
Lourdusamy Arul Antony Maria
Centre for Nanotechnology and Advanced Biomaterials (CeNTAB), School of Chemical and Biotechnology, SASTRA University, 613401, Thanjavur, India
Vidya N. Chamundeswari
Centre for Nanotechnology and Advanced Biomaterials (CeNTAB), School of Chemical and Biotechnology, SASTRA University, 613401, Thanjavur, India
Meera Parthasarathy
Centre for Nanotechnology and Advanced Biomaterials (CeNTAB), School of Chemical and Biotechnology, SASTRA University, 613401, Thanjavur, India
Asian Journal of Scientific Research
Year: 2014 | Volume: 7 | Issue: 3 | Page No.: 303-311







ABSTRACT
Electrophoretic separation of membrane proteins is limited mainly by their intrinsic hydrophobicity and poor solubility arising from association with membrane lipid components. In this investigation, polyacrylamide gel matrix is made hydrophobic by introducing one-dimensional and two-dimensional carbon nanomaterials viz., multi-walled carbon nanotubes and graphene. The nanocomposite matrices were optimized for SDS-PAGE with water-soluble molecular weight marker proteins. SDS-PAGE in the nanocomposite matrices of a commercial outer membrane porin protein, OmpA indicated a significant decrease in anomalous migration of the protein with increasing carbon nanotube loading in the gel matrix. Outer membrane proteins of Escherichia coli, were isolated and characterized by Fourier Transform Infrared Spectroscopy. When the nanocomposite gels were tested for electrophoretic separation of outer membrane proteins isolated from E. coli, the resolution of protein bands improved with respect to the pristine polyacrylamide gel. Especially, graphene/polyacrylamide composite hydrogels yielded far better resolution and faster migration of the E. coli membrane proteins compared to multi-walled carbon nanotube/polyacrylamide composite hydrogels. Based on contact angle measurements of the composite hydrogels, the improved resolution of membrane proteins is attributed to the more hydrophobic environment rendered by carbon nanotubes and graphene. The present results could be useful to develop more hydrophobic nanocomposite gels exclusively for electrophoretic separation of membrane proteins isolated from different sources.
PDF Abstract XML References Citation
Received: September 16, 2013;
Accepted: November 05, 2013;
Published: April 10, 2014
How to cite this article
Lourdusamy Arul Antony Maria, Vidya N. Chamundeswari and Meera Parthasarathy, 2014. Lotus-leaf Inspired Hydrophobic Nanocomposite Matrices for Electrophoretic Separation of Bacterial Outer Membrane Proteins. Asian Journal of Scientific Research, 7: 303-311.
DOI: 10.3923/ajsr.2014.303.311
URL: https://scialert.net/abstract/?doi=ajsr.2014.303.311
DOI: 10.3923/ajsr.2014.303.311
URL: https://scialert.net/abstract/?doi=ajsr.2014.303.311
INTRODUCTION
Membrane proteins are the key components of cell membranes responsible for a range of structural, regulatory, transportation, energy utilization and enzymatic functions. They constitute about 30% of total protein content in a single cell and serve as targets for about 50% of commercial drugs (Handbook from GE healthcare). In addition to their physiological significance, membrane proteins are also interesting candidates for fundamental research owing to the unique properties of the individual proteins especially in terms of detergent binding, folding/unfolding kinetics, amphipathic structure and charge heterogeneity. Despite their medical and fundamental significance, structure and function of individual membrane proteins are less understood compared to water soluble proteins as evident from the meager number of high resolution crystal structures resolved so far (Newstead et al., 2008; Bill et al., 2011). One of the major difficulties in the purification, handling and analysis of membrane proteins, especially the trans-membrane or integral proteins, is the poor solubility in aqueous medium owing to their hydrophobic nature, other difficulties being low natural abundance in cell membrane and association with other membrane components (Seddon et al., 2004).
