
V.A. Maina
Department of Veterinary Surgery and Reproduction, Faculty of Veterinary Medicine, University of Maiduguri, P.M.B. 1069, Maiduguri, Borno State, Nigeria
A. Muktar
Department of Veterinary Surgery and Reproduction, Faculty of Veterinary Medicine, University of Maiduguri, P.M.B. 1069, Maiduguri, Borno State, Nigeria
Y.G. Sabo
Department of Veterinary Services, Ministry of Livestock and Nomadic Settlement, P.M.B. 2030, Yola, Adamawa State, Nigeria
Asian Journal of Scientific Research
Year: 2008 | Volume: 1 | Issue: 4 | Page No.: 421-428







ABSTRACT
The ovaries of 127 Bos indicus cows were used to evaluate the influence of Body Condition Score (BCS) on ovarian activity. Each cow was given a body condition score using a scale of 1 to 5 before slaughter. The presence or absence of pregnancy was noted after slaughter and the ovaries were collected. Follicles were measured and identified as small (1 to 4 mm), medium (5 to 8 mm) or large (≥9 mm). Corpus luteum was classified as functional CL or Corpus albicans based on gross morphology. Luteal structures and follicles were compared with different phases of the oestrus cycle. Based on this, the cows were classified as cycling, pregnant, anestrus, prepubertal heifers or having abnormal ovarian cyclicity. Cows with BCS 3 exhibited more medium follicles (p<0.001) than cows with BCS 1 and BCS 2. Cows with BCS 3 had more large follicles (p<0.01) than cows with BCS 1. The incidences of cycling and pregnant animals correlated positively with BCS. Incidences of anestrus and abnormal ovarian cyclicity correlated negatively with BCS. It was concluded that change in BCS had significant effect on ovarian function of Zebus cows.
PDF Abstract XML References Citation
How to cite this article
V.A. Maina, A. Muktar and Y.G. Sabo, 2008. Effects of Body Condition Score on Ovarian Activity of Bos indicus (ZEBU) Cows. Asian Journal of Scientific Research, 1: 421-428.
DOI: 10.3923/ajsr.2008.421.428
URL: https://scialert.net/abstract/?doi=ajsr.2008.421.428
DOI: 10.3923/ajsr.2008.421.428
URL: https://scialert.net/abstract/?doi=ajsr.2008.421.428
INTRODUCTION
Cattle belong to the genus Bos and the major classes of domestic cattle in the World are Bos taurus and Bos indicus and their crosses (Bos taurus x Box indicus) (McDonald and Low, 1985). Bos indicus are the humped zebu cattle, which are indigenous to the tropical and subtropical regions of Africa and Asia. They are adapted to the tropical climate and are able to withstand greater heat stress than the temperate breeds of cattle. They are slower in attainment of puberty and generally have poor reproductive performance and productivity (Payne, 1990; Maule, 1990). The reasons for the documented poor productivity may not be unconnected with the fact that the Bos indicus breeds are usually subjected to bridge up poor and harsh extensive management practices unlike the Bos taurus.
Nigeria has an estimated cattle population of 13.9 million (Bourne et al., 1994). More than ninety percent of this cattle population is in the hands of small scale traditional livestock rearers and nomads who depend entirely on grazing natural pasture and to some extent crop residues during the dry season with little or no supplementation (Voh and Otchere, 1989).
In the tropics, cattle are valued for milk, meat production and hides used in leatherwork, dung for fuel and fertilizer, blood drank by certain tribes. Cattle horns and bones are also highly valued. Cattle are also used extensively for drought purposes in many parts of Africa, where they are involved in a variety of farm work, transportation and lifting water (Payne, 1990; Maule, 1990).
In many cattle owning tribes, cattle are regarded as a means of wealth and prestige. They serve as a means of fulfillment and a mark of respectability and status in the society (Payne, 1990). They are also used in the payment of bride price by the bridegroom, which is an ancient and well established practice in nomadic tribes (Maule, 1990).
