
M.A. Hossain
Department of Animal Science, School of Environmental and Rural Science, University of New England, NSW 2351, Armidale, Australia
A.F. Islam
Department of Animal Science, School of Environmental and Rural Science, University of New England, NSW 2351, Armidale, Australia
P.A. Iji
Department of Animal Science, School of Environmental and Rural Science, University of New England, NSW 2351, Armidale, Australia
Asian Journal of Poultry Science
Year: 2012 | Volume: 6 | Issue: 4 | Page No.: 117-128







ABSTRACT
This study was undertaken to investigate the growth performance and energy utilization of broiler chickens fed on conventional diets, based on soybean or canola, with fishmeal (SBM50 and Can50, respectively) or on Vegetable Protein (VP) diets without fishmeal (SBM75 and Can75). Feed intake was highest (p<0.001) on the SBM50 and Can50 diets and lowest on SBM75 diet. Birds in the SBM50 and Can50 diet groups were heavier (p<0.001) than the SBM75 and Can75 diet groups. Birds on SBM50 and Can50 diets achieved superior Feed Conversion Ratio (FCR), while birds on Can75 diet were the poorest. Mortality of birds was unaffected (p>0.05) between dietary groups. The Apparent Metabolizable Energy (AME) intake on the SBM50 and Can50 diets was higher (p<0.001) than in the VP. Fat intake was highest (p<0.001) on Can50 diet group while protein intake was unaffected by treatment. Heat Production (HP) was identical, but net energy of production (NEp) was improved (p<0.05) in the birds on SBM50 and Can50. Whole body energy as well as fat (p<0.05) and protein contents were also increased (p<0.01) in the SBM50 and Can50 dietary groups. Birds on SBM50 and Can50 diet groups retained higher energy as fat (REf) (p<0.05) and as protein (REp) (p<0.01). The efficiencies of utilization of ME for energy (kRE), protein (kREp) and fat (kREf) retentions were unaffected. The results demonstrated that birds on the conventional diets (SBM50; Can 50) utilized energy better and as such grew faster than the birds on VP (SBM75; Can75) diets.
PDF Abstract XML References Citation
Received: April 09, 2012;
Accepted: December 19, 2012;
Published: January 23, 2013
How to cite this article
M.A. Hossain, A.F. Islam and P.A. Iji, 2012. Energy Utilization and Performance of Broiler Chickens Raised on Diets with Vegetable Proteins or Conventional Feeds. Asian Journal of Poultry Science, 6: 117-128.
DOI: 10.3923/ajpsaj.2012.117.128
URL: https://scialert.net/abstract/?doi=ajpsaj.2012.117.128
DOI: 10.3923/ajpsaj.2012.117.128
URL: https://scialert.net/abstract/?doi=ajpsaj.2012.117.128
INTRODUCTION
Feed costs represent a major cost in poultry production (about 70%), with dietary energy sources occupying the greatest portion (70 to 75% of the diets) (Van der Klis et al., 2010). Birds tend to eat feeds mainly to satisfy their energy requirements and once this is met, they will not consume any more feeds, even if the requirements of other nutrients like protein, vitamins or minerals have not been met (Singh and Panda, 1992). For this reason, the energy contents of the diets play a pivotal role in formulating diets for poultry. Most of the dietary energy comes from plant sources in the form of starch from cereal grains. These cereal grains provide the energy component which accounts for 60 to 70% of the nutrient requirement of poultry. Protein is supplied by plant and animal sources. Plant sources are safer than animal sources but the former often lack nutrient balance. This may affect the utilization of nutrients and growth of birds on such diets. Protein sources may also supply a substantial amount of energy and their interaction with the main energy sources has a bearing on the overall energy supply and utilization. So, it is important to determine the energy value of diets containing vegetable protein sources. The performance of birds is closely associated with feed nutrients and energy utilization, which is primarily related to availability of more nutrients and energy from the feed ingredients (Olukosi et al., 2008).
In poultry research studies, Metabolizable Energy (ME) and Net Energy (NE) are usually used to measure energy availability to and utilization by the birds. Metabolizable energy can be accurately determined from the difference between the gross energy of the feed and the gross energy of excreta derived from such feeds (NRC, 1994). Metabolizable energy has been commonly accepted and extensively used to compare energy values of feedstuffs and diets for poultry and energy requirements are commonly expressed in this form. Net energy is a more accurate measure of energy utilization, as it measures the amount of gross energy that is used for productive purpose. It can be measured in a number of ways, including direct and indirect calorimetry, which are expensive. The comparative slaughter technique is cheaper and often mimics the natural rearing environment more than does calorimetry (Sakomura et al., 2003).