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) is a versatile technique used to separate proteins based on their molecular weight differences (Chrambach and Rodbard, 1971). The success of SDS-PAGE relies on the expectation that all proteins in the analyte have nearly equal detergent loading and adopt similar shapes after SDS treatment as that of the molecular weight marker proteins used for calibration (Chrambach and Rodbard, 1971). However, this requirement is not satisfied usually by membrane proteins due to unpredictable SDS loading (3.4-10 g SDS g-1 protein) arising from differences in hydrophobicity and tertiary structure of the individual proteins (Rath et al., 2009). For this reason, SDS-PAGE is being used only as a rapid screening method for sorting out membrane proteins based on differences in their tertiary/quaternary structure and not to identify membrane proteins based on their electrophoretic migration. Only a few reports are available on improving membrane protein electrophoresis in SDS-PAGE. For instance, anomalous gel shift of membrane proteins in SDS-PAGE was reduced by 13% using polyleucine molecules synthesized with a range of molecular weights as hydrophobic molecular weight markers to mimic the abundance of leucine residues in the helical trans-membrane proteins (Rath et al., 2010). While the above report demonstrated the efficacy of polyleucine standards for sieving trans-membrane proteins in the molecular weight range of 3.5-41 kDa, extension of the protocol to higher molecular weights requires synthesis of appropriate polyleucine standards.
A more feasible approach to address the issue is the modification of the polyacrylamide gel matrix itself. A noteworthy attempt in this connection is the identification of 37 new membrane proteins from the human colon carcinoma HCT-116 cells using a more hydrophobic gel matrix (Tokarski et al., 2011). Nevertheless, caution must be exercised while improving the hydrophobicity of gel matrix without compensating its porosity and homogeneous polymerization. For instance in the above report, issues due to limited choice of %T and scattered migration of certain proteins arising from inhomogeneities in polymerization in addition to longer polymerization time has been noticed when N-alkylated and N, N’-dialkylated monomers were mixed with the regular gel composition for Laemmli’s PAGE. In this regard, introduction of carbon nanotubes into the gel matrix has been identified as an alternative option to introduce hydrophobicity to the matrix without compromising its properties (Gunavadhi et al., 2012; Parthasarathy et al., 2011; Jiang et al., 2010; Huang et al., 2006; Chen and Hsieh, 2010). We have demonstrated earlier that introduction of multi-walled carbon nanotubes in the gel matrix improves the molecular weight calibration plots in SDS-PAGE (Parthasarathy et al., 2011). We have also shown that multi-walled carbon nanotubes themselves can act as catalysts for free radical polymerization of acrylamide monomers yielding nanotube-grafted polyacrylamide hydrogels with more uniform pore size compared to pristine polyacrylamide hydrogels, further improving the molecular weight calibration in SDS-PAGE (Gunavadhi et al., 2012).
In this study, we demonstrate the applicability of Polyacrylamide/Carbon Nanotube and Polyacrylamide/Graphene composite hydrogels to the electrophoretic separation of outer membrane proteins extracted from Escherichia coli. The nanocomposite gels were analyzed using a commercial bacterial outer membrane porin from E. coli (OmpA) and systematic investigation has been carried out at different stages of membrane protein extraction. Thus the present report provides a new dimension to the electrophoretic separation of bacterial membrane proteins using carbon nanomaterial matrices made of carbon nanotubes and graphene.