Cattle are non seasonal breeding animals; hence reproductive activity takes place throughout the year with cows exhibiting recurrent and regular oestrus cycles once it reaches maturity. This periodicity can however be suppressed as a result of poor nutrition and perhaps by high milk yield and calf sucking (Liang et al., 1988).
The cyclic activity of the cow`s reproductive system can be assessed from ovarian function using follicular population and sizes and the presence of corpora lutea (Dominguez, 1995).
Nutritional factors have been studied concerning their effects on reproduction in cattle (Sabo et al., 2008). It has been well established that balanced nutrition is essential for efficient reproductive functions in both sexes in cattle. In tropical countries like Nigeria where over ninety percent of cattle are managed under the extensive system, poor fertility is not an uncommon problem among the cattle breeds. In addition to nutrition, harsh climatic conditions and poor management are factors that contribute to infertility problems in the tropics (Rekwot et al., 1987; Sekoni et al., 1988; Sabo et al., 2008).
Body condition score is a technique that can be used to estimate the quantity of fat deposition in certain areas of an animal`s body (Braun et al., 1986; Nicholson and Butherworth, 1986; Msagi et al., 2005) and it offers a practical means to assess the nutritional status of animals (Dominguez, 1995).
Reports on the effects of body condition on reproduction in cattle have been studied by various researchers (Nicholson and Butherworth, 1986; Pathiraja et al., 1988).
Despite many findings and reports on the effect of nutritional factors on fertility and reproductive parameters in cows, information on the role of these factors within the ovaries is scarce (Dominguez, 1995).
The objective of this study is to determine whether body condition score has influence on follicular population and on proportion of small, medium and large follicle categories in the ovaries of zebu cows.
MATERIALS AND METHODS
Study Area and Period of Study
This study was conducted on cows slaughtered at Maiduguri Abattoir, Borno State, Nigeria, during the period between the months of April to August, 2005 (peri-rainy to rainy season). The cows were indigenous to the geographical areas which cover the Sudans Savannah and Sahels Savannah (arid to semi-arid regions of North Eastern Nigeria and neighboring parts of Niger, Chad and Cameroon Republics). The area are characterized by low annual rainfall and sparse vegetation. The cattle are generally kept in an extensive management system.
Animals and Sample Collection
This study was conducted on a total of 127 cows slaughtered at the Maiduguri Abattoir. Each cow was examined before slaughter (antemortem) and body condition scoring was conducted according to the method described by Braun et al. (1986) using a scale of 1 to 5. After slaughter (post mortem), the reproductive organs were brought out and the presence or absence of pregnancy was noted, but examination for trimester of pregnancy was not made. The ovaries were then collected and tagged with labels indicating the serial number of the animals. The samples were then transported immediately to the laboratory for examination.
| Table 1: | Monthly mean maximum temperatures and rainfall data for Maiduguri for 2005 |
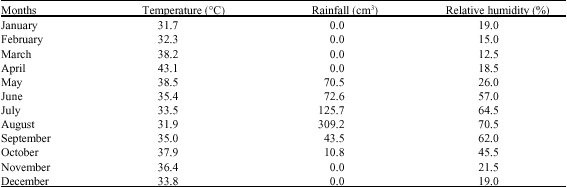 | |
Sample Processing and Examination
The ovaries were trimmed of excessive connective tissue and washed with tap water. The total number of visible follicles on the surface of each ovary was counted and the diameter of each follicle was measured using venier caliper. Follicles were classified into 3 size categories. Small follicles (1 to 4 mm), medium follicles (5 to 8 mm) and large follicles (>=9 mm) (Dominguez, 1995; Arthur et al., 1998).
