
Hassan A. Khalil
Department of Animal Production, Faculty of Agriculture, Suez Canal University, 41522, Ismailia, Egypt
Mohamed A. Yaseen
Department of Animal Production, Faculty of Agriculture, Suez Canal University, 41522, Ismailia, Egypt
Akrum M.M. Hamdy
Department of Animal Production, Faculty of Agriculture, Minia University, Minia, Egypt
Asian Journal of Poultry Science
Year: 2015 | Volume: 9 | Issue: 4 | Page No.: 191-202







ABSTRACT
This study was conducted to investigate the effect of acute short term heat exposure on behavioral activities, physiological body reactions, hematological parameters and hormonal profiles of New Zealand White (NZW) and Baladi Red (BR) bucks. Twenty male NZW and BR bucks were divided into two groups; natural winter climate of 19±1°C and 55±5%, Relative Humidity (RH) (control group) and the bucks in the other group were exposed to short term heat stress for 1 h at 37±0.5°C and 20±2% RH. The results indicated that, heat stress decreased (p≤0.001) standing and walking behavior and increased (p≤0.001) sitting behavior compared with these recorded under control temperature. Moreover, Respiration Rate (RR), Rectal Temperatures (RT) and Time of Sexual Libido (TSL) were significantly increased (p≤0.001) following stress. Concentration of plasma testosterone and T3 were significantly reduced (p≤0.001) and cortisol were significantly increased after submission to stress. The RBCs, WBCs, Hb, PCV values were insignificantly reduced and lymphocyte was significantly reduced after stress. In contrast, neutrophils, Neutrophils/Lymphocytes (N/L) ratio were increased (p≤0.05) and monocytes were insignificantly increased after the end of heat stress. On the other hand, breeds of rabbit had significant effects on most of the studied traits, especially after heat exposure. The BR bucks were significantly superior; RBCs, WBCs, neutrophils, N/L ratio and monocytes than those recorded in NZW bucks after heat stress. Also, plasma concentration of testosterone were significantly higher but cortisol and T3 were significantly lower (p≤0.05) in BR bucks than those estimated in NZW bucks. Moreover, BR bucks had insignificantly higher walking and lower standing behavior than those recorded in NZW bucks. Also, RR was significantly higher (p≤0.001), but RT and TSL were significantly lower (p≤0.001) in BR bucks than those recorded in the NZW bucks. Furthermore, the results showed that there were prevalent significant (p≤0.05) negative correlation between plasma level of cortisol and each of plasma level of testosterone, RBCs, WBCs counts, standing and walking behavior and significant (p≤0.001) positive with RR, RT, TSL and time of sitting behavior. Current results explained that acute short term heat stress, negatively affected of internal environment of rabbit bucks which were reflected in their performance. Also, behavioral traits, sexual libido, hematological parameters and hormonal profile in BR bucks seem to be better than those recorded by NZW bucks after exposed heat stress, which may be explained, why BR breed had more tolerate to heat stress compared to exotic breeds.
PDF Abstract XML References Citation
Received: July 01, 2015;
Accepted: August 07, 2015;
Published: September 15, 2015
How to cite this article
Hassan A. Khalil, Mohamed A. Yaseen and Akrum M.M. Hamdy, 2015. Behavioral Activities, Physiological Body Reactions, Hematological Parameters and Hormonal Profiles for Bucks of New Zealand White and Baladi Red Rabbits Exposed to Short Term of High Temperature. Asian Journal of Poultry Science, 9: 191-202.
DOI: 10.3923/ajpsaj.2015.191.202
URL: https://scialert.net/abstract/?doi=ajpsaj.2015.191.202
DOI: 10.3923/ajpsaj.2015.191.202
URL: https://scialert.net/abstract/?doi=ajpsaj.2015.191.202
INTRODUCTION
Rabbits, as a homeothermic animal, can regulate their body temperature by using physical, morphological, biochemical and behavioral processes (Fayez et al., 1994). The thermo-neutral zone temperature in rabbits is around 18-21°C (Habeeb et al., 1999). High temperatures, as encountered in Egypt and in many other countries during summer represent a major constraint factor for rabbit production (Askar and Ismail, 2012). Negative effects of prolonged thermal stress on functional parameters (Okab et al., 2008), on physiological performance (Kishik et al., 2009), on immune system response (Ondruska et al., 2011), on hormonal profile (Kowalska et al., 2011), physiological body reaction (Askar and Ismail, 2012) and on sexual behavior (Khalil et al., 2014). In rabbits have been investigated in recent years. These investigations focused the effects of long term reaction of rabbits to chronic thermal stress conditions. However, there is no evidence that acute thermal stress induces behavioral and physiological parameters changes during, acute short-term heat stress. On the other hand, some studies reported that climatic heat stress had deleterious effect on exotic breeds, such as New Zealand White (NZW) rabbits more than indigenous native breeds, such as Egyptian Baladi rabbits in productive and reproductive traits (Safaa et al., 2008; Khalil et al., 2014). Moreover, internal physiological changes which increase heat tolerance of Baladi breed compared to exotic breeds are poorly documented. Therefore, the aims of the present study were to evaluate; (i) the influence of acute short heat stress on behavioral traits, physiological body reaction, hematological parameters and hormonal profile, (ii) comparative between physiological responses of two breeds, NZW and Baladi Red (BR) bucks towards acute short term heat stress.