MATERIALS AND METHODS
Materials: Multi-walled carbon nanotubes (about 20 nm diameter, 5-15 microns length, 95% purity) were procured from Nanostructured and Amorphous materials Inc., Houston, Texas, USA and used as such without further purification. Acrylamide for electrophoresis, Acetic acid glacial 99-100% for synthesis,Tris buffer GR (hydroxy methyl aminomethane), Sodium dodecyl sulphate (sodium lauryl sulphate), Silver nitrate pure, Ammonium peroxodisulfate extra pure (Ammonium per sulphate), Sodium carbonate anhydrous pure, Formaldehyde solution 37-41% w/v, Glycerol about 87% GR, sodium chloride cryst. pure, Potassium chloride GR, Potassium dihydrogen Phosphate Purified, Disodium hydrogen Phosphate dihydrate GR were procured from Merck, India. Sodium thiosulfate and glycine were purchased from Qualigens and Themo Fisher Scientific India Pvt. Ltd. 2-mercapto ethanol, bromophenol blue AR were procured from SD Fine chemicals Ltd., India, TEMED from Sigma Aldrich, China, Outer membrane protein-A OmpA, a bacterial recombinant from Prospec Protein Specialists, Israel, Bisacrylamide (N,N’-methylene bisacrylamide Ultrapure for molecular biology), Nutrient agar, Nutrient Broth, Agar Agar from Himedia India, E. coli strain from Microbial type Culture Collection and Gene Bank MTCC, India and Deionized water (18.2 MÙ cm) from Siemens Ultra clear Water Purification System. Graphene was prepared by ultrasonication of a high purity graphite rod in the monomer solution containing the surfactant (SDS) and then polymerization is carried out resulting in the formation of homogeneous graphene/polyacrylamide (G-PAM) composite hydrogels.
METHODS
Extraction of membrane protein from gram negative bacteria Escherichia coli: Nutrient agar plates were prepared and E. coli from freeze dried sample was streaked and kept in incubator overnight. From the overnight grown plate individual colony was isolated and inoculated in nutrient broth and kept in shaker overnight at 120 rpm at 37°C. This inoculum was transferred to centrifuge tubes. Cells were harvested by centrifugation at 6000 rpm for 15 min and the supernatant was discarded. Thirty millilitter of ice cold (PBS) phosphate buffer saline (wash buffer NaCl 8 g L-1, KCl 0.2 g L-1, Na2HPO4.2H2O 1.44 g L-1, KH2PO4 0.24 g L-1, pH 7.4) was added to the pellet. The pellet was resuspended, centrifuged again at 6000 rpm for 15 min and the supernatant was discarded. The 10 mL of ice cold PBS was added to the pellet and resuspended. The cell suspension was sonicated for 2 min and centrifuged at 2000 rpm for 5 min and the supernatant containing the cytoplasmic fractions was discarded. The 10 mL of SDS solution (above critical micelle concentration) is added to the pellet and sonicated again. This contains cell membrane components along with integral membrane proteins which when loaded into the gel for electrophoresis, floats over the buffer like an oil droplet. So the mixture was gently agitated after sonication for 30 min in a shaker and centrifuged again at 2000 rpm for 5 min and the pellet containing lipid components of the cell membrane get discarded and the supernatant was collected. Though the supernatant contains fewer amounts of cell membrane lipid components, the protein bands obtained after SDS-PAGE were feeble and the sample aggregated with time due to the presence of SDS. The supernatant was centrifuged at 18000 rpm for 1 h and the pellet contains the crude extract of membrane proteins which was stored in freezer at -20°C for SDS-PAGE. The pellet was characterized by a Perkin-Elmer Model Fourier Transform Infrared Spectrophotometer (FTIR).
SDS-PAGE in composite gels: Multi-walled carbon nanotube/polyacrylamide composite gels were prepared with different loading of nanotubes (0.001, 0.0025, 0.005, 0.01% w/v) as reported earlier (Parthasarathy et al., 2011). Briefly, calculated amounts of pristine multi-walled carbon nanotubes were dispersed by ultrasonication in the gel buffer solution for Laemmli’s SDS-PAGE at pH 8.8, containing the monomer and cross-linker, followed by the addition of TEMED (10 μL) and APS (0.01% w/v) (Laemmli, 1970). Photographs of CNT/PAM composite gels and SEM image of multi-walled carbon nanotubes are shown in supporting information. Stacking gel composition consisted of 1 M Tris adjusted to pH 6.8 using 2N HCl. Electrophoresis (SDS-PAGE) was performed for molecular weight marker proteins (14-97 kDa) with a loading of 7 μg per well at 30°C at an applied d.c. voltage of 100 V and 20 mA current. The run was monitored by the mobility of the tracking dye (Bromophenol blue) present in the protein mixture and each run took 2-2.5 h. The gels were placed in fixer for 24 h and then subjected to silver staining.
