
L.C. Saalu
Department of Anatomy, Lagos State University College of Medicine (LASUCOM), Ikeja, Lagos, Nigeria
A.A. Osinubi
Department of Anatomy, College of Medicine, University of Lagos, Lagos, Nigeria
P.I. Jewo
Department of Anatomy, Lagos State University College of Medicine (LASUCOM), Ikeja, Lagos, Nigeria
A.O. Oyewopo
Department of Anatomy, College of Medicine, University of Ilorin, Ilorin, Nigeria
G.O. Ajayi
Department of Medical Biochemistry, Lagos State University College of Medicine (LASUCOM), Ikeja, Lagos, Nigeria
Asian Journal of Scientific Research
Year: 2010 | Volume: 3 | Issue: 1 | Page No.: 51-61







ABSTRACT
The therapeutic value of Doxorubicin (DOX) as anticancer antibiotic is limited by its organotoxicity. It has been shown that free radicals are involved in doxorubicin-induced toxicity. Doxorubicin causes the generation of free radicals and the induction of oxidative stress, associated with cellular injury. Because of the great importance of DOX in cancer therapy, researchers have expended great efforts trying to prevent or attenuate the side effects of DOX administration. There has, however, been only a little success in this regard. In this study the ameliorating role of antioxidant-rich ethanolic seed extract of Citrus paradisi (CP) on DOX-induced testicular oxidative stress and impaired sperm parameters was investigated. Three experimental groups of Wistar rats were used; CP-alone group that received orally CP 10 mg kg-1 b.wt. daily for 14 days followed by intraperitoneal (i.p.) Normal Saline (NS) 2.5 mg kg-1 b.wt. DOX-alone group that had ip DOX 10 mg kg-1 b.wt. as a single dose. CP plus DOX-group that were similarly given DOX, but also had oral CP 10 mg kg-1 b.wt. pretreatment for 14 days. There was also a corresponding control group. The animals were autopsied 8 weeks after DOX or NS injections. Results showed that DOX-induced reduction in sperm motility and epididymal sperm concentrations as well as increase in total abnormal sperm rates were all normalized in the group pretreated with CP. Pretreatment with CP ameliorated the testicular content of Glutathione (GSH) and Auperoxide Dismutase (SOD), Catalase (CAT) and Glutathione Peroxidase (GPx) activities. Similarly, CP treatment attenuated the DOX-induced increase in testicular lipid peroxidation reflected by malondialdehyde (MDA) levels. These data indicate that CP protects the rat testis against DOX-induced oxidative stress and deranged sperm characteristics.
PDF Abstract XML References Citation
How to cite this article
L.C. Saalu, A.A. Osinubi, P.I. Jewo, A.O. Oyewopo and G.O. Ajayi, 2010. An Evaluation of Influence of Citrus paradisi Seed Extract on Doxorubicin-Induced
Testicular Oxidative Stress and Impaired Spermatogenesis. Asian Journal of Scientific Research, 3: 51-61.
DOI: 10.3923/ajsr.2010.51.61
URL: https://scialert.net/abstract/?doi=ajsr.2010.51.61
DOI: 10.3923/ajsr.2010.51.61
URL: https://scialert.net/abstract/?doi=ajsr.2010.51.61
INTRODUCTION
Doxorubicin (DOX) is an anthracycline antibiotic and cytotoxic (antineoplastic) agent. Anthracycline antibiotics are among the most effective and commonly used anticancer drugs (Priestman, 2008). It is commonly used against ovarian, breast, lung, uterine and cervical cancers, Hodgkin’s disease, soft tissue and primary bone sarcomas, as well against in several other cancer types (Bonadonna et al., 1996; Atessahin et al., 2006). Unfortunately, their clinical use is restricted by dose-dependent toxicity (Yeh et al., 2007; Berthiaume and Wallace, 2007; Saalu et al., 2009a). In particular the occurrence of infertility following treatment with this anticancer drug is a serious concern (Suominen et al., 2003; Prahalathan et al., 2005).
The biochemical mechanism by which DOX causes cytotoxicity is currently unclear. Several mechanisms have been postulated to account for the effects of DOX, both in terms of anticancer potential and adverse effects. It is however thought to have three major activities that vary with the type of cell:
The drug once it penetrates into the cell binds to chromatin. It then forms a complex with the DNA by intercalation of its planar rings between nucleotide base pairs (Mornparler et al., 1976). The planar aromatic chromophore portion of the molecule intercalates between two base pairs of the DNA, while the six-membered daunosamine sugar sits in the minor groove and interacts with flanking base pairs immediately adjacent to the intercalation site, as evidenced by several crystal structures (Pigram et al., 1972; Ferderick et al., 1990).
