
M.H.H. Awaad
Departement of Poultry Diseases, Faculty of Veterinary Medicine, Cairo University, Egypt
Wafaa A. Abd El-Ghany
Departement of Poultry Diseases, Faculty of Veterinary Medicine, Cairo University, Egypt
LiveDNA: 20.3771
S.A. Nasef
Animal Health Research Institute, Dokki, Cairo, Egypt
Medhat S. El-Halawany
Departement of Biology, Faculty of Science, Cairo University, Egypt
Faten F. Mohamed
Departement of Pathology, Faculty of Veterinary Medicine, Cairo University, Egypt
Ahmed F. Gaber
Animal Health Research Institute, Dokki, Cairo, Egypt
Asian Journal of Poultry Science
Year: 2016 | Volume: 10 | Issue: 3 | Page No.: 126-133







ABSTRACT
Objectives: The efficacy of microencapsulated Na-butyrate (NaB) supplementation for reducing Salmonella enteritidis (S. enteritidis) infection was investigated. Methodology: One day old broiler chicks (n = 200) were allotted into 4 equal groups. At 2nd day of age, birds of groups 3 and 4 were challenged by crop gavage with 5x108 CFU mL–1 of S. enteritidis. Groups 2 and 3 were supplemented with NaB diet while 1 and 4 were fed on a plain ration without treatment. Scanning Electron Microscopy (SEM) of re-isolated S. enteritidis of challenged NaB treated broilers revealed broken cells with morphological changes in comparison with untreated challenged group, however, Transmission Electron Microscopy (TEM) showed several lipids like bodies that barely detected in untreated group. Results: Results from Polymerase Chain Reaction (PCR) analyses revealed that S. enteritidis strain from untreated challenged control harbored virulence genes invA, stn and pefA with incidence rate of 100%. While, NaB supplementation resulted in detection of 100% of invA gene and only 66.6% of pefA and stn genes. Gut integrity of NaB supplemented groups showed significant (p≤0.05) increase in villi height and villus height: crypt depth ratio and had positive effect in maintenance of healthy intestinal epithelial cells. Conclusion: NaB supplement is effective in reducing S. enteritidis in chickens.
PDF Abstract XML References Citation
Received: February 24, 2016;
Accepted: March 28, 2016;
Published: June 15, 2016
How to cite this article
M.H.H. Awaad, Wafaa A. Abd El-Ghany, S.A. Nasef, Medhat S. El-Halawany, Faten F. Mohamed and Ahmed F. Gaber, 2016. Effect of Na-butyrate Supplementation on Electromicroscopy, Virulence Gene Expression Analysis and Gut Integrity of Experimentally Induced Salmonella enteritidis in Broiler Chickens. Asian Journal of Poultry Science, 10: 126-133.
DOI: 10.3923/ajpsaj.2016.126.133
URL: https://scialert.net/abstract/?doi=ajpsaj.2016.126.133
DOI: 10.3923/ajpsaj.2016.126.133
URL: https://scialert.net/abstract/?doi=ajpsaj.2016.126.133
INTRODUCTION
The importance of gastrointestinal tract health in broiler chickens has been increasingly recognized due to its contribution to their overall health and performance1. Salmonellae infections can cause significant growth depression or even mortality in young birds. Salmonellosis is a zoonotic bacterial disease of significant importance2. Salmonella enteritidis (S. enteritidis) is one of the main Salmonella serovars involved in human food poisoning outbreaks and sometimes deaths3. Organic acids are among the alternative antibiotic growth promoters that have already being used in practice for decades to stimulate growth performance in poultry4-6. Amongst these organic acids, Short Chain Fatty Acids (SCFA) are considered as potential alternatives7. Butyric acid is one of these SCFA which has higher bactericidal activity when the acid is undissociated8,9. Accordingly, it can be used for the treatment of several intestinal bacterial infections like salmonellosis10.