MATERIALS AND METHODS
Animals and experimental design: This experiment was carried out at the rabbitry experimental farm belonging to Faculty of Agriculture, Suez Canal University, Ismailia, Egypt. Rabbits were kept continuously under the same managerial and environmental conditions during, the whole experimental period. Twenty mature males of each of NZW and BR bucks were used in this study. All bucks were healthy and free of any external parasites or skin diseases. They were at age between 10-12 months and their weight averaged 3.125 kg. All bucks were individually housed in galvanized wired cages, where feed and water were provided ad libitum. Provided diet was contained 17% crude protein, 2.8% fat, 10% crude fiber and 2600 kcal digestible energy/kg diet. Lighting system was 16 h light/8 h dark in the rabbitry during the experimental period. The experimental period extended for 4 weeks during, February, 2015.
All the bucks were placed individually in two climatic cages. Each cage forming from galvanized metal (100×50×50 cm). The ambient temperature and relative humidity were 19±1°C and 55±5%, respectively, for control group, which contain ten bucks from each breed. The environmental conditions of the first cage were the same rabbitry conditions. While the bucks in the second group (10 bucks from each breed) were housed in climatic cage. Climatic cage includes; heater, thermostat, hygrometer and thermometer. The ambient temperature and relative humidity were 37±0.5°C and 20±2% RH, respectively. This temperature was applied daily for 1 h between 8-12 am.
Experimental studied traits
Behavioral activities: The behavior traits of all bucks were recorded in control and stressed bucks using a video camera. The behavior activities traits were recorded for each buck individually, throughout 1 h during the morning from 9-11 am. The following traits were determined for each 5 min interval (sampling time), number of bucks standing, sitting or walking. Numbers of observations were calculated, as a percentage of bucks observed for each trait (Khalil et al., 2012).
Physiological body reactions and time of sexual libido: Thermo-respiratory responses including Rectal Temperatures (RT), Respiration Rate (RR) and Time of Sexual Libido (TSL) were recorded before and after exposed to short term heat stress. RR was measured in bucks by counting the movements of the chest muscles for 1 min. The RT was measured by using clinical thermometer, which inserted for 1 min into the rectum for about 2 cm from anus orifice after liniment by pure Vaseline (Khalil et al., 2014). The TSL was measured in terms of reaction time in seconds and was estimated between introducing the teaser doe to the buck's cage up to the point, when the buck started to mount the doe (Elkomy et al., 2015).
Hematological parameters and hormonal profile: Blood samples were collected before and after heat stress of each animal from ear vein into heparinized tubes and was placed immediately on ice box. Plasma was obtained by blood centrifugation at 3000 rpm for 20 min and stored at -20°C until analysis. The whole blood was subjected to hematological analysis shortly after collection. Red Blood Cells (RBCs) and White Blood Cells (WBCs) counts, hemoglobin concentration (Hb) and Packed Cell Volume (PCV) were measured and leucocytes were differentiated, using NIHON KOHDEN equipment (automated hematology analyzer). Also, plasma cortisol, testosterone and T3 were analyzed by ELISA kits manufactured by DiaMetra, Spello-Perugia, Italy.
Statistical analysis: Data were analyzed using the General Linear Model (GLM) procedure of SAS (SAS., 2001). Differences among means were detected using Duncan’s new multiple test (Duncan, 1955). Correlation coefficients among traits were estimated.
Two-way analysis of variance was carried out for all traits using the following model:
Where:
| Yijk | = | Observation on the kth individual from the ith temperature in jth breed |
| μ | = | Overall mean |
| Ti | = | Fixed effect of the ith temperature |
| Bj | = | Fixed effect of the jth breed |
| Tij | = | Interaction between ith temperature and jth breed |
| eijk | = | Random error associated with the ijkth individual |
RESULTS
Behavioral activities: The effects of different temperatures and breeds on behavioral activities are presented in Table 1. Short term heat stress increased significantly (p≤0.001) sitting percent and decreased significantly (p≤0.001), standing and walking percent compared with those recorded under control temperature. On the other hand, behavioral traits did not affected significantly by breed. However, BR had higher walking percent and lower standing percent than NZW bucks. Moreover, there were significant (p≤0.001) interaction between temperatures and breeds on behavioral activities. Both breeds kept in control temperature had the longest time standing and walking percent but the shortest time were obtained in both breed, which kept under heat stress. In contrast, time of sitting percent was lower in both breeds kept under control temperature than under heat stress.