It is also widely accepted that DOX-induced organopathy resides for the most part on oxidative stress and the production of free radicals (Quiles et al., 2002; Chularojmontri et al., 2005; Prahalathan et al., 2005; Saalu et al., 2009b). DOX is known to generate free radicals either by the enzymatic pathway of redox cycling between a semiquinone form and a quinone form or by the non-enzymatic pathway of forming a DOX-Fe3+ complex (Davies and Doroshow, 1986; Armstrong and Lipsy, 1993). In both pathways, molecular oxygen is reduced to superoxide anion (O2•), which is converted to other forms of reactive oxygen species such as hydrogen peroxide (H2O2) and the more toxic hydroxyl radical (OH•). These free radicals could then cause membrane and macromolecule damage by three basic mechanisms; lipid peroxidation, deoxyribonucleic acid (DNA) fragmentation and protein oxidation (Pacher, 2007).
Because of the great importance of DOX in cancer therapy, researchers have expended great efforts trying to prevent or attenuate the side effects of DOX administration. Accordingly, several approaches have been pursued, such as dosage optimization, synthesis and use of analogues or combined therapy (Singh, 2005; Jain, 2005; Banerjee and Verma, 2008; Wolinsky and Grinstaff, 2008). No hopeful results have been found and the application of different DOX-analogues did not show better antineoplastic value or lower toxicity than DOX (Bromberg, 2008). The production of free radicals as a by product of DOX metabolism is considered to be the primary mechanism of DOX-induced toxicity, which suggests some new approaches, such as the potential use of natural antioxidants (Quiles et al., 2002).
The Citrus paradisi (CP) popularly called grapefruit is a tropical and subtropical citrus tree grown for its fruit which was originally named the forbidden fruit of Barbados (Dowling and Morton, 1987). The CP seed extract contains high levels of Vitamin C, Vitamin E and the polyphenolic flavonone glycosides hesperidin, neohesperidin, nariratin and naringin (Halliwell, 2008). These compounds are powerful antioxidants individually and collectively. CP seed extract has therefore since been regarded as one of the most powerful natural antioxidants (Sachs, 1997; Armando et al., 1997).
The concept that DOX-induced testicular injury is principally mediated through the oxidative pathway and that CP seed extract possesses potential antioxidative effect by scavenging free radicals, has prompted us to address in the current study the possible testiculoprotective effects of this nutraceutical on testicular oxidative stress and impaired spermatogenesis in Wistar rats challenged with DOX.
MATERIALS AND METHODS
Chemicals
Doxorubicin hydrochloride (®Adricin, Korea United Pharm. Inc., Chungnam, Korea) was obtained from Juli Pharmacy, Ikeja, Lagos State, Nigeria in the month of May, 2008.
Plant Material and its Extraction
In the first week of May, 2008, fresh parts of Citrus paradisi (CP) tree were collected from a cultivated farmland within the deciduous forest of Odorasanyi District, Ijebu-Igbo, Ogun State, Nigeria. The plant material was collected at this site because earlier collected plant material at same site which was identified and authenticated at the Forestry Research Institute of Nigeria (FRIN), Ibadan where a voucher specimen with herbarium number FHI 107460 was deposited. Plant taxonomy was done by Mr. T. K. Odewo, Chief Superintendent Officer, Taxonomy Section, FRIN, Ibadan, Oyo State, Nigeria. Plant authentication was done by Dr. A.B. Kadiri, The Herbarium, Botany and Microbiology Department, the University of Lagos, Akoka, Lagos State, Nigeria.
Twenty eight CP fruits were cut into pieces and the seeds were separated out. These were thoroughly but gently rinsed in distilled water. The seeds were completely dried at room temperature for three weeks. The air dried seeds weighing 140 g were reduced to a powdered substance by grinding. The sample was exhaustively extracted with 99.8% ethanol (BDH scientific supplies Ltd, Lagos, Nigeria) by means of a Soxhlet apparatus and the extract was evaporated in vacuo. The residue was processed to give 10 g (7.5% yield) of dark yellowish solid crude mass, which was stored at 4°C for the study. Fresh solution of the extract was prepared in peanut oil as vehicle when required. The concentration of the extract in peanut oil was 5 mg mL-1.