Graphene/polyacrylamide composite gels were prepared by ultrasonication of a high purity graphite rod in the monomer solution containing the surfactant (SDS) and then polymerization is carried out resulting in the formation of homogeneous graphene/polyacrylamide (G-PAM) composite hydrogels. Formation of monolayer graphene was confirmed by Transmission Electron Microscopy (JEOL) and X-ray diffraction of the dried nanocomposite gels. SDS-PAGE and visualization conditions were the same as mentioned above.
RESULTS AND DISCUSSION
Figure 1 shows the FTIR spectrum of membrane protein mixture extracted from E. coli. The FTIR absorption at 1654 cm-1 indicates the predominance of α helical conformation in the crude extract of membrane proteins (Haris and Severcan, 1999; Kong and Yu, 2007). The presence of a strong C-N, N-H stretching band at 1223 cm-1 is characteristic of the Amide III band. After characterizing by FTIR, the crude membrane protein extract was subjected to SDS-PAGE.
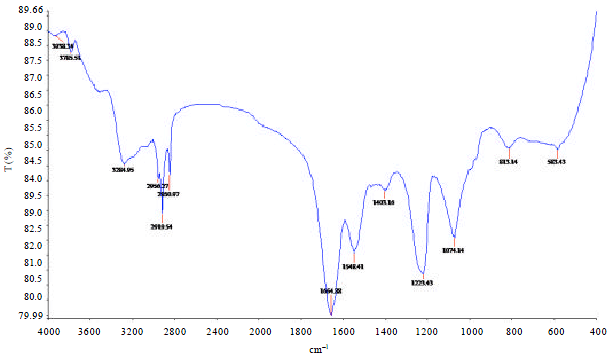 | |
| Fig. 1: | FTIR spectrum of crude membrane proteins extracted from E. coli |
 | |
| Fig. 2(a-b): | Comparing the electrophoretic mobility of OmpA and molecular weight marker proteins in (a) Pristine PAM, 0.005 wt% CNT/PAM, 0.01 wt% CNT/PAM, (b) Shifting of OmpA band with respect to 45 kDa molecular weight marker protein with varying CNT content in polyacrylamide gel |
Electrophoresis of the membrane protein extract in gel matrices of 12 and 15% T pristine polyacrylamide. The protein bands were observed too close at the lower molecular weight region in the 12% T gel while a better migration range was obtained with the 15% T gel due to the smaller pore size that helps in the better separation of proteins ranging from high to low molecular weight. Hence 15% T gel was used as the benchmark for the rest of the investigation. However, in both 12 and 15% T, the membrane protein bands were found to be curved towards the anode.