The present trial was conducted to elucidate the effects of usage of sodium butyrate (NaB) encapsulated in palm fat on intestinal enumeration, electomicroscopic morphology, virulence genes and intestinal histomorphometry of experimentally induced S. enteritidis infection in broilers.
MATERIALS AND METHODS
Na-butyrate (NaB): That product of Na-butyrate encapsulated in palm fat(Admix®30) (produced by NUTRI-AD International, Belgium) was used in this trial in the following dietary levels in the test group; starter diet 1 kg t–1, grower diet 0.5 kg t–1 and finisher diet 0.25 kg t–1.
Experimental birds: One day-old cobb broiler chickens (n = 205) were used in this study. Chick box paper liners as well as organs of 5 sacrificed chicks were cultured to screen for presence of Salmonellae. The remaining birds were allotted into 4 equal groups (1-4). Each group divided into 5 replicates consisting of 10 birds each. The chickens were vaccinated against different diseases according to the vaccination programs usually adopted in Egyptian chicken broiler farms. The commercial diets used were formulated to meet the nutrient requirements of the broiler chicks during starter, grower and finisher periods according to the National Research Council11. Feed and water were given ad libitum.
Experimental design: The experiment was carried out according to the National Regulations on Animal Welfare and approved by Institutional Animal Ethical Committee, Faculty of Veterinary Medicine, Cairo University. Chickens of groups 2 and 3 were supplemented with NaB diet, while groups 1 and 4 fed on a plain ration without treatment. At 2nd day of age, birds of groups 3 and 4 individually infected by crop gavage with 5×108 CFU mL–1 of S. enteritidis12. The challenge inoculate checked for purity and confirmed by PCR. Chickens of group 1 were kept without infection or NaB supplementation as blank controls. At 7th and 19th day of age, 5 birds/group were sacrificed and subjected to bacteriological examination for S. enteritidis re-isolation from caecum and liver. At the end of the experiment (35 days), 5 caecal contents from each group were examined for S. enteritidis enumeration (colonization)13.
Electron microscopy examination: At 17th day of age, caecal specimens of 3 birds of groups 3 and 4 were similarly subjected to S. enteritidis re-isolation and the isolated strains were subjected to electron microscopy examination for detection of any morphological alteration. In SEM, S. enteritidis randomly selected colonies (isolated from group 3 and 4) cultured on agar medium were first excised and trimmed, then fixed by 2.5% glutaraldehyde. The samples were dehydrated by serial dilution of ethanol and then dried using Audissamdri-815 CO2 critical point drier (Tousimis Inc., Rockville, MD, USA). The samples were coated by gold sputter coater (SPI-module) and examined using JOEL-JSM 5500 (JEOL Ltd., Tokyo, Japan) scanning electron microscope was adopted at 20 kV at the Regional Center of Mycology and Biotechnology, Al-Azhar University, Cairo, Egypt. The TEM was performed using standard procedures. Briefly, grown S. enteritidis cells (isolated from 3 and 4) at mid-log phase were harvested by centrifugation (4000×g for 10 min) and fixed in 2.5% glutaraldehyde in PBS (pH 7.4). This was followed by fixation in 1% osmium tetroxide and then dehydrated by serial dehydration in ascending ethanol rinses. Thin sections of epoxy resin-embedded specimens were imaged on a JEM-1400 (JEOL Ltd., Tokyo, Japan) operated at accelerating voltage of 80 kV at the electron microscopy unit, Cairo University Research Park (CURP), Giza, Egypt.
Detection of S. enteritidis virulence gene expression using conventional PCR: The DNA of the isolated bacterial strains was extracted and amplified for virulence genes (invA, pefA and stn). Salmonella enteritidis DNA was extracted using commercially available kit, QIAamp DNA Mini Kit, (Cat. # 51304) according to the manufacturer’s instructions. Virulence genes were amplified using their specific primers14,15. The amplification products were photographed by a gel documentation system and the data was analyzed through computer software.