Phytochemical Analysis of Plant Material
Phytochemical screenings were performed.
Animals and Interventions
Experimental procedures involving the animals and their care were conducted in conformity with International, National and institutional guidelines for the care of laboratory animals in Biomedical Research and Use of Laboratory Animals in Biomedical Research as promulgated by the Canadian Council of Animal Care (1985). Further, the animal experimental models used conformed to the guiding principles for research involving animals as recommended by the Declaration of Helsinki and the Guiding Principles in the Care and Use of Animals (American Physiological Society, 2002).
The rats were procured from a breeding stock maintained in the Animal House of Lagos State University College of Medicine (LASUCOM). The animals were housed in well ventilated wire-wooden cages in the Animal Facility of the Department of Anatomy, LASUCOM, Ikeja, Lagos. An approval was sought and obtained from the Departmental ad-hoc Ethical Committee. The rats were maintained under standard natural photoperiodic condition of twelve hours of light alternating with twelve hours of darkness (i.e., L: D; 12:12) with room temperature of between 25 to 26°C and humidity of 65±5%. They were allowed unrestricted access to water and rat chow (Feedwell Livestock Feeds Ltd., Ikorodu, Lagos, Nigeria). They were allowed to acclimatize for 28 days before the commencement of the experiments. The weights of the animals were estimated at procurement, during acclimatization, at commencement of the experiments and twice within a week throughout the duration of the experiment, using an electronic analytical and precision balance (BA210S, d = 0.0001 g) (Satorius GA, Goettingen, Germany).
Forty male adult (11 to 13 weeks old) Wistar rats weighing 185-210 g were used for this research work. The rats were randomly divided into four groups of ten rats each such that the average weight difference between and within groups did not exceed ±20% of the average weight of the sample population. The control group of rats were each given 2.5 mL kg-1 b.wt. peanut oil daily by gastric gavage for 2 weeks. Then 24 h after, they were given 2.5 mL kg-1 b.wt. normal saline as a single dose ip. They were then sacrificed 8 weeks after, the duration of spermatogenesis in rat being about 8 weeks (Heller and Clermont, 1964; Jegou et al., 2002). The CP group of animals were each given reference dose (Heggers et al., 2002). Then 24 h after, they were given a single dose normal saline 2.5 mL kg-1 b.wt. i.p. and then they were sacrificed 8 weeks after normal saline administration.
The DOX group of rats were each given 2.5 mL kg-1 b.wt. peanut oil by gastric gavage daily for 2 weeks. Then the next day they were given 10 mg kg-1 b.wt. DOX intraperitoneally (i.p.) as a single dose. This dosage being well documented to cause testicular damage in rats (Atessahin et al., 2006). The rats were then sacrificed 8 weeks after DOX administration. The CP+DOX group of animals each received 10 mg kg-1 b.wt. 99.8% ethanol extract of CP kg-1 b.wt. by gastric gavage daily for 2 weeks. Then 24 h after they were given 10 mg kg-1 b.wt. DOX i.p. as a single dose and were sacrificed 8 weeks after.
Animal Sacrifice and Sample Collection
The rats were at the time of sacrifice first weighed and then anaesthesized by placing them in a closed jar containing cotton wool sucked with chloroform anaesthesia. The abdominal cavity was opened up through a midline abdominal incision to expose the reproductive organs. Then the testes were excised and trimmed of all fat. The testes weights of each animal were evaluated .The testes were weighed with an electronic analytical and precision balance (BA 210S, d = 0.0001-Sartoriusen GA, Goettingen, Germany). The testes volumes were measured by water displacement method. The two testes of each rat were measured and the average value obtained for each of the two parameters was regarded as one observation.
Sperm Characteristics
The testes from each rat were carefully exposed and removed. They were trimmed free of the epididymides and adjoining tissues.