| Table 1: | Behavioral activities of rabbit bucks as affected by treatments |
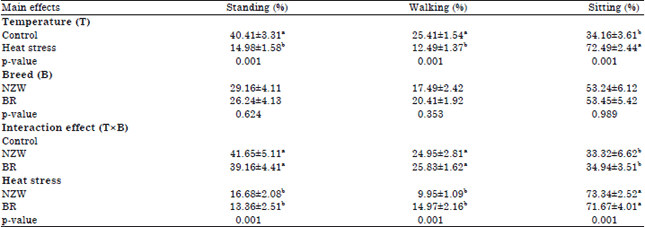 | |
| a,bMean within a column and within a source not sharing a common superscript differed significantly (p≤0.05), values are given in Mean±SE | |
| Table 2: | Respiration rate (breaths/m), rectal temperature (°C) and time of sexual libido (Sc) of rabbit bucks as affected by treatments |
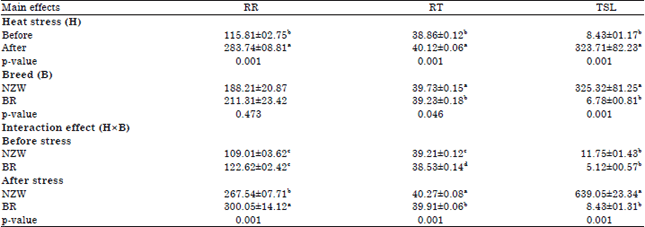 | |
| a,bMean within a column and within a source not sharing a common superscript differed significantly (p≤0.05), values are given in Mean±SE, RR: Respiration rate, RT: Rectal temperature, TSL: Time of sexual libido | |
Physiological body reactions and time of sexual libido (TSL): Results in Table 2 showed that treatments had a significant effect on physiological body reactions (RR and RT) and Time of Sexual Libido (TSL). Rabbit bucks after exposed to heat stress had significantly superior (p≤0.001) RR, RT and TSL than those recorded before heat stress. Baladi Red rabbit bucks had significantly shorter time of sexual libido (p≤0.001) and significantly lower rectal temperature (p≤0.046) and insignificantly higher respiration rate (p = 0.473) than those recorded in NZW rabbits bucks. Moreover, there was a significant interaction between treatments on the same traits. The highest RR (p≤0.001) was recorded in BR bucks after heat stress compared with all experimental groups. Also, the highest rectal temperature (p≤0.001) was obtained in NZW bucks after heat stress but the lowest value was obtained in BR bucks before heat stress. The TSL was significantly delayed (p≤0.001) in NZW bucks after heat stress compared with all experimental groups.
Hematological parameters: Results in Table 3 indicated that neutrophils and Neutrophils/Lymphocytes (N/L) ratio were significantly increased (p≤0.05) and monocytes were insignificantly increased (p = 0.058) after heat stress compared to before heat stress. However, heat stress caused insignificantly reduced RBCs, Hb, PCV, WBCs and eosinophils values compared to before stress.
| Table 3: | Blood cell count and hemoglobin concentration of rabbit bucks as affected by treatments |
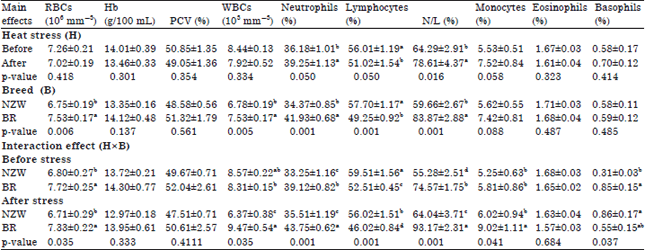 | |
| a,bMean within a column and within a source not sharing a common superscript differed significantly (p≤0.05), values are given in Mean±SE, RBCs: Red blood cells, Hb: Hemoglobin, WBCs: White blood cells, PCV: Packed cell volume, N/L: Neutrophils/lymphocytes | |
Moreover, BR bucks had a significantly (p≤0.01) increased RBCs, WBCs, neutrophils, lymphocytes and N/L ratio and insignificantly increased Hb, PCV and monocytes in comparison with those recorded in NZW rabbit bucks after stress. The interaction between treatments had a significant (p≤0.05) effect on most of these traits. The highest values of neutrophils, N/L ratio and monocytes were found in BR after stress but the lowest values were recorded in NZW before stress. Also, the highest RBCs counts were found in BR but the lowest value was obtained in NZW before and after exposed to high temperature. Also, the highest WBCs count was recorded in BR but the lowest value was recorded in NZW after stress.