There are a number of constraints in vegetable protein sources, which reduce their optimal utilization, thus limiting the benefits of these feeds to broiler chickens. This results in reduced digestibility and poor performance of the birds. From the results obtained by Hossain et al. (2011) in one of their study revealed out that the nutrient digestibility and overall performance of the broiler chickens fed on vegetable protein diets were comparatively poorer than those of birds fed on conventional diets.
The current study was designed to provide answers to these differences. Apart from direct measurements of energy intake and retention, the deposition of fat and protein were assessed. This information is required to understand the partitioning of retained energy in the body between fat and protein and the utilization of ME intake in broilers for this purpose (Lopez and Leeson, 2005). The objectives of the present study were to determine the energy intake and utilization from vegetable protein diets; evaluate protein and fat retention and assess the efficiency with which fat and protein are used for depositing energy.
MATERIALS AND METHODS
A total of 204 day-old Cobb-500 broiler male chicks (46.65±0.21 g) were used in this study. At one day old, a total of 192 chicks were allocated in a Completely Randomized Design (CRD) into four dietary treatment groups (SBM75, Can75, SBM50 and Can50) with 48 chicks in each group. Each treatment had six replicates, with 8 birds per replicate cage. The remaining 12 birds were used for the initial slaughter group to provide basal compositional data. The second group of 192 birds was slaughtered at 21 days, to enable measurement of nutrient retention. The study was conducted in the University of New England (UNE), Australia, during May-June, 2011.
A total of 24 brooder cages were set up on the floor in two climate-controlled rooms (12 cages in each room); and after 17 days, four birds from every pen were transferred to metabolic cages in order to collect excreta samples for rest of the trial period. The birds were brooded at 33°C for the first two days and then the temperature was reduced gradually to 24°C at 19 days of age and maintained at this level to the end of the trial. Eighteen hours of lighting and six hours darkness per day were provided throughout the trial period except for first week only and at this period chicks were exposed to continuous lighting (23 h light:1 h darkness) program. Feed in pellet form and water were provided ad libitum. The birds were on a starter diet for the duration of the trial. Body weight and feed intake data of the birds were recorded weekly. Mortality was recorded as it occurred. Feed conversion ratio was calculated weekly and corrected for mortality.
Dietary treatments: Four experimental diets, identified as SBM75, Can75, SBM50 and Can50 were formulated with maize, wheat and vegetable oil as main energy sources, along with soybean meal, canola meal and fish meal as protein sources and later pelleted to reduce segregation (Table 1). Diets SBM75 and Can75 were vegetable protein diets formulated entirely with ingredients of plant origin to meet or exceed NRC (1994) recommendations. The two vegetable protein diets (SBM75 and Can75) were formulated with a combination of soybean meal and canola meal at a ratio of 3:1. In contrast, diets SBM50 and Can50 were conventional diets formulated with ingredients of both animal and plant origin.
| Table 1: | Composition of starter diets fed during the trial period |
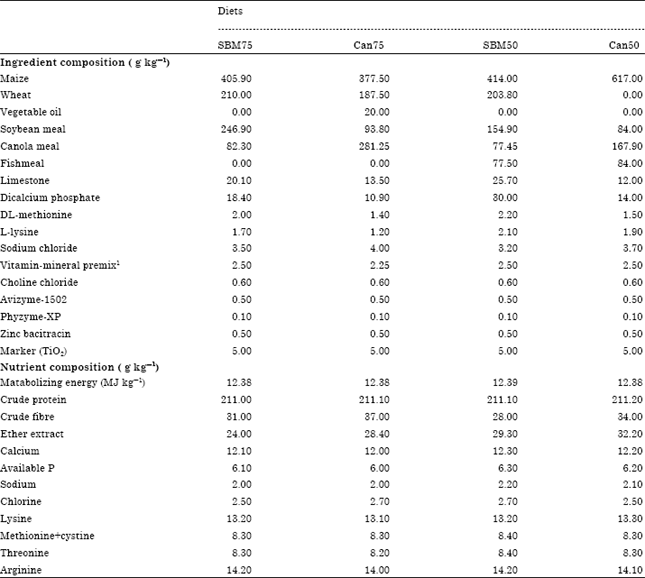 | |
| 1Provided per kg of diet (mg): Vitamin A (as all-trans retinol): 3.6 mg, Cholecalciferol: 0.09 mg, Vitamin E (as d-α-tocopherol): 44.7 mg, Vitamin K3: 2 mg: Thiamine: 2 mg, Riboflavin: 6 mg, Pyridoxine hydrochloride: 5 mg, Vitamin B12: 0.2 mg, Biotin: 0.1 mg, Niacin: 50 mg, D-calcium pantothenate: 12 mg, Folic acid: 2 mg, Mn: 80 mg, Fe: 60 mg: Cu: 8 mg: 1 mg, Co: 0.3 mg, Mo: 1 mg | |
These two diets (SBM50 and Can50) contained soybean, canola and fish meals as protein sources at a ratio of 2:1:1 one to another as shown belo (Table 1). All experimental diets were iso-caloric and iso-nitrogenous and supplemented with exogenous enzymes (Avizyme 0.5 g and Phyzyme, 0.1 g per kg of each diet) as well as Zinc Bacitracin (0.5 g kg-1). Each diet was incorporated with titanium dioxide (TiO2) at a rate of 5 g kg-1 as an indigestible marker to enable measurement of nutrient digestibility. On day 19, 20 and 21, excreta samples were collected from each cage and pooled. The excreta were immediately frozen before being dried (80°C for 8 h) in an oven to a constant weight. Excreta were pooled within each pen and ground prior to analyses. At the end of the trial, two birds per replicate cage were killed by cervical dislocation and processed as described below. All animal cares, handling and management procedures were approved by the University of New England (UNE) Animal Welfare and Ethics Committee (Approval No: AEC10/076).