Luteal structures were identified on the ovaries and each structure was classified as active Corpus luteum, Corpus albicans or Corpus luteum of pregnancy based on gross morphology as described by Arthur et al. (1998) and Hafez (1993). Ovaries showing the presence of active Corpus luteum were considered to belong to cycling cows. Corpus luteum of pregnancy was identified during sample collection by the presence of a foetus in the uterus. Cows that had luteal structure(s) on its ovaries with complete absence of follicles were defined as anestrus. A cow that had no Corpus luteum in any of its ovaries was defined as prepubertal heifer irrespective of the presence of follicles and number and size of the follicles.
Statistical Analysis and Presentation of Results
One way analysis of variance was used to evaluate the effects of body condition score on the populations of small follicles, medium follicles and large follicles and the level of significance determined for different follicular populations (p<0.01). Data on cycling cows, pregnancy, anestrus, prepubertal heifers and those with abnormal ovarian activity was used to evaluate the incidence of each with respect to body condition score. These incidences were subjected to correlation analysis to find the value of correlation coefficient (r) for each incidence in relation to body condition score.
The mean temperature and rainfall for the year 2005 from January to December are presented in Table 1.
RESULTS
Out of the 127 cows that were studied, the maximum body condition score (BCS) recorded was 3 on a scale of 1 to 5. There were 33 (25.98%) cows in BCS1, (44.88%) cows in BCS 2 and 37 (29.13%) cows in BCS 3 group.
Follicular Populations
Out of the 127 cows, 112 (88. 19%) show visible follicular growth on either of their ovaries, while 15 (11.81%) did not show visible follicles on any of their ovaries. A total of 783 follicles were identified and measured, out of which 607 (77.52%) were small follicles, 162(20.69%) were medium follicles and 14(1.79) were large follicles.
| Table 2: | Effect of body condition score or follicular population |
 | |
| Within each column, values with different superscripts differ significantly (a, b: p<0.001; c, d: p<0.001) | |
| Table 3: | Incidences of various functional states of ovaries with respect to body condition scores |
 | |
Body condition score did not show effect on number of small follicles, but exerted a significant effect on the population of medium and large follicles (Table 2). Thus cows with BCS 3 exhibited significantly more medium follicles than cows with BCSI and BCS2 (p>0.001), similarly cows with body condition score 3 exhibited significantly more large follicles than cows with body condition score 1 (p<0.01).
However, cows with BSC 1 and 2 as well as BSC 2 and 3 did not significantly differ (p<0.005) between them with respect to population of larger follicles.
Luteal Structures, Follicular Development and Ovarian Activity
Of the 127 cows that were evaluated 20(15.75%) showed active Corpus luteum on either of the ovaries indicating a state of cyclicity, 15(11.81%) were pregnant, 70(55.18%) exhibited ovarian structures that suggested abnormal activity, 15(11.81%) were anestus, while 9(7.09%) exhibited ovarian structures of prepubertal heifers.
The incidences of all the state of activity showed correlation with body condition score (Table 3). Incidences of cycling cows, pregnant cows and prepubertal heifers were correlated positively to body condition score with the correlation coefficient (r) of 0.89, 0.93 and 0.81, respectively. Conversely, the incidences of anestrous and abnormal ovarian activity were negatively correlated to the body condition score with the correlation coefficient (r) of 0.93 and 0.67, respectively.
DISCUSSION
From the result obtained in this study, the average number of small follicles was significantly higher (p<0.001) than medium and large follicles. This can be explained by the fact that bovine ovaries possess a large amount of small follicular reserve from which graafian follicles are continuously recruited for further development (Sabo et al., 2008).
It has been documented by Fortune (1993) that follicular developments in cows occurs in a wave pattern throughout the length of the oestrus cycle, in which three to six cohorts of follicles of 5 mm or more in diameter are recruited from a pool of smaller follicles to initiate each wave. One dominant follicle becomes larger than the others and proceeds to ovulation or regression while the subordinate follicles undergo regression. This phenomenon may account for fewer medium and large follicles than small follicles recorded in this study.