Plasma hormonal profile: Highly significant (p≤0.001) differences were found between treatments in most hormonal profile (Table 4). Plasma cortisol was significantly increased but testosterone and T3 concentration were significantly decreased in rabbit bucks after exposed to heat stress compared with those estimated before heat stress. Moreover, BR bucks had significantly higher (p≤0.001) concentration of plasma testosterone and significantly lower (p≤0.001) cortisol and T3 (p≤0.036) than those obtained in NZW rabbit bucks. On the other hand, plasma hormones levels were significantly affected by the interactions among treatments. The highest levels of cortisol and testosterone were recorded in NZW bucks after stress and in BR bucks before heat stress, but the lowest values were obtained in BR bucks before stress and in NZW bucks after stress, respectively. Moreover, plasma concentration of T3 was significantly decreased (p≤0.004) in BR rabbit bucks after stress compared with those obtained in other treatment groups.
Correlation coefficients among studied traits: The results showed that there were prevalent significant (p≤0.05) negative correlation between plasma level of cortisol and each of plasma level of testosterone, RBCs, WBCs and time of standing and walking values. Also, similar trend was observed between plasma level of testosterone and each of RR, RT, TSL and time of sitting values. Also, between both RR and RT and both time of standing and walking behavior. In contrast, significant (p≤0.001) positive correlations were found between plasma level of cortisol and each of RR, RT, TSL and time of sitting values. Moreover, plasma level of testosterone and each of RBCs, WBCs and time of walking values showed positive correlation coefficients. As well as between RR value and each of RT, TSL and time of sitting values (Table 5).
| Table 4: | Plasma levels of cortisol, testosterone and T3 of rabbit bucks as affected by treatments |
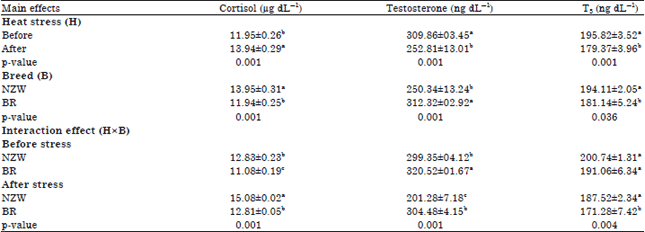 | |
| a,bMean within a column and within a source not sharing a common superscript differed significantly (p≤0.05), values are given in Mean±SE | |
| Table 5: | Correlation coefficients among some studied traits |
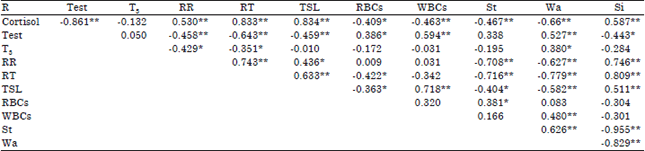 | |
| R: Correlation coefficient, Test: Testosterone, RR: Respiration rate, RT: Rectal temperature, TSL: Time of sexual libido, St: Standing (%), Wa: Walking (%), Si: Sitting (%), *p≤0.05, **p≤0.01 | |
DISCUSSION
Results of the present study showed clearly that short time heat exposure and breeds of rabbits affected significantly on most the studied traits. Short term heat stress decreased significantly time of standing and walking behavioral activities and increased significantly time of sitting compared with those recorded under control temperature. These results are similar with the results obtained by Khalil et al. (2012), who recorded that high housing temperature has adversely effect on behavior activities in Japanese quail. He added that time of sitting was significantly superior under heat stress than those recorded under control temperature. The reason of increase in time of sitting behavior during heat stress may be due to try the animals to increase heat dissipation by conduction to the floor of the cage. The design of rabbit housing is governed by the behavioral characteristics of the animals and their reactions to environmental temperature and humidity.
Moreover, short term heat stress increased significantly RR, RT and TSL compared with those recorded before stress. These results are in a good agreement with previous reports by Kishik et al. (2009), Kowalska et al. (2011) and Askar and Ismail (2012), who reported that high ambient temperature caused a significant increase in RR and RT as compared with those recorded under comfort conditions. Thermoregulation in rabbits is rather poor as they have few functional sweat glands (Naqvi et al., 1995). Marai et al. (2001) reported that the comfort zone temperature for rabbits is around 21°C. When, the air temperature rises beyond the upper limit of thermo-neutrality range, physical regulation of body temperature is insured by the adjustment of blood flow to skin and by the perspiration mechanisms. Vasomotor and cardio-respiratory mechanisms are also involved, in addition to other physiological mechanisms (El-Sobhy, 2000). Rabbits do not tolerate heat well and can actually die from being overheated. Each rabbit will tolerate heat differently and it is important to observe rabbit daily. Rabbits with thick or long coats of hair, overweight and young or old are at an even greater risk. Temperature, humidity and air ventilation are all factors that contribute to heatstroke in a rabbit. Rabbits are individuals and could respond to these conditions somewhat differently. It is important to check the rabbits consistently to insure they are comfortable and do not overheat. Early detection of heatstroke and proper corrective steps could mean the difference between life and death for rabbits.
