmonella typhimurium by polymerase chain reaction as a specific method of detection of Salmonella. Mol. Cell. Probes., 6: 271-279.
CrossRefPubMedDirect Link - Sharma, I. and K. Das, 2016. Detection of InvA gene in isolated Salmonella from marketed poultry meat by PCR assay. J. Food Process Technol., Vol. 7.
CrossRefDirect Link - Malorny, B., J. Hoorfar, C. Bunge and R. Helmuth, 2003. Multicenter validation of the analytical accuracy of Salmonella PCR: towards an international standard. Applied Environ. Microbiol., 69: 290-296.
CrossRefDirect Link - Harper, J. and K.F. Shortridge, 1969. A selective motility medium for routine isolation of Salmonella. J. Hyg. (Lond.), 67: 181-186.
Direct Link - Sanger, F. and A.R. Coulson, 1975. A rapid method for determining sequences in DNA by primed synthesis with DNA polymerase. J. Mol. Biol., 94: 441-446.
CrossRefDirect Link - Alvarez-Fernandez, E., C. Alonso-Calleja, C. Garcia-Fernandez and R. Capita, 2012. Prevalence and antimicrobial resistance of Salmonella serotypes isolated from poultry in Spain: comparison between 1993 and 2006. Int. J. Food Microbiol., 153: 281-287.
CrossRefDirect Link - Vose, D., T. Koupeev and K. Mintiens, 2013. A quantitative microbiological risk assessment of Salmonella spp. in broiler (Gallus gallus) meat production. EFSA Supporting Publ., Vol. 8.
CrossRefDirect Link - Abd El-Aziz, D.M., 2013. Detection of Salmonella typhimurium in retail chicken meat and chicken giblets. Asian Pac. J. Trop. Biomed., 3: 678-681.
CrossRefDirect Link - Abdellah, C., R.F. Fouzia, C. Abdelkader, S. Rachida and Z. Mouloud, 2009. Prevalence and anti-microbial susceptibility of Salmonella isolates from chicken carcasses and giblets in Meknes, Morocco. Afr. J. Microbiol. Res., 3: 215-219.
Direct Link - Andino, A. and I. Hanning, 2015. Salmonella enterica: Survival, colonization and virulence differences among serovars. Scient. World J.
CrossRefDirect Link - Parvej, S., M. Rahman, F. Uddin, N.H. Nazir, S. Jowel, F.R. Khan and B. Rahman, 2016. Isolation and characterization of Salmonella enterica Serovar typhimurium circulating among healthy chickens of Bangladesh. Turk. J. Agric. Food Sci. Technol., 4: 519-523.
Direct Link - Jamshidi, A., M.R. Bassami and S. Afshari-Nic, 2008. Identification of Salmonella spp. and Salmonella typhimurium by a multiplex PCR-based assay from poultry carcasses in Mashhad, Iran. Int. J. Vet. Res., 3: 43-48.
Direct Link - Shanmugasamy, M., T. Velayutham and J. Rajeswar, 2011. Inv A gene specific PCR for detection of Salmonella from broilers. Vet. World, 4: 562-564.
Direct Link - Shi, Q., Y. Zhang, Q.Y. Wang, G. Gao and H. Fang et al., 2012. Phylogenetic analysis of virulence factor gene of Salmonella isolated from clinically symptomatic chickens. Afr. J. Microbiol. Res., 6: 1718-1722.
Direct Link - Jarvik, T., C. Smillie, E.A. Groisman and H. Ochman, 2010. Short-term signatures of evolutionary change in the Salmonella enteric serovar typhimurium 14028 genome. J. Bacteriol., 192: 560-567.
CrossRefPubMedDirect Link - Luo, Y., Q. Kong, J. Yang, G. Golden and S. Wanda et al., 2011. Complete genome sequence of the universal killer Salmonella enterica serovar typhimurium UK-1 (ATCC 68169). J. Bacteriol., 193: 4035-4036.
CrossRefDirect Link - Edwards, R.A., G.J. Olsen and S.R. Maloy, 2002. Comparative genomics of closely related Salmonellae. Trends Microbiol., 10: 94-99.
CrossRefPubMedDirect Link - Kingsley, R.A., S. Kay, T. Connor, L. Barquist and L. Sait et al., 2013. Genome and transcriptome adaptation accompanying emergence of the definitive type 2 host-restricted Salmonella enterica serovar typhimurium pathovar. mBio, Vol. 4.
CrossRefDirect Link - Galan, J.E., C. Ginocchio and P. Costeas, 1992. Molecular and functional characterization of the Salmonella invasion gene i3vA: Homology of invA to members of a new protein family. J. Bacteriol., 174: 4338-4349.
Direct Link - Galan, J.E. and R. Curtiss, 1991. Distribution of InvA,-B, -C and -D genes of Salmonella typhimurium among other Salmonella serovars: InvA mutants of Salmonella typhi are deficient for entry into mammalian cells. Infect. Immun., 59: 2901-2908.
Direct Link