The estrus cycle is hormonally controlled and for the purpose of understanding may be divided into four phases, estrus, metestrus, diestrus and pro-estrus. The pro-estrus and estrus (follicular phase) period are primary under the influence of estrogen while the diestrus and metestrus which are associated with growth of the Corpus luteum and are primarily under the influence of progesterone (Parker and Mathins, 2002; Chaudhari and Sabo, 2006).
The ovaries that showed a functional Corpus luteum were in state of normal function and that the cows were in a diestrus state (Sabo et al., 2008). The presence of follicles ranging from small to large sizes in these ovaries was a further indication that the wave pattern of follicular turnover was taking place, therefore a state of normal cyclicity.
The ovaries that were considered as having abnormal cyclicity although exhibited follicles at various stages of growth could not be classified into any of the phases of the oestrus cycle, namely metestrus, diestrus, proestrus or estrus based on the luteal structures present on them.
The presence of Corpora albicantia at various stages of degeneration is evidence that the cows had, previous oestrus cycles, because corpora albicanot persist and remain visible as fibrous connective tissue scars on the ovaries for several additional oestrus cycles (Mc Donald, 1980).
Metestrus is the stage of early Corpus luteum formation characterized by the presence of corpus heamorrhagicum which appears as a blood clot filled follicular cavity, while the diestrus state is characterized by a prominent functional Corpus luteum (Arthur et al., 1998). This implies these ovaries were in either of the luteal phases of the estrus cycle.
Also proestrus and oestrus are referred to as the follicular phase of the estrus cycle in which there is a prominent pre-ovulatory follicle together with a regressing Corpus luteum. The ovaries classified as being in an abnormal state exhibited entirely small and medium follicles (Chaudhari and Sabo, 2006).
The occurrence of abnormal ovarian function has been reported by many workers in cows. Dawuda et al. (1988) described various pattern associated with ovarian activity in postpartum Nigerian cows. Such patterns included short, prolonged or irregular cycles ending in many weeks of luteal inactivity prior to the initiation of the next cycle.
Short estrus cycle is also a common abnormality in postpartum beef cows where the Corpus luteum formed is smaller in size, sub functional and undergoes pre-mature regression, which renders the cycle shorter than normal (Short et al., 1990).
Going by these facts, it could be postulated that the ovaries classified as being in a state of abnormal cyclicity could have been from cows that were in a postpartum state. Experiencing similar abnormalities and irregularities of cycles like the ones described above, since Bos indicus cows have a postpartum period of one year or more considering the average calving interval of 2 years as reported by Voh and Otchere (1989) in Nigerian herds.
In addition some of the cows might be old aged cows that were culled from herds due to decline in reproductive performance (Voh and Otchere, 1989).
Comparing the ovarian structures and the type of abnormal functions mentioned earlier, it can be said that the cows with ovaries that had corpora albicantia and small or medium follicles might be experiencing prolonged, irregular estrus cycles, (Dawuda et al., 1988) in which the Corpus luteum of diestrus regressed, but the follicles failed to proceed development to pre ovulatory stage, perhaps due to suppression of pulsatile secretion of LH as it occurs in early post partum cows (Short et al., 1990) or in severely malnourished cows (Richards et al., 1989).
In other situations, the cows might be in the process of re-initiating their normal cyclicity after recovering from suckling stress or under-nutrition. In this case, the dormant follicles might be in the process of resumption of development to mature stage. Also animals experiencing irregular estrus cycles may still maintain the wave of follicular development and regression if the basal level of gonado-tropins is maintained. Thus large follicles could be present in ovaries that are in a state of prolonged luteal inactivity without ovulating, if the pre-ovulatory LH surge fails to occur.
