
T.A.A. Mohammed
National Institute of Oceanography and Fisheries, Red Sea Branch, Hurghada, Egypt
M.H. Yassien
National Institute of Oceanography and Fisheries, Suez Branch, Egypt
Asian Journal of Scientific Research
Year: 2013 | Volume: 6 | Issue: 1 | Page No.: 98-106







ABSTRACT
The crab size, abundance, distribution and density of two gall crabs species (Hapalocarcinus marsupialis Stimpson, 1859 and Opecarcinus aurantius Kropp, 1989) associated with the most abundant and common coral species have been studied in 5 sites along the Egyptian Red Sea coast. Three hundred and forty five coral galls were collected; 276 coral galls were found in Stylophora pistillata and 59 galls in Pocillopora verrucosa. The mean size of coral galls is ranged between 1.42±0.37 cm and 2.17±0.62 cm for S. pistillata (at Qula'an and Hurghada, respectively), while the size of galls in P. verrucosa is ranged between 1.35±0.19 cm and 1.48±0.17 cm (at Hamata and Hurghada, respectively). S. pistillata showed the highest gall crab numbers per colony (10.43 galls colony-1) at Hurghada, while P. verrucosa recorded the highest number (3.83 galls colony-1) at Qula'an site. S. pistillata recorded the highest coral galls density (63 with a mean of 9±3.32) and crab density (20.59% with mean 3±2.6) at Hurghada site. P. verrucosa recorded the optimum galls number (23 with mean of 3.8±1.5) and crab density (6.86% and mean of 1.3±1.7) at Qula'an.
PDF Abstract XML References Citation
Received: March 23, 2012;
Accepted: May 23, 2012;
Published: October 06, 2012
How to cite this article
T.A.A. Mohammed and M.H. Yassien, 2013. Assemblages of Two Gall Crabs within Coral Species Northern Red Sea, Egypt. Asian Journal of Scientific Research, 6: 98-106.
DOI: 10.3923/ajsr.2013.98.106
URL: https://scialert.net/abstract/?doi=ajsr.2013.98.106
DOI: 10.3923/ajsr.2013.98.106
URL: https://scialert.net/abstract/?doi=ajsr.2013.98.106
INTRODUCTION
Many authors have recorded and observed the associated animals within the living corals as crabs (Cantera et al., 2003; Kotb and Hartnoll, 2002), Bivalves (Mohammed and Yassien, 2008) and sponges (Palpandi et al., 2007). The living corals provide the potential symbionts not only food but also the suitable habitats that allowing greater diversity of symbiotic modes of life than do many other types of host (Patton, 1976; Oigman-Pszczol and Creed, 2006).These associated fauna were attached mainly by passive-endoliths that embed themselves in the live tissue and allow the coral skeleton to grow up around them (Scoffin and Bradshaw, 2000). Mohammed and Yassien (2008) illustrated that, coral reefs are one of the most important substrate, where they are associated as a major type of hard substrate characterized by a living surface especially among mollusks. Generally, the crab distribution and diversity were studied by many authors such as Soundarapandian et al. (2008) in Pichavaram mangrove environment of India and Rozihan and Ismail (2011) in the west coast of Peninsular Malaysia. Soundarapandian et al. (2008) illustrated that, the families Grapsidae and Ocypodidae are the most dominant forms.
Gall-crabs associated with living corals forming galls, tunnels and/or pits inside the coral skeleton and feed on their mucus and tissues (Kropp, 1986, 1990; Ross and Newman, 2000), whereas the coral reefs provide suitable substrate and habitat for their growth and development (Rinkevich et al., 1991; Carricart-Ganivet et al., 2004). The later author pointed out that, the gall crabs consume deposited materials or algae on expected coral skeleton as a commensals relation. Kotb and Hartnoll (2002) studied the reproduction of gall crab Hapalocarcinus marsupialis in Stylophora pistillata and divided the coral galls into five stages, where the later stages contained larger and matured crabs and had a high crab production of ovigerous females.