Chemical analysis: The starter diets were analysed for nutrient composition. Excreta and diet samples were analysed for gross energy in order to determine the ME. Samples were dried at 105°C in a drying oven (Precision Scientific Co., Chicago, IL, USA) for 24 h for DM determination. Gross energy was determined in a bomb calorimeter (Parr 1261; Parr Instruments Co., Moline, IL, USA) using benzoic acid as a calibration standard. Titanium dioxide (TiO2) concentrations in the diets and excreta samples were measured after ashing the samples and treating the ash with boiling 7.4 M sulphuric acid according to the method of Short et al. (1996). At 21 days, two birds per replicate were killed and the whole intact carcasses were frozen immediately and later processed. Both chicks from the same cage were processed together. After chopping and coarse-grinding individual chickens, they were thoroughly mixed and two subsamples (approximately 200 g each, wet weight) were taken, finely ground and freeze-dried as described by Olukosi et al. (2008). The two subsamples were mixed together after drying and ground again. Hence chemical analysis was on one sample from each cage and not from individual chickens. The ground carcass samples were analysed for Gross Energy (GE), diethyl ether extractable fat (EE) and Nitrogen (N).
Nitrogen content of the diets and meat samples was determined according to the Dumas combustion technique as described by Sweeney (1989) using a Leco® FP-2000 automatic nitrogen analyser (Leco FP analyser model 602600; Leco Corp., St Joseph, Michigan, USA) with EDTA as a calibration standard. Crude Protein (CP) equivalent of the ingredients was calculated as N (%)x6.25. The EE was determined indirectly by the Soxhlet method for fat extraction. Around 6-8 g of finely ground sample was weighed into pre-weighed filter paper (No. 1 Whatman 185 mm) and extracted for 48 to 50 h with chloroform using a Soxhlet apparatus. After that, the samples were allowed to drain and dry at 80°C for 72 h. The EE was calculated as loss in weight and expressed as a proportion of dried sample weight. To calculate the nutrient retention and other variables the following formulae were used:
where, GEi is gross energy (MJ kg-1) in feed, GEo is the gross energy (MJ kg-1) in excreta, Ti is the concentration of titanium dioxide in the diets and To is the concentration of titanium dioxide in the excreta.
Net energy of production (NEp) was calculated as follows:
| (1) |
| (2) |
Heat Production (HP), which consists of the heat increment of feeding and fasting HP was calculated as the difference between NEp and ME intake:
where, ME intake (MEI) was calculated using the following formula:
Energy retained as fat (REf) and as protein (REp) were calculated as follows:
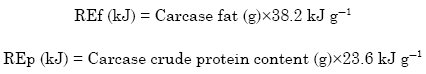 |
The values 38.2 and 23.6 kJ g-1 are energy values per gram of fat and protein, respectively, as derived by Larbier and Leclercq (1992):
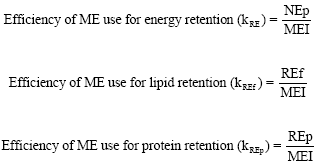 |
Statistical analysis: Statistical analyses were performed using minitab software (minitab version 15, Minitab, 2000). The data were analyzed using one-way ANOVA with diet as factor. The significance of difference between means was determined by Fisher’s least significant difference at p≤0.05.