Epididymal Sperm Concentration
Spermatozoa in the right epididymis were counted by a modified method of Yokoi and Mayi (2004). Briefly, the epididymis was minced with anatomic scizzors in 5 mL physiologic saline, placed in a rocker for 10 min and allowed to incubate at room temperature for 2 min. After incubation, the supernatant fluid was diluted 1:100 with solution containing 5 g sodium bicarbonate and 1 mL formalin (35%). Total sperm number was determined by using the new improved Neuber`s counting chamber (haemocytometer). Approximately, 10 μL of the diluted sperm suspension was transferred to each counting chamber of the haemocytometer and was allowed to stand for 5 min. This chamber was then placed under a binocular light microscope using an adjustable light source. The ruled part of the chamber was then focused and the number of spermatozoa counted in five 16-celled squares. The sperm concentration was then calculated multiplied by 5 and expressed as [X]x106 mL-1, where [X] is the number of spermatozoa in a 16-celled square.
Sperm Progressive Motility
This was evaluated by an earlier method by Sonmez et al. (2005). The fluid obtained from the left cauda epididymis with a pipette was diluted to 0.5 mL with Tris buffer solution. A slide was placed on light microscope with heater table, an aliqout of this solution was on the slide and percentage motility was evaluated visually at a magnification of x400. Motility estimates were performed from three different fields in each sample. The mean of the three estimations was used as the final motility score. Samples for motility evaluation were stored at 35°C.
Sperm Morphology
The sperm cells were evaluated with the aid of light microscope at x 400 magnification. Caudal sperm were taken from the original dilution for motility and diluted 1:20 with 10% neutral buffered formalin (Sigma-Aldrich, Oakville, ON, Canada). Five hundred sperm from the sample were scored for morphological abnormalities (Atessahin et al., 2006). Briefly, in wet preparations using phase-contrast optics, spermatozoa were categorized. In this study a spermatozoon was considered abnormal morphologically if it had one or more of the following features: rudimentary tail, round head and detached head and was expressed as a percentage of morphologically normal sperm.
Assay of Testicular Enzymatic Antioxidants
Assay of Catalase (CAT) Activity
Catalase activity was measured according to the method of Aebi (1983). The 0.1 mL of the testicular homogenate (supernatant) was pipetted into cuvette containing 1.9 mL of 50 mM phosphate buffer, pH 7.0. Reaction was started by the addition of 1.0 mL of freshly prepared 30% (v/v) hydrogen peroxide (H2O2). The rate of decomposition of H2O2 was measured spectrophotometrically from changes in absorbance at 240 nm. Activity of enzyme was expressed as μ mg-1 protein.
Assay of Superoxide Dismutase (SOD) Activity
Superoxide dismutase activity was measured according to the method of Winterbourn et al. (1975) as described by Rukmini et al. (2004). The principle of the assay was based on the ability of SOD to inhibit the reduction of nitro-blue tetrazolium (NBT). Briefly, the reaction mixture contained 2.7 mL of 0.067 M phosphate buffer, pH 7.8, 0.05 mL of 0.12 mM riboflavin, 0.1 mL of 1.5 mM NBT, 0.05 mL of 0.01 M methionine and 0.1 mL of enzyme samples. Uniform illumination of the tubes was ensured by placing it in air aluminum foil in a box with a 15W fluorescent lamp for 10 min. Control without the enzyme source was included. The absorbance was measured at 560 nm. One unit of SOD was defined as the amount of enzyme required to inhibit the reduction of NBT by 50% under the specific conditions. It was expressed as μ mg-1 protein.
Assay of Glutathione Peroxidase (GPx) Activity
Glutathione peroxidase activity was measured by the method described by Rotruck et al. (1973). The reaction mixture contained 2.0 mL of 0.4 M Tris-HCl buffer, pH 7.0, 0.01 mL of 10 mM sodium azide, 0.2 mL of enzyme. The 0.2 mL of 10 mM glutathione and 0.5 mL of 0.2 mM. H2O2. The contents were incubated at 37°C for 10 min followed by the termination of the reaction by the addition of 0.4 mL 10% (v/v) TCA, centrifuged at 5000 rpm for 5 min. The absorbance of the product was read at 430 nm and expressed as nmol mg-1 protein.
Assay of Testicular Non-enzymatic Antioxidants
Assay of Testicular Reduced Glutathione (GSH) Concentration
The GSH was determined by the method of Ellman (1959). The 1.0 mL of supernatant was treated with 0.5 mL of Ellmans reagent (19.8 mg of 5, 5’-dithiobisnitro benzoic acid (DTNB) in 100 mL of 0.1% sodium nitrate) and 3.0 mL of phosphate buffer (0.2 M, pH 8.0). The 0.4 mL of distilled water was added. The mixture was thoroughly mixed and the absorbance was read at 412 nm, expressed as nmol mg-1 protein.