Hartnoll (2006) mentioned that, the physiological state of the gall crabs declines the growth rates and may cause mortality increasing due to feed reduction or the reproduction diversion. Johnsson et al. (2006) studied two other cryptochiridae associated with Siderastrea stellata and pointed out that these species inhabit adjoining cavities of their colonies at Brazilian. Moreover, the salinity degree is a major controlling factor for egg hatching and survival at, where the optimum degree is ranged between 25-35 ppt (Mia et al., 2001; Zaleha et al., 2011).
This study is aimed to determine the assemblages, abundance and distribution of gall crabs on some branching coral species. On the other hand identifying the collected crabs and determining their density on the associated coral species pointing out the coral percentage cover and the different impacts at the studied sites.
MATERIALS AND METHODS
Five stations were chosen, from north to middle of the Egyptian Red Sea coast (Fig. 1), to study the assemblages of gall crabs inside some coral species indicating to the relationship between them. The first station is Ras El-Behar (RB) that lies at 60 km north of Hurghada (at 27°43'45"N-27°43'51"N and 33°32'58"E-33°33'04"E); the second site is facing Hurghada (at 27°17'13"N and 33°46'43"E); the third site is Hamata which lies south Hurghada (at 24°38'52"N and 35°05'54"E); the fourth site is Qula'an (at 24°21'35"N and 35°17'47"E) and the fifth is Shlateen (at 23°09'10"N and 35°36'58"E). Samples were collected during a period from September 2009 to December 2010 from the mentioned sites using snorkeling and SCUBA diving equipments. However, the collected crabs were found only within the branching corals Stylophora pistillata Esper and Pocillopora verrucosa at depths ranged between 0.5 and 2 m, whenever, the collected crabs were taken only from the complete and closed coral galls. Galls were randomly collected using long-nose pliers to break off the branches with galls at their ends, without otherwise damaging the colonies as it possible.
The samples were preserved in a mixture of formalin, alcohol and glycerine. Laboratory examination and measurements of coral galls and crabs number were carried out within 7-15 days to reduce their color change or any changes in body size contents (Kotb and Hartnoll, 2002). The width of coral galls were also measured using vernier caliper. The ranges of Carapace Length (CL) and Carapace Width (CW) were also calculated in mm. The abundance and density of gall crabs were determined as well as the percentage cover of the associated coral species using line intercept transect (20 m long). The collected data were analyzed using SPSS ver. 11, soft ware for windows.
The measurements of coral galls width are based only on 33 complete samples of coral galls and are calculated in cm. while crab carapace are measured in mm. One-way ANOVA for homogeneity of variance was used for data analysis to test the null hypothesis of no differences in the infestation density between sites and the collected coral species.
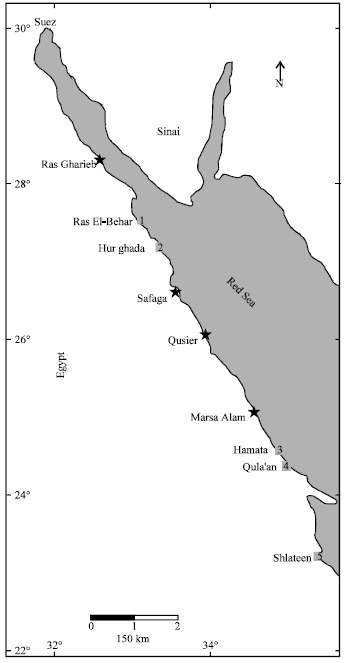 | |
| Fig. 1: | The location map of the studied sites (from 1 to 5) |
RESULTS
During the present investigation two different crab species were collected and identified as Hapalocarcinus marsupialis Stimpson (1859) and Opecarcinus aurantius Kropp (1989).