Gut integrity assay (histomorphometric assay): At 35 days of age, 3 birds from groups 1-4 were sacrificed and small intestine were collected and immediately immersed in 10% buffered formalin. After fixation, 1 cm-thick samples were taken from the jejunum and ileum. Routine histological laboratory methods including dehydration, clearing and paraffin embedding were used and paraffin blocks were made. The slides stained by haematoxylin and eosin16. Histological indices were measured using digital photography and light microscopy. The villus height was measured from the apical to the basal region, which corresponds to the superior portion of the crypts. Crypts were measured from the basis until the region of transition between the crypt and the villus.
Statistical analysis: One-way analysis of variance adopted using SAS software general liner models procedure17. The main factor was NaB supplementation as a mean effect. Mean values assessed for significance using Duncan’s multiple range tests18.
RESULTS AND DISCUSSION
In the current study, re-isolation of S. enteritidis from liver and caecum of experimentally infected chickens at day 19th of age revealed 0% in NaB supplemented group as compared with 80% in infected untreated broilers. Reduction in S. enteritidis enumeration in the caecum of NaB supplemented group at 35th day of age (a mean of 101 and 1.1×104 CFU g–1 in NaB supplemented and positive control groups, respectively) was important for the microbiological safety of poultry products, as this site and cloaca represent 2 common locations in the birds where the bacteria are present in high numbers19. This might be due to the continuous slow release of the acidifier20. These results are in agreement with the findings of Cox et al.21, who showed that butyric acid in particular was effective in reducing Salmonella colonization of the intestine and Van Immerseel et al.7,10, who found that coated butyric acid was superior to uncoated butyric acid in reducing Salmonella colonization of the caeca and internal organs of chickens shortly after infection with S. enteritidis. Moreover, Zou et al.22 reported that the populations of Salmonellae, Escherichia coli and Clostridium perfringens in the caecum were decreased by supplementation of NaB.
Either scanning or transmission electron microscopy (SEM) or (TEM) examination of S. enteritidis strains re-isolated from the caeca of broiler chickens (with or without NaB supplementation) revealed some changes, according to the conditions of examination.
 | |
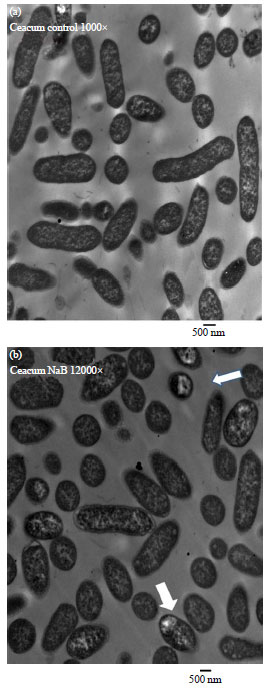
| Fig. 1(a-c): | Scanning Electron Microscopy (SEM) morphological features of examined Salmonella enteritidis isolated from the caecum of chicken broilers after 17 days of infection, (a) SEM of Salmonella enteritidis colonies re-isolated from the caecum of experimentally infected broilers without NaB supplementation (positive control group) and (b and c) SEM of Salmonella enteritidis colonies re-isolated from the caecum of experimentally infected broilers supplemented with NaB, notice the white arrows showing remnants of broken cells |
Figure 1 indicates that SEM of samples isolated from challenged NAB treated birds revealed some degraded and broken cells and the bacterial population looked aged for some extent. While the bacterial cells looked healthy in control group. The TEM of samples isolated from treated group showed transparent lipids like bodies, while normal features were observed in the untreated group (Fig. 2). Further investigations are necessary to understand the association between NaB treatment and detected changes in bacterial cells.
The pathogenesis of salmonellosis depends upon a large number of factors controlled by an array of genes those synergies into the actual virulence of Salmonella15. Nucleic acid based diagnostic techniques are being employed for the detection of various gene-encoded virulence factors23,24.
 | |
| Fig. 2(a-b): | Transmission Electron Microscopy (TEM) morphological features of examined Salmonella enteritidis isolated from the caecum of chicken broilers after 17 days of infection, (a) TEM of thin sections of Salmonella enteritidis colonies re-isolated from the caecum of experimentally infected broilers without NaB supplementation (positive control group) and (b) TEM of Salmonella enteritidis colonies re-isolated from the caecum of experimentally infected broilers supplemented with NaB, notice the transparent bodies are evident (white arrows). Scale bar = 500 nm |
Accordingly, in vivo assay using conventional PCR was conducted to express these genes of S. enteritidis strain after its re-isolation from livers of experimentally infected birds at 7th day post infection from treated or untreated groups with NaB.