On the other hand, short term heat stress decreased the values of RBCs, Hb, PCV, WBCs, eosinophils (p>0.05) and lymphocytes (p≤0.05) and increased neutrophils, N/L ratio (p≤0.05) and monocytes and basophils (p>0.05) compared with these values recorded before heat stress. These results are in agreement with other researchers (Okab et al., 2008; Kishik et al., 2009; Ondruska et al., 2011; Askar and Ismail, 2012), who found that high ambient temperature had adversely affected in most hematological parameters in rabbits. They added that the values of RBCs, Hb and lymphocytes were lower but neutrophils and monocytes were higher under high temperature than those recorded under control temperature. Also, El-Lethey et al. (2003) found that when used chronic stress with corticosterone feeding caused higher N/L ratios in laying hens. Moreover, Kadam et al. (2009) reported that N/L ratio is reliable indicator of stress and added that neutrophil and N/L ratio increased during hot season in broiler chicken.
The reduction in hematological parameters especially RBCs may be returned to two reasons: 1) reduce oxygen carrying by RBCs to reduce metabolic heat production under this hot condition (Ashour, 2001) and 2) lysis of RBCs during heat stress. Abdul Wahab et al. (2010) reported that heat stress induced lysis of RBCs during hot season in Wister rats. Therefore, RBCs are frequently used to evaluate oxidative stress. Because the high lipid content and iron and oxygen rich intracellular environment of RBCs makes them highly susceptible to auto-oxidation and radical-mediated damage (Jani et al., 2012). Also, membrane of RBCs is rich in polyunsaturated fatty acids, a primary target for reaction involving free radical and is very susceptible to lipid peroxidation (Brzezinska-Slebodzinska, 2003; Mahmoud, 2013). Peroxidation results in the loss of membrane fluidity and cellular lysis. Moreover, increased hemolysis has been used as an indirect way of quantifying oxidative stress in animals during stress (Adenkola and Ayo, 2009).
Also, numerous studies have shown that stress and stress hormones (catecholamines and the glucocorticoids) induce significant changes in absolute numbers and relative proportions of WBCs in the blood (Redwine et al., 2003). The WBCs responses or lymphoid organ regressions are considered as better indicators of chronic stress (Siegel, 1995). Dhabhar et al. (1995) reported that stress causes decreased blood WBCs numbers. This reduction in WBCs may be interpreted in two possible ways, 1) it could reflect a large-scale destruction of circulating WBCs (Cohen, 1992) and 2) it could reflect a redistribution of leukocytes from the blood to other organs such as the skin and lining of the gastrointestinal and urinary-genital tracts and draining sentinel lymph nodes. This redistribution may enhance immune function in those compartments to which immune cells traffic during stress (Toft et al., 1993). On the other hand, Dhabhar et al. (1995) reported that stress reduced significantly numbers and percentages of lymphocytes by an increase in numbers and percentages of neutrophils. Also, corticosteroids and catecholamine-induced increased accumulation of lymphocytes in the spleen, lymph nodes and mucosal sites which decrease in lymphocytes in the blood (Viswanathan and Dhabhar, 2005). In contrast, some studies have shown that stress increased blood WBCs numbers in rats (Harris et al., 1995) and humans (Bosch et al., 2003).
In the same context, short term heat stress led to significantly increased plasma cortisol and decreased plasma testosterone and T3 levels compared with those recorded before heat stress. Our results confirmed with Kowalska et al. (2011), who reported that the level of blood cortisol hormone was increased in NZW rabbit when exposed to thermal stress. Also, with Koko et al. (2004), who reported that 60 min heat (38°C) exposure of rats induced a significant rise in circulating ACTH and glucocorticoids levels.
In this context, Kalaba (2012) reported that increasing cage density of growing rabbits induced stress effects which increased level of blood cortisol. Also, Tanchev et al. (2014) recorded that road transportation leads to an increase in the levels of ACTH and cortisol in blood of California rabbits compared with before transportation. Cortisol, as a representative glucocorticoid is produced in the zona fasciculata of the adrenal cortex and is needed in times of stress to maintain blood glucose levels and prevent shock. Cortisol regulates its own secretion by a negative feedback effect on the hypothalamic-pituitary-adrenal axis (Burtis et al., 2006). It has been shown that temperature is an important factor in the regulation of endocrine hormone release (Squires, 2003). Our results showed that there were a negative correlation between plasma level of cortisol and most traits, such as, RBCs, WBCs, time of standing and walking behavior values and levels of plasma testosterone and T3.