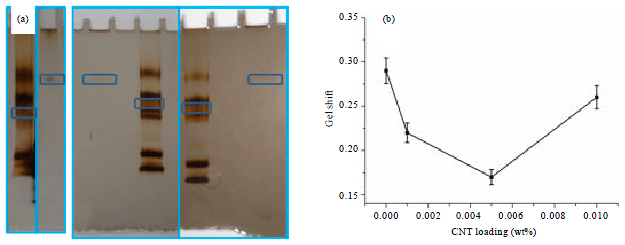
To verify the effect of loading CNTs into the gel matrix on the migration pattern of membrane proteins, we have compared the migration of a commercial outer membrane porin from E. coli (OmpA: M.Wt. 48 kDa) with water-soluble molecular weight marker proteins (Fig. 2). Generally membrane proteins run faster than expected either due to incomplete denaturation or due to binding of more SDS, resulting in a positive error in the apparent molecular weight determined by SDS PAGE (Handbook from GE healthcare). However, porins are known to migrate slower than expected based on their real molecular weight, as they bind less amount of SDS to achieve near-complete denaturation despite their heat-modification behavior (Arcidiacono et al., 2002; Doig and Trust, 1994). The results obtained here are in accordance with this. Interestingly, ‘gel shift’ of the 48 kDa commercial protein, OmpA with respect to the 45 kDa marker protein is found to decrease with increase in CNT loading from 0.001-0.005 wt%. This indicates that membrane proteins move faster in CNT/PAM gels than in pristine PAM gels which could be explained using a mechanism similar to that reported for the case of hydrophobic alkylated polyacrylamide gels as follows (Gunavadhi et al., 2012). The composite gel matrix consists of SDS-bound multi-walled carbon nanotubes as well as SDS in the gel buffer. Due to certain level of hydrophobicity of pristine (unfunctionalized) carbon nanotubes the dodecyl chains of SDS will be lying on the nanotube walls while the sulfate groups could be exposed. This is verified by contact angle measurements on dry CNT/PAM composite gels: Pristine PAM gels (contact angle 40°) immediately absorb the water droplet while CNT/PAM gels (contact angle 95°) absorb water only slowly.
 | |
| Fig. 3(a-e): | Electrophoregrams of membrane proteins extracted from E. coli in 15% T, (a) pristine Polyacrylamide, (b) 0.001% CNT/PAM, (c) 0.002% CNT/PAM, (d) 0.005% CNT/PAM and (e) 0.01% CNT/PAM |
As a result, sulfate groups exposed on the surface of SDS-bound membrane proteins could slide past sulfate groups exposed on the surface of pores due to higher electrostatic repulsion between the negatively charged sulfate groups compared to the case of hydrophilic polyacrylamide gel matrices. Nevertheless, at higher CNT loading viz., 0.01 wt%, gel shift is found to increase, probably due to the increasing heterogeneity in the matrix with increasing CNT content.
Having compared the electrophoretic behavior of OmpA in CNT/PAM and PAM gels, the next objective of the investigation was to analyze mixtures of outer membrane proteins extracted from E. coli. Electrophoresis of the membrane protein extract was performed in gel matrices of 12 and 15% T pristine polyacrylamide. The protein bands were more resolved in 15% T gel due to the smaller pore size that helps in the better separation of proteins ranging from high to low molecular weight. Hence 15% T gel was used as the benchmark for the rest of the investigation. Accordingly, Fig. 3 shows the electrophoregrams of crude membrane protein extract from E. coli in 15% T pristine PAM compared with 15% T CNT/PAM composite gels with varying loading of CNTs. In all the cases, a dark band is observed at the loading point which could be due to the precipitation of membrane-associated proteins with high lipid content. However, when electrophoresis is continued the precipitates re-dissolve and enter the gel, leading to a continuous dark background of unresolved proteins. In the present report, such a dark background is observed prominently in the case of pristine PAM which surprisingly diminishes with increasing CNT loading in the matrix. This could be attributed to better re-dissolution of lipid-associated proteins in the more hydrophobic CNT/PAM gel matrices. Moreover, the membrane proteins are also found to be more resolved with increasing CNT loading as evident from the increasing separation between two prominent bands encircled in Fig. 3.
After analyzing CNT/PAM composite gels, experiments were repeated with graphene/PAM composite gels. Graphene/PAM hydrogels prepared in the study are more hydrophobic (contact angle 120°) compared to CNT/PAM and hence membrane proteins are expected to be more resolved in these matrices. Formation of monolayer graphene after ultrasonication was confirmed by Transmission Electron Microscopy and electron diffraction of the dry dispersion (Fig. 4). Also, the X-ray diffraction pattern of the dry composite hydrogel shows two peaks at 2-theta values 22 and 24° corresponding to d-values, 0.41 and 0.37 nm respectively, close to the interlayer spacing in graphite (0.35 nm) (Fig. 4).