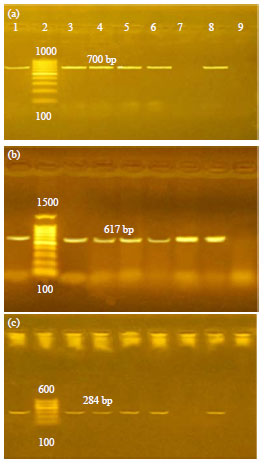 | |
| Fig. 3(a-c): | PCR results, (a) pefA gene: Positive amplification of 700 bp fragment was detected in three samples of group 4 (positive control) and two samples of groups 3 (NaB supplemented), (b) stn gene. Positive amplification of 617 bp fragment was detected in three samples of both groups 3 (NaB treated) and 4 (positive control) and (c) invA gene: Positive amplification of 284 bp fragment was detected in two samples of group 3 (NaB treated) and three samples of group 4 (positive control). Lane 1: Positive control (Salmonella eteritidis primary inoculum). Lane 2: Ladder. Lanes 3, 4, 5: Group 4 (Positive control Salmonella enteritidis reisolated from infected chickens). Lanes: 6,7, 8: Group 3 (Salmonella enteritidis reisolated from infected chickens treated with NaB). Lane 9: Negative control (E. coli) |
Three virulence genes were assayed including, Salmonella invasion (invA) Salmonella enterotoxin (stn) and plasmid encoded fimbrial (pefA) genes. In Fig. 3, S. enteritidis strains that re-isolated from untreated group (group 4) detected all these genes with an incidence of 100%. While modulation and deletion of pefA or stn genes were recorded in NaB treated strains (group 3) with an incidence of 33.33%.
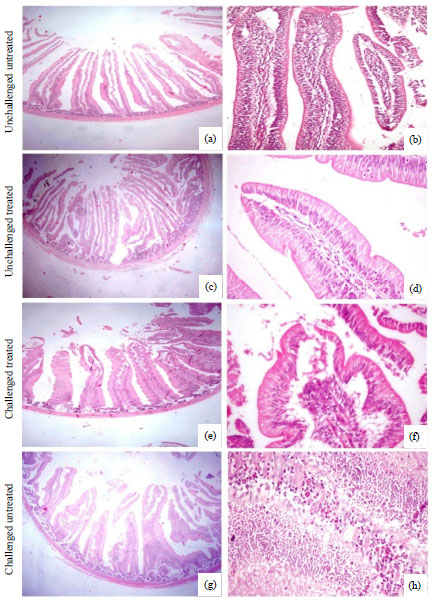 | |
| Fig. 4(a-h): | Small intestine of broiler chickens (a, c, e, g) Demonstrating the histomorphology of intestine in unchallenged untreated group, unchallenged NaB treated group, Salmonella enteritidis, challenged NaB treated group and Salmonella enteritidis challenged untreated group (X40), (b) Unchallenged untreated group showing normal histological structure on intestinal villi (X200) and (d) Unchallenged NaB treated group showing intact normal histological structure of enterocytes and lamina propria (X400), (f) Salmonella enteritidis challenged NaB treated group showing intact enterocytes with increased number of goblet cells and folding of intestinal surface with mild inflammatory reaction involving the lamina propria (X400), (h) Salmonella enteritidis challenged untreated group showing severe necrosis of enterocytes with massive heterophiles infiltration in lamina propria (X400) |
Durant et al.25 reported that SCFA modulate the expression of the hilA and invF genes of S. typhimurium. Boyen et al.26 found that some frequently used SCFA and medium-chain fatty acids were able to alter virulence gene expression and decrease S. typhimurium colonization and shedding in pigs using well established and controlled in vitro and in vivo assays. Modulation of these virulence genes in the present work might explain the antibacterial effect of NaB, a conclusion that could be partially confirmed by reports of Baloda et al.27 and Chopra et al.28 who mentioned that Salmonella induced diarrhoea was a complex phenomenon involving several pathogenic mechanisms including production of enterotoxin which was mediated by stn gene. Thorns et al.29 have shown that pef genes product play an important role in bacterial adhesion to the epithelial cells. Also, Csiko et al.30 found that butyrate, a commonly applied feed additive in poultry nutrition, could modify the expression of certain genes, including those encoding cytochrome P450 (CYP) enzymes.