In contrary, the results showed that short term heat stress decreased significantly plasma testosterone level compared with those estimated before heat stress. This result consistent with Chiericato et al. (1995) and El-Hanoun et al. (2007), who recorded that testosterone was significantly decreased in male rabbits exposed to heat stress compared to control conditions. Testosterone is a steroid hormone from the androgen group in mammals, it is primarily is formed and secreted from Leydig cells in the testicles of males and the ovaries of females, although, small amounts are also secreted by the adrenal glands (Burtis et al., 2006). Testosterone the main circulating androgen of testicular origin is vital in maintaining libido, body fat, maintaining muscle mass, bone density and the regulation of blood sugar (Yahaya and Ajuogu, 2014). Our results showed that there was negative correlation between levels of plasma cortisol and plasma testosterone. These results are in agreement with Brownlee et al. (2005), who reported that there are a negative relationship between cortisol hormone and total blood testosterone in human. Moreover, Bambino and Hsueh (1981) showed a direct inhibitory effect of high doses of glucocorticoids upon testicular Leydig cell function in rats and in humans (Cumming et al., 1983). In this context, short term heat exposure decreased significantly plasma T3 level compared with those recorded before stress. These results are in agreement with Chiericato et al. (1995), who recorded that T3 was significantly decreased in male rabbits exposed to heat stress compared to control conditions. Also, Marai et al. (2004) recorded that T3 decreased in growing rabbits under hot conditions compared to mild conditions. This reduction may be due to an attempt the animals to reduce heat production under heat stress.
Moreover, short term heat exposure had adverse effect on TSL, which more delayed significantly after exposed to heat stress compared with before stress. This result are in agreement with Kishik et al. (2009), who recorded that time of sexual libido was faster under winter conditions than in summer conditions in NZW bucks. Our results showed that there was a positive correlation between plasma testosterone and TSL. These results are in agreement with El-Hanoun et al. (2014), who reported that a good relation between increased blood testosterone concentration and increase libido of male rabbits.
On the other hand, our results indicated that breed of rabbits had significantly affected in most the studied traits especially after exposed to heat stress. The BR bucks had significantly higher RR, plasma testosterone level, WBCs count, neutrophils (%), N/L ratio and monocytes (%) and significantly lower RT, TSL values, lymphocytes (%) than those recorded in NZW bucks after exposed to heat stress. These results are in agreement with the results obtained by Safaa et al. (2008) and Khalil et al. (2014), who recorded that climatic heat stress had deleterious effect on exotic breeds, such as NZW rabbits more than indigenous breeds, such as Egyptian Baladi rabbits. Safaa et al. (2008) recorded that NZW bucks were higher sensitivity to temperature changes than Baladi Black (BB) bucks. He added that TSL and semen quality were the worst in NZW bucks compared to BB bucks under high temperature. Also, El-Azim and El-Kamash (2011) reported that TSL in Baladi bucks was faster than that of NZW under environmental conditions in the middle of Egypt. The faster of sexual libido in Baladi bucks compared with NZW bucks may be due to the direct increased in plasma testosterone and its action. Our results indicated that plasma testosterone level had a significant positive correlation with faster TSL, which seems higher in BR bucks compared with NZW bucks. These results are in agreement with Sharpe et al. (1988), who found that male sexual desire and spermatogenesis process depends on the action of testosterone. In contrast, NZW bucks had significantly higher plasma cortisol and T3 than those recorded in BR bucks breed after heat stress. This confirms that NZW bucks are less adapted to high heat exposure than BR bucks. Kalaba (2012) who reported that increased level blood cortisol due to negative effect in physiological parameters in rabbits.
CONCLUSIONS
In conclusion, results of this study indicated that acute short term heat exposure had negatively affected of internal environment of bucks especially plasma hormonal concentration, which were reflected in their performance. Also, behavioral traits, sexual libido, hematological parameters and hormonal profile seem to be better in BR bucks than those recorded by NZW bucks after heat stress, which may be explained why BR breed had more tolerate to heat stress compared to exotic breeds.
ACKNOWLEDGMENTS
The authors would like to thank Prof. Dr. Mostafa A. Ayoub "Head of Animal Production Department, Faculty of Agriculture, University of Suez Canal" for his efforts during this study.
REFERENCES
- Adenkola, A.Y. and J.O. Ayo, 2009. Effect of road transportation on erythrocyte osmotic fragility of pigs administered ascorbic acid during the harmattan season in Zaria, Nigeria. J. Cell Anim. Biol., 3: 4-8.
Direct Link - Mahmoud, A.A., 2013. Protective effect of myricetin on proteins and lipids of erythrocytes membranes. Asian J. Biol. Sci., 6: 76-83.
CrossRefDirect Link - Elkomy, A.E., A.M. Abd El-hady and O.A. Elghalid, 2015. Dietary boron supplementation and its impact on semen characteristics and physiological status of adult male rabbits. Asian J. Poult. Sci., 9: 85-96.