Estimation of Lipid Peroxidation (Malondialdehyde)
Lipid peroxidation in the testicular tissue was estimated calorimetrically by thiobarbituric acid reactive substances TBARS method of Buege and Aust (1978). A principle component of TBARS being malondialdehyde (MDA), a product of lipid peroxidation. In brief, 0.1 mL of tissue homogenate (Tris-HCl buffer, pH 7.5) was treated with 2 mL of (1:1:1 ratio) TBA-TCA-HCl reagent (thiobarbituric acid 0.37%, 0.25 N HCl and 15% TCA) and placed in water bath for 15 min, cooled. The absorbance of clear supernatant was measured against reference blank at 535 nm. Concentration was calculated using the molar absorptivity of malondialdehyde which is 1.56x105/M/cm and expressed as nmol mg-1 protein.
Statistical Analysis
All data were expressed as Mean±SD of number of experiments (n = 10). The level of homogenecity among the groups was tested using Analysis of Variance (ANOVA) as done by Snedecor and Cochran (1980). Where heterogenecity occurred, the groups were separated using Duncan Multiple Range Test (DMRT). A value of p<0.05 was considered to indicate a significant difference between groups (Duncan, 1957).
RESULTS
Results of Phytochemical Screening of Ethanolic Extract of Grapefruit Seed
The phytochemical analysis of ethanolic extract of grapefruit seed showed the presence of the following compounds: Alkaloids, Flavonoids, Tannins, Phlobatannins and Saponins.
Body Weight Changes
Table 1 shows that rats in control and CP groups significantly (p<0.05) increased in weight by 41% of their initial mean live weight. Both DOX-administered groups lost weights when compared with their initial weights. However, the weight loss by the DOX-administered alone rats was higher than the losses by the group received CP prior to DOX treatment.
Weights and Volume of Testes Mean
The testicular weights, testis weigh b.wt.-1 ratio and volumes of the DOX-alone rats were the least, being significantly lower (p<0.001) compared to the mean testicular weights, testis weight b.wt.-1 ratio and volumes of the DOX rats that in addition had CP, which in turn were also lower but not significantly (p>0.05) lower than those of the control and CP-alone rats (Table 1).
| Table 1: | The changes in gross anatomical parameters of wistar rats |
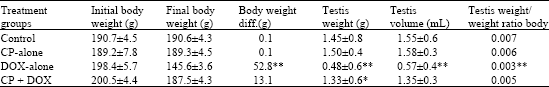 | |
| Values are Means±SEM n = 10 in each group. *,** Represent significant decreases at p<0.05 and p<0.001, respectively when compared to control values | |
| Table 2: | Testicular antioxidative enzymes and testicular contents of GSH and MDA |
 | |
| Values are Means±SEM n = 10 in each group. *, **Represent significant differences at p<0.5 and p<0.001, respectively compared to controls | |
Testicular Oxidative Stress
Activities of testicular enzymes-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPx): Administration of CP alone caused no significant (p>0.05) change in testicular SOD activity, whereas, DOX provoked a statistically significant (p<0.001) decrease in SOD activity compared to control animals. Pretreatment with CP ahead of doxorubicin challenge significantly (p<0.05) increased the testicular SOD activity compared to animals that received the anthracycline antibiotic alone. However, the SOD activity was still statistically significantly (p<0.05) lower than the control values (Table 2).
The testicular activities of CAT after CP alone were comparable to that of the control values. DOX, however, resulted in a statistically significant (p<0.001) reduction in testicular CAT activity compared to control rats. Prior administration of CP before DOX significantly increased the testicular CAT activity by more than 100% compared to doxorubicin- challenged animals. The CAT activity was approximately comparable to normal control values.
The GPx activities following CP administration approximated that of the control animals. DOX, however, markedly (p<0.001) decreased the enzyme activity compared to control values. Administration of CP before DOX significantly (p<0.05) increased the GPx activity in testicular tissue compared to animals treated with DOX alone. As shown in Table 2, CP-pretreatment before DOX did not result in significant changes in GPx activity compared with the control.
Testicular content of glutathione (GSH) and malondialdehyde (MDA): Following treatment with CP, the testicular GSH level was not significantly different from the control group. A notable reduction (p<0.05) in GSH content was, however, observed after DOX challenge when compared to the control animals. Administration of CP before DOX significantly (p<0.05) elevated the testicular content of GSH compared to animals that had the cytotoxic drug alone and returned it back to nearly its normal value (Table 2).