Results of histopathological examination are seen in Table 1 and Fig. 4. Salmonella enteritidis challenged untreated group showed severe necrosis of enterocytes with massive heterophiles infiltration in lamina propria.
| Table 1: | Histomorphometric analysis of jejunal intestinal segments in different treated chicken groups |
 | |
| NaB: Sodium butyrate, Different superscripts are significantly different (p<0.05) and results are reported as Mean±SEM | |
On the other hand, challenged NaB treated group showed intact enterocytes with increased number of goblet cells besides folding of intestinal surface with mild inflammatory reaction involving the lamina propria. Brouns et al.31 and Pryde et al.32 mentioned that butyrate appeared to play a role in the development of the intestinal epithelial cells. Jiang et al.33 investigated the effects of micro-encapsulated sodium butyrate on oxidative stress and apoptosis induced by dietary corticosterone in the intestinal mucosa of broiler chickens. The protective effect of NaB supplementation in birds challenged with S. enteritidis was reflected on intestinal histomorphometric parameters as the presence of microbial load like S. enteritidis caused significant reduction in villous height, villous height: crypt depth ratio and significant (p≤0.05) increase in crypt depth that is not related to increase villous height as shown but related to increased cell turn over and sloughing induced by Salmonella. Accordingly, NaB has a protective effect against fast tissue turn over induced by S. enteritidis, relatively and partially increased villous height and villous height: crypt depth ratio as compared with NaB unchallenged group (group 2) that achieving significant (p≤0.05) increase in intestinal parameters and the final results reflected on villous height: crypt depth ratio to be similar to uninfected untreated group (normal histological structure of intestinal villi) (group 1). These results indicated that NaB supplementation achieved a beneficial effect on intestinal histomorphomatric parameters by increasing villous height and villous height: crypt depth ratio, which in turn positively might reflect on nutrient digestion and absorption and finally the body weight. Also, in group 2, NaB supplementation increased crypt depth that was beneficial whereas crypt depth is considered as a progenitor cells for villous epithelium and this may be assumed to be related to the demand for increase of the villous height and hence total increase in gut surface area. Mallo et al.34 concluded that inclusion of butyrate in the diet improves the digestibility of energy and protein by increasing intestinal absorption surface. Chamba et al.35 mentioned that jejunal villi of birds fed sodium butyrate and colistin at 42 days were higher than those in birds fed the control diet.
CONCLUSION
In conclusion, the used microencapsulated NaB in the present investigation had a bacteriocidal action against S. eneteritidis as it reduced its enumeration in cecum and liver and altered its morphology as well as modulated some of its virulence genes. It also played a positive role in the development of intestinal epithelial cells.
ACKNOWLEDGMENT
The authors appreciate the kind support of NUTRI-AD International, Belgium for supplying the product (Na-butyrate) encapsulated in palm fat under the trade name of Admix®30 that used in this study.
REFERENCES
- Mountzouris, K.C., P. Tsirtsikos, E. Kalamara, S. Nitsch, G. Schatzmayr and K. Fegeros, 2007. Evaluation of the efficacy of a probiotic containing Lactobacillus, Bifidobacterium, Enterococcus and Pediococcus strains in promoting broiler performance and modulating cecal microflora composition and metabolic activities. Poult. Sci., 86: 309-317.