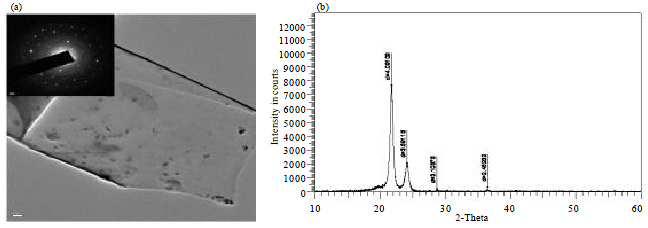 | |
| Fig. 4(a-b): | (a) Transmission electron micrograph and (b) X-ray diffraction pattern of graphene synthesized by ultrasonication of graphite |
 | |
| Fig. 5(a-c): | Silver-stained SDS-PAGE electrophoregrams of (a) Water-soluble molecular weight marker proteins in 12% T pristine PAM (left) and 12% T Graphene/PAM (right), (b) bacterial membrane proteins in 12% T pristine PAM (left) and 12% T Graphene/PAM (right); (c) Bacterial membrane proteins in 15% T pristine PAM (left) and 15% T Graphene/PAM (right) |
First, water soluble molecular weight marker proteins were analyzed by SDS-PAGE in the Graphene/12% T PAM composite gels and compared with pristine 12% T PAM hydrogel. Accordingly, in the electrophoregrams in Fig. 5a no distinct bands are observed in Graphene/PAM composite gel due to the poor retention of the water-soluble proteins in the highly hydrophobic matrix. On the other hand, drastic improvement in resolution and migration rate was observed when the bacterial membrane protein mixture was loaded in the Graphene/PAM gel both in 12% T (Fig. 5b) and 15% T (Fig. 5c) gel composition compared to the respective pristine PAM gels. Graphene/PAM composite gels yield far better results than CNT/PAM gels not only in terms of resolution but also in terms of faster migration probably resulting from good electrical conductivity of the graphene filler. Electrical conductivity of the composite gel, however, could not be measured because the filler resided deep inside the hydrogel and were not accessible to the electrical contacts on the surface.
CONCLUSION
In summary, the effect of adding multi-walled carbon nanotubes and graphene into polyacrylamide gel matrix on the electrophoretic separation of bacterial outer membrane proteins has been investigated. Commercial outer membrane porin, OmpA, from E. coli when loaded in CNT/PAM indicated a significant decrease in anomalous migration behaviour with increasing CNT loading in the gel matrix matrix from 0.001-0.005 wt%. This is explained using a conceptual model based on the exposure of SDS molecules on the surface of proteins and gel pore walls. Outer membrane protein mixture extracted from E.coli, when loaded in CNT/PAM and Graphene/PAM yielded well-resolved bands compared to pristine polyacrylamide gel. Based on contact angle measurements performed on the pristine and composite hydrogels and in the light of the proposed model, the improved resolution of membrane proteins in the composite hydrogels is attributed to the increase in hydrophobicity of the gel matrix. In addition, background staining which normally arises due to poor dissolution of lipid-associated membrane proteins in polyacrylamide matrix is observed to decrease after introducing the nanomaterials in the gel matrix with the appearance of more distinct bands compared to pristine polyacrylamide gel. Graphene/Polyacrylamide composite gels enhance the overall migration rate of the proteins thereby reducing the duration of electrophoresis. This could be due to the enhanced electrical conductivity of the hydrogel due to the presence of grapheme. Thus the present investigation paves way to the development of novel nanocomposite hydrophobic hydrogels exclusively for electrophoretic separation of membrane proteins. The results could be easily extended to the separation and purification of membrane proteins isolated from different sources and organisms.