RESULTS
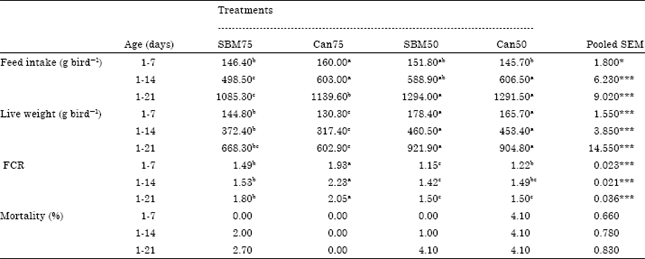
Growth performance: The growth performance of birds fed on the different diets demonstrated in Table 2. Feed intake up to 14 and 21 days was highest (p<0.001) for the Can50 diet and SBM50 diet groups, respectively, with SBM75 diet group being the lowest in feed consumption. Birds fed the SBM50 and Can50 diets were significantly heavier (p<0.001) at 7, 14 and 21 day than the birds on the VP diets, of which Can75 group birds weighed the least.
| Table 2: | Gross response of broilers fed vegetable protein and conventional diets |
 | |
| Values are mean of eight replicate groups consisting of eight birds per replicate during 1-18 days and four birds during 19 to 21 days, Means bearing uncommon superscripts within a row are significantly different at *p<0.05 and ***p<0.001 | |
| Table 3: | Metabolizable energy (ME) of diets, intake of gross energy (GEI), ME intake (MEI) and protein and fat of different diets fed to broilers |
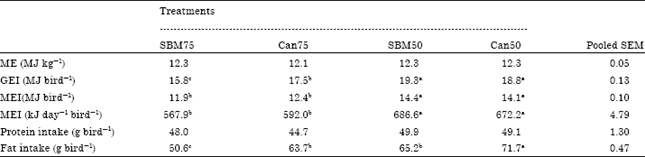 | |
| Values are mean of six replicate cages with four broilers per replicate cage at 21 days, Means bearing uncommon superscripts within a row are significantly different at p<0.001 | |
Over the entire test period, birds of Can75 diet group were the poorest (p<0.001) in Feed Conversion Ratio (FCR), while the FCR of SBM50 and Can50 diet groups were the best. The birds of SBM75 diet group were better in feed conversion efficiency (lower FCR) than the Can75 diet group throughout the experimental period. The FCR of the SBM50 diet group was similar to the FCR of Can50 dietary group during 1-14 days and 1-21 days. The mortality (%) of the chicks to 21 day was 2.7; 0; 4.1 and 4.1% on SBM75, Can75, SBM50 and Can50 diets, respectively, but did not differ significantly (p = 0.27), as shown in Table 2.
Energy utilization of birds fed on different diets: Results of energy utilization of broiler chickens fed on the two sets of diets shown in Table 3-5 and in Fig. 1, respectively. The ME content of the experimental diets was similar, with a range between 12.1 and 12.3 MJ kg-1, but Gross Energy Intake (GEI), ME intake (MEI) and fat intake were significantly higher (p<0.001) in the SBM50 and Can50 diet groups than in other diet groups (Table 3). There was no significant difference in the protein intake of the birds.
| Table 4: | Whole body energy, fat and protein contents at age of 21 day |
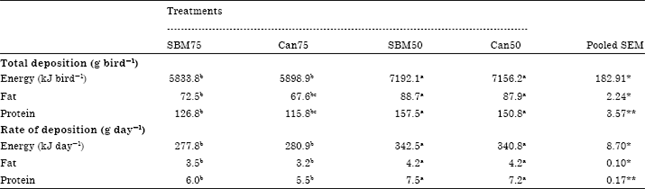 | |
| Data denote mean values of six replicate cages with four broilers per replicate cage at age of 21 day, Means bearing uncommon superscripts within a row are significantly different at *p<0.05 and **p<0.01 | |
| Table 5: | Energy retention as fat (REf) and as protein (REp) and efficiencies of energy, fat and protein use towards energy retention |
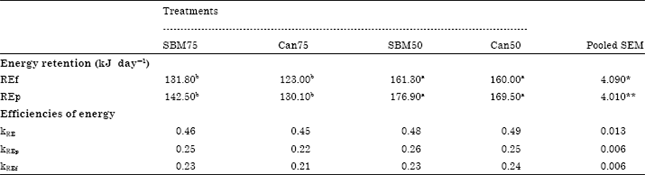 | |
| Values are mean of six replicate cages with four broilers per replicate cage at age of 21 day, Means bearing uncommon superscripts within a row are significantly different at *p<0.05 and **p<0.01 | |
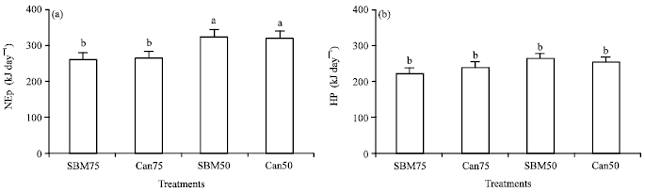 | |
| Fig. 1(a-b): | (a) Net energy for production (NEp) and (b) Heat production (HP) of broilers fed with different diets, Bars with different letter are significantly different at p<0.05 |
Birds on SBM50 and Can50 diet achieved significantly (p<0.05) higher NEp (327.9 and 326.2 kJ day-1, respectively), while birds on Can75 and SBM75 diet groups had the least NEp (263.2 and 266.3 kJ day-1, respectively) (Fig. 1). Heat production ranged from 304.7 to 358.7 kJ day-1, between the groups and was not affected by treatment.