CrossRefPubMedDirect Link - Abdalla, E.F., S.A. Nasef, M. El-Hariri and M. Refai, 2015. Genotypic characterization of antibiotic resistant Salmonella isolates recovered from local and imported poultry. J. Global Biosci., 4: 2611-2622.
Direct Link - Babu, U.S., K. Sommers, L.M. Harrison and K.V. Balan, 2012. Effects of fructooligosaccharide-inulin on Salmonella-killing and inflammatory gene expression in chicken macrophages. Vet. Immunol. Immunopathol., 149: 92-96.
CrossRefDirect Link - Chowdhury, R., K.M.S. Islam, M.J. Khan, M.R. Karim, M.N. Haque, M. Khatun and G.M. Pesti, 2009. Effect of citric acid, avilamycin and their combination on the performance, tibia ash and immune status of broilers. Poult. Sci., 88: 1616-1622.
CrossRefPubMedDirect Link - Awaad, M.H.H., A.M. Atta, M. Elmenawey, B. Shalaby and G.A. Abdelaleem et al., 2011. Effect of acidifiers on gastrointestinal tract integrity, zootechnical performance and colonization of Clostridium perfringens and aerobic bacteria in broiler chickens. J. Am. Sci., 7: 618-628.
Direct Link - Awaad, M.H.H., A. Atta, B. Shalaby, H.B. Gharieb and M. Elmenawey et al., 2014. The efficacy of Na-butyrate encapsulated in palm fat on experimentally induced necrotic enteritis and enumeration of intestinal resident clostridium perfringens in broiler chickens. IOSR J. Agric. Vet. Sci., 7: 40-44.
Direct Link - Van Immerseel, F., F. Boyen, I. Gantois, L. Timbermont and L. Bohez et al., 2005. Supplementation of coated butyric acid in the feed reduces colonization and shedding of Salmonella in poultry. Poult. Sci., 84: 1851-1856.
CrossRefPubMedDirect Link - Zhou, Z.Y., B. Packialakshmi, S.K. Makkar, S. Dridi and N.C. Rath, 2014. Effect of butyrate on immune response of a chicken macrophage cell line. Vet. Immunol. Immunopathol., 162: 24-32.
CrossRefDirect Link - Van Immerseel, F., V. Fievez, J. de Buck, F. Pasmans, A. Martel, F. Haesebrouck and R. Ducatelle, 2004. Microencapsulated short-chain fatty acids in feed modify colonization and invasion early after infection with Salmonella enteritidis in young chickens. Poult. Sci., 83: 69-74.
CrossRefPubMedDirect Link - NRC., 1994. Nutrient Requirements of Poultry. 9th Edn., National Academy Press, Washington, DC., USA., ISBN-13: 9780309048927, Pages: 176.
Direct Link - Thushani, W., K.W.S. Ariyawansa and P.C. Arampath, 2003. Recovering ability of freeze-stressed Salmonella typhimurium and Staphylococcus aureus cells in frozen shrimp. Cey. J. Sci., 31: 61-67.
Direct Link - Oliveira, S.D., C.R. Rodenbusch, M.C. Ce, S.L.S. Rocha and C.W. Canal, 2003. Evaluation of selective and non-selective enrichment PCR procedures for Salmonella detection. Lett. Applied Microbiol., 36: 217-221.
CrossRefPubMedDirect Link - Murugkar, H.V., H. Rahman and P.K. Dutta, 2003. Distribution of virulence genes in Salmonella serovars isolated from man and animals. Indian J. Med. Res., 117: 66-70.
PubMedDirect Link - Li, L., C.J. Stoeckert Jr. and D.S. Roos, 2003. OrthoMCL: Identification of ortholog groups for eukaryotic genomes. Genome Res., 13: 2178-2189.
CrossRefPubMedDirect Link - Favaro-Trindade, C.S. and C.R.F. Grosso, 2002. Microencapsulation of L. acidophilus (La-05) and B. lactis (Bb-12) and evaluation of their survival at the pH values of the stomach and in bile. J. Microencpasulat., 19: 485-494.