CrossRefDirect Link - Abdul Wahab, A., M.A.A. Mabrouk, J.O. Ayo, A.F. Sulaimon, S. Muftau, A.A. Yahaya and S.E. Oluwatobi, 2010. Effects of co-administration of antioxidants on erythrocyte osmotic fragility of wistar rats during the hot-dry season. Eur. J. Sci. Res., 46: 73-79.
Direct Link - El-Azim, A.A. and E.M. El-Kamash, 2011. Evaluation of semen quality and its relation to mating system for some breeds of rabbits under environmental conditions in the middle of Egypt. Egypt. Poult. Sci., 31: 467-480.
Direct Link - Askar, A.A. and E.I. Ismail, 2012. Impact of heat stress exposure on some reproductive and physiological traits of rabbit does. Egypt. J. Anim. Prod., 49: 151-159.
Direct Link - Bambino, T.H. and A.J.W. Hsueh, 1981. Direct inhibitory effect of glucocorticoids upon testicular luteinizing hormone receptor and steroidogenesis in vivo and in vitro. Endocrinology, 108: 2142-2148.
CrossRefDirect Link - Bosch, J.A., G.G. Berntson, J.T. Cacioppo, F.S. Dhabhar and P.T. Marucha, 2003. Acute stress evokes selective mobilization of T cells that differ in chemokine receptor expression: A potential pathway linking immunologic reactivity to cardiovascular disease. Brain Behav. Immunity, 17: 251-259.
CrossRefDirect Link - Brzezinska-Slebodzinska, E., 2003. Species differences in the susceptibility of erythrocytes exposed to free radicals in vitro. Vet. Res. Commun., 27: 211-217.
CrossRefDirect Link - Chiericato, G.M., C. Boiti, C. Canali, C. Rizzi and L. Ravarotto, 1995. Effects of heat stress and age on growth performance and endocrine status of male rabbit. World Rabbit Sci., 3: 125-131.
CrossRefDirect Link - Cohen, J.J., 1992. Glucocorticoid-induced apoptosis in the thymus. Semin. Immunol., 4: 363-369.
PubMedDirect Link - Cumming, D.C., M.E. Quigley and S.S.C. Yen, 1983. Acute suppression of circulating testosterone levels by cortisol in men. J. Clin. Endocrinol. Metab., 57: 671-673.
CrossRefDirect Link - Dhabhar, F.S., A.H. Miller, B.S. McEwen and R.L. Spencer, 1995. Effects of stress on immune cell distribution. Dynamics and hormonal mechanisms. J. Immunol., 154: 5511-5527.
PubMedDirect Link - Kowalska, D., A. Gugolek and P. Bielanski, 2011. Effect of stress on rabbit meat quality. Ann. Anim. Sci., 3: 465-475.
Direct Link - El-Hanoun, A.M., H. El-Saieed, M.S. El-Sbeiy and K.I. Kamel, 2007. Effect of bee pollen supplementation on some productive, reproductive and biochemical traits of growing male rabbits during winter and summer seasons. Proceedings of the 5th International Conference on Rabbit Production in Hot Climates, December 4-7, 2007, Hurghada, Egypt, pp: 417-433.
- El-Hanoun, A.M., A.E. Elkomy, W.A. Fares and E.H. Shahien, 2014. Impact of royal jelly to improve reproductive performance of male rabbits under hot summer conditions. World Rabbit Sci., 22: 241-248.
CrossRefDirect Link - Habeeb, A.A.M., A.M. El-Maghawry, I.F.M. Marai and A.E. Gad, 1999. Physiological thermoregulation mechanism in rabbits drinking saline water under hot summer conditions. Proceedings of the 1st International Conference on Indigenous Versus Acclimatized Rabbits, September 7-9, 1999, El-Arish, North Sinai, Egypt, pp: 443-456.
- Harris, T.J., T.J. Waltman, S.M. Carter and A.S. Maisel, 1995. Effect of prolonged catecholamine infusion on immunoregulatory function: Implications in congestive heart failure. J. Am. Coll. Cardiol., 26: 102-109.
CrossRefDirect Link - El-Lethey, H., B. Huber-Eicher and T.W. Jungi, 2003. Exploration of stress-induced immunosuppression in chickens reveals both stress-resistant and stress-susceptible antigen responses. Vet. Immunol. Immunopathol., 95: 91-101.
CrossRefDirect Link - Kadam, A.S., V.D. Lonkar, V.R. Patodkar, S.M. Kolangath and T.A. Bhosale, 2009. Comparative efficacy of supplementation of natural (Citrous limon Juice), herbal and synthetic vitamin c on the immune response of broiler chicken during summer stress. Asian J. Poult. Sci., 3: 57-62.