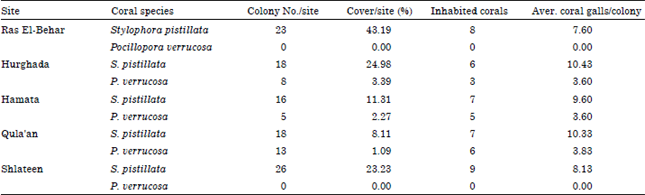
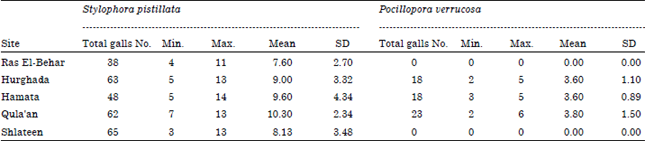
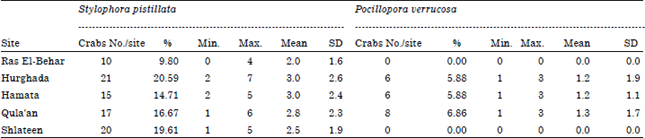
Densities of coral galls and gall crabs: During the present study, a total of 345 coral galls were observed and calculated either found in a closed and complete stage or in incomplete gall stage on the five selected sites. About 276 coral galls were found in the Stylophora pistillata colonies and 59 galls were observed in Pocillopora verrucosa. The recorded number of inhabited coral colonies with gall crabs are ranged between 6 at Hurghada to 9 colonies at Shlateen site (Table 1) for Stylophora pistillata; while the colony number of Pocillopora verrucosa was ranged between 3 colonies at Hurghada and 6 colonies at Qula'an and not recorded in Ras El-Behar and Shlateen. Moreover, the percentage cover of the inhabited coral Stylophora pistillata recorded its maximum value (43.19%) at Ras El-Behar, while the coverage percentage of Pocillopora verrucosa is relatively low and recorded its highest value (3.39%) at Hurghada. Table 1 illustrating also the percentage cover of the inhabited coral species at the studied sites, where S. pistillata recorded the highest value of coral galls/colony (7.6-10.30 galls). Moreover, the density of the collected coral galls was illustrated in Table 2, where Shlateen recorded the highest number of coral galls (65 galls with a mean of 8.13±3.48) followed by Hurghada (63 galls with a mean of 9.0±3.32) for Stylophora pistillata; while Qula'an recorded the maximum coral galls number for Pocillopora verrucosa (23 with mean of 3.8±1.5). On the other hand, the density of gall crabs (±SD) in the closed and complete coral galls only recorded the maximum number (21 crabs representing 20.59% with a mean of 3.0±2.6) at Hurghada (Table 3) followed by Shlateen (20 gall crabs representing 19.61% and a mean of 2.5±1.9) for Stylophora pistillata; while Qula'an recorded the maximum number of gall crabs (8 gall crabs representing only 6.86% and a mean of 1.3) for Pocillopora verrucosa (Table 3).
| Table 1: | No. of coral colonies, percentage cover and average No. of coral galls/colony (aver.) at the different sites |
 | |
| Table 2: | Coral gall densities (±SD) and total No. of the inhabited species at the studied sites |
 | |
| Table 3: | Gall crab densities of the complete and closed coral galls of the two coral species at the studied sites |
 | |
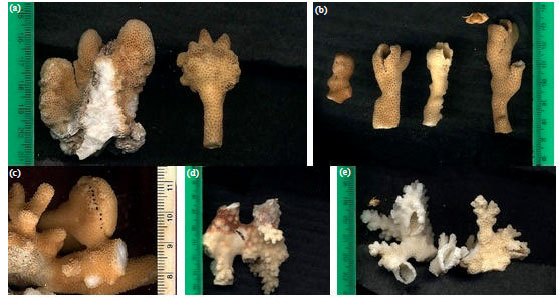 | |
| Fig. 2(a-e): | The crab induced coral galls of (a-c) Stylophora pistillata colonies and (d-e) Pocillopora verrucosa colonies |
| Table 4: | Mean sizes of the collected coral galls of Stylophora pistillata and Pocillopora verrucosa |
 | |
| Table 5: | The carapace ranges of length and width of the two collected gall crabs |
 | |
Sizes of coral galls and crabs carapace: The mean size of coral galls (based on 5-8 samples at least for each coral species at each site) is ranged between 1.42±0.37 cm and 2.17±0.62 cm (Table 4) for Stylophora pistillata (at Qula'an and Hurghada respectively); while the size of the galls in Pocillopora verrucosa is ranged between 1.35±0.19 cm and 1.48±0.17 cm (at Hamata and Hurghada, respectively). Whoever, the measurements of these galls is mainly depended on the closed coral galls for the collected two coral species (Fig. 2a-e), where, the carapace width of the collected crab (CW) is varied from 2.75-5.25 mm and the Carapace Length (CL) is varied from 3.75-5.25 mm for Hapalocarcinus marsupialis. While the CW of Opecarcinus aurantius is ranged between 3.91 and 5.25 mm, while the CL is ranged between 4.15 and 6.25 mm (Table 5).