The complete absence of follicular structures observed in the ovaries of some cows was defined as a state of anestrus due to complete ovarian inactivity, because the presence of follicles of various sizes is one of the indications of an active ovary (Hopper et al., 1993). Furthermore remnants of Luteal structures observed on the ovaries was an affirmation that the cows had experienced successful ovulations and Corpus luteum formation prior to complete atresia or disappearance of the follicles rendered the cows to be in an anestrus state.
Other ovaries were classified as prepubertal because no luteal structures could be detected on them, despite the presence of follicles. The absence of luteal structures means that the cows had never ovulated as documented by Hopper et al. (1993).
The influence of body condition score on follicular development manifested only at the level of medium and large follicle populations in this study. This differs from the report of Dominguez (1995) which revealed that a significant effect of body condition score on small and large follicles but not medium follicles.
Despite this difference, however the significant increase in number of large follicles with increase in body condition score indicated the changes in body nutrient reserve have an effect on follicular development and maturation in cattle.
Low nutrition can prevent final growth of follicles due to a negative hypothalamus feed back effect of oestradiol on the GnRH. The resultant decrease in LH pulses makes follicles not to grow sufficiently to reach to pre-ovulatory size, so that sufficient oestradiol can be produced to elicit an LH surge and ovulation (Milo, 1998; Sabo et al., 2008).
Nutritional stress has been found to lead to anestrus behaviour while the demands of lactation can lead to lactational anestrus (Perera, 1999), culminating in low fertility in cows.
Richards et al. (1989) induced anestrus in sexually mature beef cows by feeding them restricted dietary energy to enable loss of weight and body condition score and he demonstrated a decrease in the frequency of LH pulses associated with anestrus.
Schillo (1992) documented that increase in the frequency of pulsatile secretion of LH is necessary for growth and maturation of ovarian follicles to the pre-ovulatory stage and thus for induction of oestrus and ovulation.
Milo (1998) stated the number of follicular waves during an oestrus cycle can be altered by nutrition which causes an increased percentage of heifers with three follicular waves than when lower amount of energy are fed.
It could therefore be stated that the apparent suppression of medium and large follicular growth observed in seriously emaciated cows (BCS1) compared to extremely high number of large follicles in BCS 3 group is due to difference in LH secretion between the 2 groups.
In addition to follicular growth, the incidences of other functional states of the ovaries also exhibited a clear influence of body condition score.
These were positive correlation between body condition score and incidences of cycling and pregnant cows. The incidence of prepubertal heifers also correlated positively with Body Condition Score (BCS).
Schillo (1992) reported that changes in body fatness is associated with changes in reproductive activity in cattle. There is also restoration of normal ovarian activity and conception, when the state of under-nutrition or negative energy balance is reversed (Imakawa et al., 1986; Sabo et al., 2008).
The negative correlation between body condition score and the incidence of anestrus cows on one hand and that of cows with abnormal ovarian cyclicity on the other agrees with the report of (Imakawa et al., 1986) that cows that are losing body weight or are thin do not have estrus cycles or have irregular cycles.
Apart from cessation of estrus cycle in seriously malnourished and emaciated cows, there is restoration of normal ovarian activity and conception when the sate of under-nutrition or negative energy balance is reversed (Johnson et al., 1987; Richards et al., 1989).
In conclusion, this study indicated that body condition score a measure of body fat and nutrient reserve has a significant effect on ovarian function of Bos indicus (zebu) cows through increased follicular development, ovulation and corpus luteum formation, normal cyclicity and pregnancy rate in cows with good body condition and increased proportion of anestrus and abnormal ovarian function in severely emaciated cows having poor body condition.
REFERENCES
- Braun, R.K., G.A. Donovan, T.Q. Tran, J.K. Shearer and E.L. Bliss et al, 1986. Body condition scoring, dairy cows as a herd management tool. Comp. Contin. Edu. Pract. Vet., 10: 62-67.
Direct Link - Chaudhari, S.U.R. and Y.G. Sabo, 2006. Techniques for reproductive efficiency with reference to oestrus detection and timing of insemination in cattle. J. Applied Sci., 6: 2141-2150.