CP had no effect on the testicular content of lipid peroxides expressed as MDA when compared to control animals. Doxorubicin significantly elevated the testicular MDA by about 5-folds compared to the control value (Table 2).
Prior administration of CP exhibited a marked (p<0.001) reduction in the testicular MDA level compared to doxorubicin alone treated rats. However, the lipid peroxide level was still significantly higher (p<0.05) than control values (Table 2).
| Table 3: | Sperm parameters |
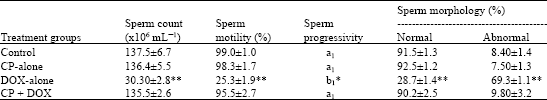 | |
| Values are Means±SEM n = 10 in each group. *,** Represent significant differences at p<0.05 and p<0.001, respectively when compared to the control values. In this study a spermatozoon was considered abnormal morphologically if it had one or more of the following features: rudimentary tail, round head and detached head. a1: rapid linear progressive motility, b1: show sluggish linear or non-linear motility | |
Sperm Parameters
Sperm Count and Sperm Motility
The CP-alone Wistar rats did not demonstrate any significant (p>0.05) difference in their sperm count and motility when compared to the control values. The animals that were given DOX without pretreatment with CP showed a significant (p<0.001) reduction in both their sperm count and motility as compared to the control groups and those groups that had a CP pretreatment (Table 3).
Sperm Progressivity and Sperm Morphology
The results of the sperm progressivity and morphology were similar to that of the sperm count and motility stated earlier. The CP-alone group similarly showed normal sperm progressivity and morphology. The animals that were given DOX without pretreatment with CP showed a significant (p<0.05 and p<0.001) reduction in both their sperm progressivity and normal sperm morphology rates, respectively as compared to the control group and those groups that had a CP pretreatment (Table 3). Again CP pretreatment abated the derangement in sperm progressivity and percentage abnormal sperm morphology.
DISCUSSION
Doxorubicin (DOX) is one of the most potent broad-spectrum antitumor anthracycline antibiotics, widely used to treat a variety of cancers (Atessahin et al., 2006). On the other hand, the clinical use of DOX is restricted because of its serious toxicity. The cytotoxic effect of Dox on malignant cells and its toxic effects on various organs (heart, liver, lung, kidney, blood cells and testis) are thought to be related to the nucleotide base intercalation and the cell membrane lipid-binding activities of DOX (Ng et al., 2006). So, the present work was designed to investigate the potential testiculoprotective effect of seed extract of CP as antioxidant-rich nutraceutical.
It is well documented that, long-term treatment by DOX causes irreversible, severe male infertility (Prahalathan et al., 2005). The mechanisms involved in such toxicity have been documented by many investigators (Yokoi and Mayi, 2004; Atessahin et al., 2006; Priestman, 2008). The involvement of oxygen free radical, superoxide radical and oxidative stress have been strongly accepted as crucial factors in the pathogenesis of DOX-induced cytotoxicity (Quiles et al., 2002; Chularojmontri et al., 2005; Prahalathan et al., 2005).
The results of the present study indicate that administration of DOX in a dose of 10 mg kg-1 b.wt. i.p. decreased the absolute testicular weights, testicular weight b.wt.-1 ratio and testicular volumes of rats. This agrees with the findings by many investigators (Kato et al., 2001; Endo et al., 2003; Atessahin et al., 2006), who provide well documented evidences of testicular morphologic impairment following DOX challenge in animal models.
Assessment of lipid peroxidation, GSH content as well as SOD, CAT, GPx and other antioxidant enzyme activities in biological tissue have been always used as markers for tissue injury and oxidative stress (Chularojmontri et al., 2005; Prahalathan et al., 2005; Atessahin et al., 2006; Priestman, 2008). In our study, testiculotoxicity and oxidative damage induced by DOX administration are also manifested by a significant increase in the activities of antioxidant enzymes, SOD, CAT, GPx and the testicular content of MDA and a significant decease of GSH. More interestingly, pretreatment of animals with seed extract of CP markedly modulates the oxidative damage induced by DOX administration.