ACKNOWLEDGMENTS
Authors acknowledge financial support from the Department of Science and Technology, India under Fast Track Scheme for Young Scientists (No. SR/FT/CS-121/2010), Nanomission Council-PG Teaching Grant (No. SR/NM/PG-IG/2007), Department of Science and Technology, New Delhi for providing infrastructural facilities and SASTRA University for providing financial support under Prof. T. R. Rajagopalan Research Funds Scheme.
REFERENCES
- Newstead, S., J. Hobbs, D. Jordan, E.P. Carpenter and S. Iwata, 2008. Insights into outer membrane protein crystallization. Mol. Membrane Biol., 25: 631-638.
CrossRef - Bill, R.M., P.J. Henderson, S. Iwata, E.R. Kunji and H. Michel et al., 2011. Overcoming barriers to membrane protein structure determination. Nat. Biotechnol., 29: 335-340.
CrossRef - Seddon, A.M., P. Curnow and P.J. Booth, 2004. Membrane proteins, lipids and detergents: Not just a soap opera. Biochim. Biophys. Acta, 1666: 105-117.
CrossRef - Chrambach, A. and D. Rodbard, 1971. Polyacrylamide gel electrophoresis. Science, 172: 440-451.
CrossRef - Rath, A., M. Glibowicka, V.G. Nadeau, G. Chen and C.M. Deber, 2009. Detergent binding explains anomalous SDS-PAGE migration of membrane proteins. Proc. Natl. Acad. Sci., 106: 1760-1765.
CrossRef - Rath, A., V.G. Nadeau, B.E. Poulsen, D.P. Ng and C.M. Deber, 2010. Novel hydrophobic standards for membrane protein molecular weight determinations via sodium dodecyl sulphate-polyacrylamide gel electrophoresis. Biochemistry, 49: 10589-10591.
CrossRef - Tokarski, C., M. Fillet and C. Rolando, 2011. Improved gel electrophoresis matrix for hydrophobic protein separation and identification. Anal. Biochem., 410: 98-109.
CrossRef - Gunavadhi, M., L.A.A. Maria, V.N. Chamundeswari and M. Parthasarathy, 2012. Nanotube-grafted polyacrylamide hydrogels for electrophoretic protein separation. Electrophoresis, 33: 1271-1275.
CrossRef - Parthasarathy, M., J. Debgupta, B. Kakade, A.A. Ansary, M. Islam Khan and V.K. Pillai, 2011. Carbon nanotube-modified sodium dodecyl sulphate-polyacrylamide gel electrophoresis for molecular weight determination of proteins. Anal. Biochem., 409: 230-235.
CrossRef - Jiang, F., Y. Wang, X. Hu, N. Shao, N. Na, J.R. Delanghe and J. Ouyang, 2010. Carbon nanotubes‐assisted polyacrylamide gel electrophoresis for enhanced separation of human serum proteins and application in liverish diagnosis. J. Separation Sci., 33: 3393-3399.
CrossRef - Huang, G., Y. Zhang, J. Ouyang, W.R. Baeyens and J.R. Delanghe, 2006. Application of carbon nanotube-matrix assistant native polyacrylamide gel electrophoresis to the separation of apolipoprotein AI and complement C3. Anal. Chim. Acta, 557: 137-145.
CrossRef - Chen, J.L. and K.H. Hsieh, 2010. Polyacrylamide grafted on multi‐walled carbon nanotubes for open-tubular capillary electrochromatography: Comparison with silica hydride and polyacrylate phase matrices. Electrophoresis, 31: 3937-3948.
CrossRef - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Haris, P.I. and F. Severcan, 1999. FTIR spectroscopic characterization of protein structure in aqueous and non-aqueous media. J. Mol. Catal. B, 7: 207-221.
CrossRef - Kong, J. and S. Yu, 2007. Fourier transform infrared spectroscopic analysis of protein secondary structures. Acta Biochimica Biophysica Sinica, 39: 549-559.
CrossRefDirect Link