CrossRefDirect Link - Kalaba, Z.M., 2012. Physiological response and stress indicators of California rabbits under intensive conditions in Egypt. Asian J. Poult. Sci., 6: 65-78.
CrossRef - Brownlee, K.K., A.W. Moore and A.C. Hackney, 2005. Relationship between circulating cortisol and testosterone: Influence of physical exercise. J. Sports Sci. Med., 4: 76-83.
Direct Link - Khalil, H.A., M. Gerken, A.M. Hassanein and M.E. Mady, 2012. Behavioural responses of two Japanese quail lines differing in body weight to heat stress. Egypt. J. Anim. Prod., 47: 151-158.
Direct Link - Khalil, H.A., W.H. Kishk, O. Essa and M.M. Awad, 2014. Evaluation of productive and physiological performance of Baladi red compared to New Zealand white rabbits under the same managerial conditions. Egypt. J. Anim. Prod., 51: 200-209.
Direct Link - Kishik, W.H., R.A. Tharwat, H.A. Khalil and M.A. Ayoub, 2009. Seasonal effects on White New Zealand and Californian rabbit bucks semen characteristics, hematological parameters and body reaction performance in Egypt. Proceedings of the 5th International Poultry Conference, March 10-13, 2009, Taba, Egypt, pp: 1582-1590.
- Fayez, I., M. Marai, A. Alnaimy and M. Habeeb, 1994. Thermoregulation in rabbits. CIHEAM-Options Mediterraneennes, 8: 33-41.
Direct Link - Marai, I.F.M., A.A.M. Haeeb and A.E. Gad, 2004. Growth performance traits and the physiological background of young doe rabbits as affected by climatic conditions and lighting regime, under sub-tropical conditions of Egypt. Proceedings of the 8th World Rabbit Congress, September 7-10, 2004, Pueblo, Mexico, pp: 288-297.
- Marai, I.F.M., M.S. Ayyat and U.M. Abd El-Monem, 2001. Growth performance and reproductive traits at first parity of New Zealand white female rabbits as affected by heat stress and its alleviation under Egyptian conditions. Trop. Anim. Health Prod., 33: 451-462.
CrossRefDirect Link - Naqvi, S.M.K., R. Gulyani and G. Singh, 1995. Physiological responses of broiler rabbits in hot semi-arid environment. Indian J. Anim. Sci., 65: 718-720.
Direct Link - Jani, N., J. Ziogas, J.A. Angus and C.E. Wright, 2012. Exogenous glutathione is essential in the testing of antioxidant capacity using radical-induced haemolysis. J. Pharmacol. Toxicol. Methods, 65: 142-146.
CrossRefDirect Link - Okab, A.B., S.G. El-Banna and A.A. Koriem, 2008. Influence of environmental temperatures on some physiological and biochemical parameters of New Zealand rabbits male. Slovak J. Anim. Sci., 41: 12-19.
Direct Link - Ondruska, L., J. Rafay, A.B. Okab, M.A. Ayoub and A.A. Al-Haidary et al., 2011. Influence of elevated ambient temperature upon some physiological measurements of New Zealand white rabbits. Vet. Med., 4: 180-186.
Direct Link - Redwine, L., S. Snow, P. Mills and M. Irwin, 2003. Acute psychological stress: Effects on chemotaxis and cellular adhesion molecule expression. Psychosomatic Med., 65: 598-603.
Direct Link - Safaa, H.M., M.E. Emarah and N.F.A. Saleh, 2008. Seasonal effects on semen quality in Black Baladi and White New Zealand rabbit bucks. World Rabbit Sci., 16: 13-20.
CrossRefDirect Link - Sharpe, R.M., K. Donachie and I. Cooper, 1988. Re-evaluation of the intratesticular level of testosterone required for quantitative maintenance of spermatogenesis in the rat. J. Endocrinol., 117: 19-26.
CrossRefDirect Link - Toft, P., P. Svendsen, E. Tonnesen and N.J. Christensen, 1993. Redistribution of lymphocytes after major surgical stress. Acta Anaesthesiol. Scand., 37: 245-249.
CrossRefDirect Link - Koko, V., J. Djordjeviae, G. Cvijiae and V. Davidoviae, 2004. Effect of acute heat stress on rat adrenal glands: A morphological and stereological study, 2004. J. Exp. Biol., 207: 4225-4230.
PubMedDirect Link - Viswanathan, K. and F.S. Dhabhar, 2005. Stress-induced enhancement of leukocyte trafficking into sites of surgery or immune activation. Proc. Natl. Acad. Sci. USA., 102: 5808-5813.
CrossRefDirect Link - Yahaya, M.A. and P.K. Ajuogu, 2014. Respose of rabbit's testosterone and estrogen status to graded levels of white mangrove plant (Laguncularia racemosa). Int. J. Sci. Nat., 2: 196-198.
Direct Link