DOX exerts its antitumor effects as well as other organ toxicity by generation of free radicals and reactive oxygen species. This increased oxidative stress damages the sperm membranes, proteins and DNA (Sikka, 1996; Kalender and Yell, 2005). This explains the reduced sperm concentration, sperm motility and sperm progressivity, with accompanying increase in abnormal sperm rates as seen in DOX alone group of rats. Pretreatment with CP containing phenolics and other potent antioxidants resulted in a remarkable normalization of these parameters.
Attempts have been made to explain the mechanisms of antioxidant effect of phenolics. These include the binding of metal ions, scavenging of reactive oxygen species (ROS); reactive nitrogen species (RNS); or their precursors, up-regulation of endogenous antioxidant enzymes, or the repair of oxidative damage to biomolecules (Ursini et al., 1999).
Clearly, present study proves the beneficial effect of the CP seed extract in protecting animals against DOX-induced testicular oxidative damage. This protecting potential of CP could be at least in part due to its free radicals scavenging capability. The fact that CP seed extract exerts profound ameliorating effects on doxorubicin-induced testicular toxicity in laboratory animals warrants a further approach for its testiculoprotective potential during DOX chemotherapy in clinical practice.
REFERENCES
- WMA and APS, 2002. Guiding principles for research involving animals and human beings. Am. J. Physiol.: Regul. Integr. Comp. Physiol., 283: R281-R283.
CrossRefPubMedDirect Link - Atessahin, A., I. Karahan, G. Turk, S. Gur, S. Yilmaz and A.O. Ceribasi, 2006. Protective role of lycopene on cisplatin-induced changes in sperm characteristics, testicular damage and oxidative stress in rats. Reprod. Toxicol., 21: 42-47.
CrossRefDirect Link - Banerjee, H.N. and M. Verma, 2008. Application of nanotechnology in cancer. Technol Cancer Res. Treat., 7: 149-154.
Direct Link - Berthiaume, J.M. and K.B. Wallace, 2007. Adriamycin-induced oxidative mitochondrial cardiotoxicity. Cell Biol. Toxicol., 23: 15-25.
CrossRefDirect Link - Chularojmontri, L., S.K. Wattanapitayakul, A. Herunsalee, S. Charuchongkolwongse, N. Niumsakul and S. Srichairat, 2005. Antioxidative and cardioprotective effects of Phyllanthus urinaria L. on doxorubicin-induced cardiotoxicity. Biol. Pharm. Bull., 28: 1165-1171.
CrossRefPubMedDirect Link - Davies, K.J. and J.H. Doroshow, 1986. Redox cycling of anthracyclines by cardiac mitochondria. I. Anthracycline radical formation by NADH dehydrogenase. J. Biol. Chem., 261: 3060-3067.
PubMedDirect Link - Duncan, D.B., 1957. Multiple range tests for correlated and heteroscedastic means. Biometrics, 13: 164-176.
CrossRefDirect Link - Ellman, G.L., 1959. Tissue sulfhydryl groups. Arch. Biochem. Biophys., 82: 70-77.
CrossRefPubMedDirect Link - Endo, F., F. Manabe, H. Takeshima and H. Akaza, 2003. Protecting spermatogonia from apoptosis induced by doxorubicine using the luteinizing hormone-releasing hormone analog leuprorelin. Int. J. Urol., 10: 72-77.
Direct Link - Frederick, C.A., L.D. Williams, G. Ughetto, G.A. van der Marel, J.H. van Boom, A. Rich and A.H. Wang, 1990. Structural comparison of anticancer drug-DNA complexes: Adriamycin and daunomycin. Biochemistry, 29: 2538-2549.
Direct Link - Halliwell, B., 2008. Are polyphenols antioxidants or pro-oxidants? What do we learn from cell culture and in vivo studies? Arch. Biochem. Biophys., 476: 107-112.
CrossRefPubMedDirect Link - Heggers, J.P., J. Cottingham, J. Gusman, L. Reagor and L. McCoy et al., 2002. The effectiveness of processed grapefruit-seed extract as an antibacterial agent: II. Mechanism of action and in vitro toxicity. J. Altern Compl. Med., 8: 333-340.
CrossRefPubMedDirect Link - Heller, C.G. and Y. Clermont, 1964. Spermatogenesis in man: An estimate of its duration. Science, 140: 184-186.
Direct Link - Kalender, Y., M. Yel and S. Kalender, 2005. Doxorubicin hepatotoxicity and hepatic free radical metabolism in rats: The effects of vitamin E and catechin. Toxicology, 209: 39-45.