CrossRefDirect Link - Dawuda, P.M., E.O. Oyedipe, N. Pathiraja and A.A. Voh, 1988. Serum progesterone concentration during the postpartum period of indigenous Nigerian zebu cows. Br. Vet. J., 114: 213-257.
PubMed - Dominguez, M.M., 1995. Effects of body condition, reproductive status and breed on follicular population and oocyte quality in cows. Theriogenology, 43: 1405-1418.
CrossRefDirect Link - Fortune, J.E., 1993. Follicular dynamics during the bovine estrous cycle: A limiting factor in improvement of fertility? Anim. Reprod. Sci., 33: 111-125.
CrossRefDirect Link - Hopper, W.W., R.W. Silox, D.L. Byerley and T.E. Kiser, 1993. Follicular development in prepubertal heifers. Anim. Reprod. Sci., 31: 7-12.
Direct Link - Imakawa, K., M.L. Day, M. Garcia, D.D. Winder and R.J. Zalesky et al., 1986. Endocrine changes during restoration of estrus cycles following induction of anestrus by restricted nutrient intake in beef, heifers. J. Anim. Sci., 63: 565-571.
Direct Link - Johnson, M.S., T.N. Wegner and D.E. Ray, 1987. Effects of elevating serum lipids on luteinizing hormone response to gonadotropin releasing hormone challenge in dietary deficient anestrus heifers. Theriogenology, 27: 421-429.
CrossRef - Milo, C.W., 1998. Information on regulation of reproductive cyclicity in cattle. Bovine Proc., 31: 26-33.
Direct Link - Parker, R. and C. Mathins, 2002. Reproductive tract anatomy and physiology of the cow. Guide B., 212: 1-4.
Direct Link - Pathiraja, N., E.O. Oyedipe, P.M. Dawuda and S.U.R. Chaudhari, 1988. Intensive breeding schemes in Zebu cattle. World Anim. Rev. (FAO), 65: 55-57.
Direct Link - Rekwot, P.I., A.A. Voh, E.O. Oyedipe, G.I. Opaluwa, V.O. Sekoni and P.M. Dawuda, 1987. Influence of season on characteristics of the ejaculates from bulls in an artificial insemination centre in Nigeria. Anim. Reprod. Sci., 14: 187-194.
Direct Link - Richards, M.W., R.P. Wettemann and H.M. Schoenemann, 1989. Nutritional anestrus in beef cows: Body weight change, body condition, luteinizing hormone in serum and ovarian activity. J. Anim. Sci., 67: 1520-1526.
CrossRefDirect Link - Sabo, Y.G., U.K. Sandabe, V.A. Maina and H.G. Balla, 2008. Schemes for oestrus synchronization protocols and controlled breeding programs in cattle. J. Applied Sci., 8: 241-251.
CrossRefDirect Link - Schillo, K.K., 1992. Effects of dietary energy on control of luteinizing hormone secretion in cattle and sheep. J. Anim. Sci., 70: 1271-1282.
Direct Link - Sekoni, V.O., J. Kumi-Diaka, D.I. Saror, C.O. Njoku and S.A.S. Olurunju, 1988. Seasonal and monthly variations in the incidence of morphological abnormalities in bovine spermatozoa in Shika, Zaria, Northern Nigeria. Anim. Reprod. Sci., 17: 61-67.
Direct Link - Short, R.E., R.A. Bellows, R.B. Staigmiller, J.G. Berardinelli and E.E. Custer, 1990. Physiological mechanisms controlling anestrus and infertility in postpartum beef cattle. J. Anim. Sci., 68: 799-816.
CrossRefDirect Link - Voh, A.A. and E.O. Otchere, 1989. Reproductive performance of zebu cattle under traditional agropastoral management in Northern Nigeria. Anim. Reprod. Sci., 19: 191-203.
Direct Link