Crabs and coral species association: It was observed that, the collected crab species are closely associated with the inhabited coral species where, O. aurantius (Fig. 3a-c) is closely associated in the galls of P. verrucosa only.
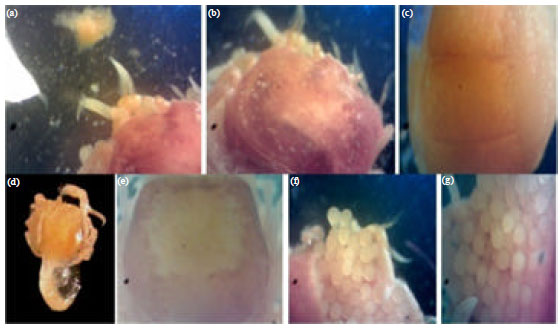 | |
| Fig. 3(a-g): | The collected gall crabs (a-c) Opecarcinus aurantius closely associated to P. verrucosa (d-e) Hapalocarcinus marsupialis associated with S. pistillata and (f-g) Hapalocarcinus marsupialis with fully matured eggs |
| Table 6: | One-way ANOVA of the mean coral gall sizes |
 | |
On the other hand, H. marsupialis (Fig. 3d-e) is closely associated with S. pistillata only and never collected from P. verrucosa. By comparing the two crab densities in the two coral species, it was observed that, H. marsupialis has a higher density than O. aurantius at the studied sites. While the crab number (the present crabs in the complete and closed coral galls) is ranged between 10 and 21 crab/site (at Ras El-Behar and Hurghada, respectively) for S. pistillata; while the density of O. aurantius is relatively low and ranged between 6 and 8 crabs/site for P. verrucosa. Some samples of H. marsupialis were found in a full and complete egg maturation (Fig. 3f-g) during collection in the spring and summer seasons. On the other hand, O. aurantius was not observed nor collected in a mature stage of gonads.
One-way ANOVA of the sizes of coral galls at the studied sites illustrated that, there is no significantly differences between groups of the site but there is a significantly differences in the mean sizes of the coral galls at p = 0.036 and f = 6.37 (Table 6).
DISCUSSION
Some symbiotic crabs such as families cryptochiridae and Trapeziidae have been suggested as dependant on coral hosts for protection against predators and as food source produced by the coral mucus or particulate materials (Preston, 1973; Reed et al., 1982; Abelson et al., 1991; Simon-Blecher et al., 1999; Carricart-Ganivet et al., 2004; Johnsson et al., 2006). Family Chiridae includes a small group of cryptic crabs that inhabit cavities of scleractinian corals forming galls. Where, the formed galls derived from coral growth modifications and are likely to be associated with the alimentary mechanisms of the crabs (Carricart-Ganivet et al., 2004). The studied crab species are belonging to families cryptochiridae and Trapeziidae. Only one crab per gall in each colony was recorded in all coral samples of the studied sites. The present study illustrated that Hapalocarcinus marsupialis was associated only with Stylophora pistillata as described by Kotb and Hartnoll (2002) and Opecarcinus aurantius was associated only with Pocillopora verrucosa. O. aurantius has specific interrelation with coral species P. verrucosa. Each gall contains either the male or female crab only as showed by Johnsson et al. (2006).