CrossRefPubMedDirect Link - Cox, N.A., F. McHan, J.S. Bailey and E.B. Shotts, 1994. Effect of butyric or lactic acid on the In vivo colonization of Salmonella typhimurium. J. Appl. Poult. Res., 3: 315-318.
CrossRefDirect Link - Prager, R., A. Fruth and H. Tschape, 1995. Salmonella enterotoxin (stn) gene is prevalent among strains of Salmonella enterica, but not among Salmonella bongori and other Enterobacteriaceae. FEMS Immunol. Med. Microbiol., 12: 47-50.
CrossRefDirect Link - Rahman, H., R. Prager and H. Tschape, 2000. Occurrence of sef and pef genes among different serovars of Salmonella. Indian J. Med. Res., 111: 40-42.
PubMed - Durant, J.A., D.E. Corrier and S.C. Ricke, 2000. Short-chain volatile fatty acids modulate the expression of the hilA and invF genes of Salmonella typhimurium. J. Food Protect., 63: 573-578.
Direct Link - Boyen, F., F. Haesebrouck, A. Vanparys, J. Volf and M. Mahu et al., 2008. Coated fatty acids alter virulence properties of Salmonella typhimurium and decrease intestinal colonization of pigs. Vet. Microbiol., 132: 319-327.
CrossRefDirect Link - Baloda, S.B., A. Faris, K. Krovacek and T. Wadstrom, 1983. Cytotonic enterotoxins and cytotoxic factors produced by Salmonella enteritidis and Salmonella typhimurium. Toxicon, 21: 785-796.
CrossRefDirect Link - Chopra, A.K., C.W. Houston, J.W. Peterson, R. Prasad and J.J. Mekalanos, 1987. Cloning and expression of the Salmonella enterotoxin gene. J. Bacteriol., 169: 5095-5100.
Direct Link - Thorns, C.J., C. Turcotte, C.G. Gemmell and M.J. Woodward, 1996. Studies into the role of the SEF14 fimbrial antigen in the pathogenesis of Salmonella enteritidis. Microbial Pathogen., 20: 235-246.
CrossRefPubMedDirect Link - Csiko, G., G. Nagy, G. Matis, Z. Neogrady and A. Kulcsar et al., 2014. Effects of dietary sodium butyrate on hepatic biotransformation and pharmacokinetics of erythromycin in chickens. J. Vet. Pharmacol. Therapeut., 37: 406-412.
CrossRefDirect Link - Brouns, F., B. Kettlitz and E. Arrigoni, 2002. Resistant starch and the butyrate revolution. Trends Food Sci. Technol., 3: 251-261.
CrossRefDirect Link - Pryde, S.E., S.H. Duncan, G.L. Hold, C.S. Stewart and H.J. Flint, 2002. The microbiology of butyrate formation in the human colon. FEMS Microbiol. Lett., 217: 133-139.
CrossRefDirect Link - Jiang, Y., W.H. Zhang, F. Gao and G.H. Zhou, 2014. Micro-encapsulated sodium butyrate attenuates oxidative stress induced by corticosterone exposure and modulates apoptosis in intestinal mucosa of broiler chickens. Anim. Prod. Sci., 55: 587-594.
CrossRefDirect Link - Mallo, J.J., M. Puyalto and S.V.R. Rao, 2011. Evaluation of the effect of sodium butyrate addition to broilers diet on energy and protein digestibility, productive parameters and size of intestinal villi of animals. Proceedings of the 48th AECA Scientific Poultry Symposium, November 4, 2011, Australia, pp: 343-349.
- Chamba, F., M. Puyalto, A. Ortiz, H. Torrealba, J.J. Mallo and R. Riboty, 2014. Effect of partially protected sodium butyrate on performance, digestive organs, intestinal villi and E. coli development in broilers chickens. Int. J. Poult. Sci., 13: 390-396.
CrossRefDirect Link