CrossRefDirect Link - Kato, M., S. Makino, H. Kimura and T. Ota, 2001. Sperm motion analysis in rats treated with adriamycin and its applicability to male reproductive toxicity studies. J. Toxicol. Sci., 26: 51-59.
PubMedDirect Link - Mornparler, R.L., M. Karon, S.E. Siegel and F. Avila, 1976. Effect of Adriamycin on DNA, RNA and Protein synthesis in cell free systems and intact cells. Cancer Res., 36: 2891-2895.
Direct Link - Ng, R., N. Better and M.D. Green, 2006. Anti-cancer agents and cardiotoxicity. Semin. Oncol., 33: 2-14.
Direct Link - Pacher, P., J.S. Beckman and L. Liaudet, 2007. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev., 87: 315-424.
CrossRefPubMedDirect Link - Pigram, W.J., W. Fuller and L.D. Hamilton, 1972. Stereochemistry of intercalation: Interaction of daunomycin with DNA. Nat. New Biol., 235: 17-19.
PubMed - Prahalathan, C., E. Selvekuma and P. Varalakshmi, 2005. Lipoic acid ameliorates adriamycin-induced testicular mitochondriopathy. Reprod. Toxicol., 20: 111-116.
CrossRef - Priestman, T., 2008. Cancer Chemotherapy in Clinical Practice. Springer-Verlag, London, UK., SBN: 978-1-84628-989-7, pp:130-136.
Direct Link - Quiles, J.L., J.R. Huertas, M. Battino, J. Mataix and C.M. Ramirez-Tortosa, 2002. Antioxidant nutrients and adriamycin toxicity. Toxicology, 180: 79-95.
CrossRefPubMedDirect Link - Rotruck, J.T., A.L. Pope, H.E. Ganther, A.B. Swanson, D.G. Hafeman and W.G. Hoekstra, 1973. Selenium: Biochemical role as a component of glutathione peroxidase. Science, 179: 588-590.
CrossRefPubMedDirect Link - Rukmini, M.S., B. D'Souza and V. D'Souza, 2004. Superoxide dismutase and catalase activities and their correlation with malondialdehyde in schizophrenic patients. Indian J. Clin. Biochem., 19: 114-118.
CrossRefDirect Link - Saalu, L.C., L.A. Enye and A.A. Osinubi, 2009. An assessment of the histomorphometric evidences of doxorubicin-induced testicular cytotoxicity in wistar rats. Int. J. Med. Med. Sci., 1: 370-374.
Direct Link - Saalu, L.C., G.O. Ajayi, A.A. Adeneye, I.O. Imosemi and A.A. Osinubi, 2009. Ethanolic seed extract of grapefruit (Citrus paradisi Macfad) as an effective attenuator of doxorubicin-induced oxidative stress in the rat heart. Int. J. Cancer Res., 5: 44-52.
CrossRefDirect Link - Sikka, S.C., 1996. Oxidating stress and role of antioxidants in normal and abnormal sperm function. Front Bisobene, 1: 78-86.
PubMed - Singh, K.K., 2005. Nanotechnology in cancer detection and treatment. Technol. Cancer Res. Treat. 4: 583-584.
Direct Link - Sonmez, M., G. Turk and A. Yuce, 2005. The effect of ascorbic acid supplementation on sperm quality, lipid peroxidation and testosterone levels of male Wistar rats. Theriogenology, 63: 2063-2072.
CrossRefDirect Link - Ursini, F., F. Tubaro, J. Rong and A. Sevanian, 1999. Optimization of nutrition: Polyphenols and vascualr protection. Nutr. Rev., 57: 241-249.
PubMed - Winterbourn, C.C., R.E. Hawkins, M. Brian and R.W. Carrell, 1975. The estimation of red cell superoxide dismutase activity. J. Lab. Clin. Med., 85: 337-341.
PubMedDirect Link - Wolinsky, J.B. and M.W. Grinstaff, 2008. Therapeutic and diagnostic applications of dendrimers for cancer treatment. Adv. Drug Deliv. Rev., 60: 1037-1055.
PubMed - Yeh, Y.C., L. Hui-Chi and T. Chih-Tai, 2007. The effects of Doxocycline on Doxorubcin induced oxidative damage and apoptosis in mouse testis. Biol. Reprod., 77: 217-217.
Direct Link
emmanuel ogundimu. Reply
the authors of this journal are successful in making their research. therefore, this work is practicable.