The high cover of S. pistillata (8.11-43.19%) in the different sites relative to P. verrucosa illustrated the dominancy of H. marsupialis than O. aurantius. Hurghada recorded the highest crab density in S. pistillata while the lowest crab density in Qula'an attributed to the low coral cover due to the flash floods. The branch sizes (or thickness) and the crab sizes in Stylophora is relatively lower than in Pocillopora that may cause crab density increasing in Stylophora than in Pocillopora. Oigman-Pszczol and Creed (2006) and Kropp and Manning (1987) found that, the small crabs are found abundantly in a large number of coral hosts. Also, the ability of these crabs to hide and live in galls or depressions of corals with a symbiotic relation.
The crabs infested S. pistillata more than P. verrucosa may be attributed to the coral host characteristics as the low skeletal density of S. pistillata, vertical growth (Young and Christoffersen, 1984) and due to the defense mechanisms against stinging nematocysts. Association of gall crabs tends to live in the most locally abundant coral species within their potential host assemblage and diversity (Sin and Lee, 2000; Cantera et al., 2003). The increased density and abundance of associated crabs with host does not affect the live and growth of corals and create simple holes parallel the growth structure. The commensal coral-crab association and the local environmental conditions are the controlling factors for the crab assemblages and the larval settlement stages of crabs on the coral hosts. Moreover, the coral morphology and number of branches can provide many niches for associations of these assemblages (Patton, 1974, 1994; Goh et al., 1999; Oigman-Pszczol and Creed, 2006).
One-way ANOVA illustrated a significantly differences in the mean sizes of the coral galls at p = 0.036 and f = 6.37, whenever, this is related to the small branch sizes and the higher covers in S. pistillata which caused increasing crab number and density.
CONCLUSION
There are more than one species of crab (Hapalocarcinus marsupialis Stimpson (1859) and Opecarcinus aurantius Kropp (1989) that associated and inhabited some coral species as Stylophora pistillata and Pocillopora verrucosa forming galls.
Numbers and densities of gall crabs are associated with the percentage cover and density of the coral host and the completely formed coral galls. The increased density and abundance of crabs with host does not cause any impact on the coral live and allow them grow normally.
Stylophora pistillata has a higher density and assemblages of gall crabs at Hurghada and Shlateen (recorded the maximum covers) than Pocillopora verrucosa which recorded it maximum density at Qula'an.
The size of gall crab Opecarcinus aurantius is larger than that of Hapalocarcinus marsupialis. Moreover, O. aurantius is closely associated with galls of P. verrucosa only and never collected from S. pistillata which contains only the crab H. marsupialis.
REFERENCES
- Cantera, K., R. Jaime, C. Orozco, E. Londono-Cruz and G. Toro-Farmer, 2003. Abundance and distribution patterns of infaunal associates and macroborers of the branched coralb (Pocillopora damicornis) in Gorgona Island (Eastern tropical Pacific). Bull. Mar. Sci., 72: 207-219.
Direct Link - Carricart-Ganivet, J.P., L.F. Carrera-Parra, L. Quan-Young and M.S. Garcia-Madrigal, 2004. Ecological note on Troglocarcinus corallicola (Brachyura: Cryptochiridae) living in symbiosis with Manicina areolata (Cnidaria: Scleractinia) in the Mexican caribbean. Coral Reefs, 23: 215-217.
CrossRef - Goh, N.K.C., P.K.L. Ng and L.M. Chou, 1999. Notes on the shallow water gorgonian-associated fauna on coral reefs in Singapore. Bull. Mar. Sci., 65: 259-282.
Direct Link - Johnsson, R., E. Neves, G.M.O. Franco and F.L. da Silveira, 2006. The association of two gall crabs (brachyura: Cryptochiridae) with the reef-building coral Siderastrea stellata verrill, 1868. Hydrobiologia, 559: 379-384.
CrossRefDirect Link - Kotb, M.M.A. and R.G. Hartnoll, 2002. Aspects of the growth and reproduction of the coral gall crab Hapalocarcinus marsupialis. J. Crustacean Biol., 22: 558-566.
Direct Link - Kropp, R.K., 1986. Feeding biology and mouthpart morphology of three species of coral gall crabs (Decapoda: Cryptochiridae). J. Crustacean BioI., 6: 377-384.
Direct Link - Kropp, R.K., 1990. Revision of the genera of gall crabs (Crustacea: Cryptochiridae) occurring in the Pacific Ocean. Pacific Sci., 44: 417-448.
Direct Link - Mia, M.Y., S. Shokita and N. Shikatani, 2001. Early life history of estuarine grapsid crab, Helice tridens latimera. Pakistan J. Biol. Sci., 4: 1437-1439.
CrossRefDirect Link - Mohammed, T.A. and M.H. Yassien, 2008. Bivalve assemblages on living coral species in the Northern Red Sea, Egypt. J. Shellfish Res., 27: 1217-1223.
CrossRef - Oigman-Pszczol, S.S. and J.C. Creed, 2006. Distribution and abundance of fauna on living tissues of two Brazilian Hermatypic corals (Mussismilia hispida (Verril 1902) and siderastrea stellata verril, 1868). Hydrobiologia, 563: 143-154.
CrossRef - Palpandi, C., G. Ananthan and A. Shanmugam, 2007. Cryptofaunal association of Spirastrella inconstans (Dendy) and S. inconstans var. digitat (Dendy), the coral reef sponges of palk bay. J. Fish. Aqut. Sci., 2: 71-76.
CrossRef - Patton, W.K., 1994. Distribution and ecology of animals associated with branching corals (Acropora spp) from the great-barrier-reef Australia. Bull. Mar. Sci., 55: 193-211.
Direct Link - Preston, E.M., 1973. A computer simulation of competition among five sympatric congeneric species of xanthid crabs. Ecology, 54: 469-483.
Direct Link - Rozihan, M. and E. Ismail, 2011. Genetic structure and Haplotype diversity of Tachypleus gigas population along the West Coast of Peninsular Malaysia-inferred through mtDNA at rich region sequence analysis. Biotechnology, 10: 298-302.
CrossRefDirect Link - Reed, J.K., R.H. Gore, L.E. Scotto and K.A. Wilson, 1982. Community composition, structure, areal and trophic relationships of decapods associated with shallow-and deep-water oculina varicosa coral reefs: Studies on decapod crustacea from the Indian river region of Florida. Bull. Mar. Sci., 32: 761-786.
Direct Link - Rinkevich, B., Z. Wolodarsky and Y. Loya, 1991. Coral-crab association: A compact domain of a multilevel trophic system. Hydrobiologia, 216-217: 279-284.
CrossRefDirect Link - Scoffin, T.P. and C. Bradshaw, 2000. The taphonomic significance of endoliths in dead-versus live-coral skeletons. PALAIOS, 15: 248-254.
Direct Link - Simon-Blecher N., A. Chemedanov, N. Eden and Y. Achituv, 1999. Pit structure and trophic relationship of the coral pit crab Cryptochirus coralliodytes. Mar. Biol., 134: 711-717.
CrossRef - Soundarapandian, P., N.J. Samuel, S. Ravichandran and T. Kannupandi, 2008. Biodiversity of crabs in pichavaram mangrove environment, South East Coast of India. Int. J. Zool. Res., 4: 113-118.
CrossRefDirect Link - Young, P.S. and M.L. Christoffersen, 1984. Recent coral barnacles of the genus ceratoconcha (cirripedia: Pyrgomatidae) from Northeast Brazil. Bull. Mar. Sci., 35: 239-252.
Direct Link - Zaleha, K., I. Hazwani, H.S. Hamidah, B.Y. Kamaruzzaman and K.C.A. Jalal, 2011. Effect of salinity on the egg hatching and early larvae of horseshoe crab Tachypleus gigas (Muller, 1785) in laboratory culture. J. Applied Sci., 11: 2620-2626.
CrossRefDirect Link