Energy, fat and protein contents were higher (p<0.05; p<0.01) in the conventional diet groups (SBM50; Can50) than in the VP diet groups (Table 4). Energy and fat deposition was increased (p<0.05) on the SBM50 and Can50 diets and protein deposition was also elevated (p<0.01) on these two diet groups than in the VP diets. Energy, fat and protein contents of chicks on the SBM50 diet were similar to those of on the Can50 diet, but differed significantly (p<0.05; p<0.01) from the two VP dietary groups.
Chickens in the SBM50 and Can50 diets deposited energy, fat and protein at a higher (p<0.05; p<0.01) rate than those on the VP diets. Birds of the SBM75 and Can75 diet groups had similar rates of energy, fat and protein deposition. Rate of nutrients (energy, fat and protein) deposition were also similar in SBM50 and Can50 diets, but the rate of energy as well as fat (p<0.05) and protein (p<0.01) differed significantly from the other diet (SBM75; Can75) groups.
Energy retained as fat (REf) and as protein (REp) was significantly different (p<0.05; p<0.01) between the treatment groups; these being higher in the SBM50 and Can50 diet groups than in the VP diet groups (Table 5). Energy retention as protein (REp) (p<0.01) as well as energy retained as fat (REf) were higher (p<0.05) in birds raised on the SBM50 and Can50 diets than in those on the VP diets. The efficiencies in utilization of ME, protein and fat for energy retention were 0.45-0.49, 0.22-0.26 and 0.21-0.24, respectively and were generally not affected by dietary treatment.
DISCUSSION
Growth performances of birds: Growth response in broiler chickens has been regarded as the primary criterion for determining the feed nutrient requirements because the broiler chick is an ideal experimental subject with a limited nutrient store, high nutrient demand and rapid growth rate (Ammerman, 1995). In this study, overall growth performance of broilers was influenced adversely by feeding VP diets, confirming the results obtained by Hossain et al. (2011, 2012) in previous studies. The reduced performance of broilers on VP diets may be reflected by lower feed intake of the birds. The reason behind the decreased feed consumption of broilers on VP diets may be due to deficiency of essential amino acids, poor nutrient digestibility, lower palatability and anti-nutritive effects of the vegetable protein diets fed the birds. Jackson et al. (1982) reported that essential amino acid imbalances in diets reduced the biological value of the diets and decreased the feed intake. Lower palatability of the diets might be another reason for reduced feed consumption by the broiler chickens (Mahmoudnia et al., 2011). Furthermore, vegetable ingredients, particularly canola meal, contain anti-nutrients, predominantly glucosinolates, which impart bitter taste and thus affect the feed consumption of birds (Peron and Partridge, 2010). Other deleterious substances which may be present in the diets are protease inhibitors, lectins, polyphenolic compounds, saponins and non-starch polysaccharides (Hughes and Choct, 1999; Ravindran et al., 2005) can also affect the feed consumption.
The broiler chickens raised on the conventional diets demonstrated significantly improved live weight, with a superior feed conversion ratio to the VP diet groups. This trend was maintained throughout the whole trial period and became more pronounced towards the end of the experimental period. The results suggest that birds fed on conventional diets containing fish meal supported better growth and better efficiency through mainly stimulation of feed intake. The results agree with the findings of previous studies (Radhakrishnan et al., 2001; Alali et al., 2011; Hossain et al., 2011, 2012), who observed similar results with diets containing animal proteins. The improvement in growth of the birds might also result from unidentified growth factors present in fish meal (Barlow and Windsor, 1984; El Boushy and van der Poel, 1994) along with connections of known nutrients such as vitamin B12, selenium (Grastilleur, 2007), carnitine and taurine (Comb, 1998). Conversely, growth on VP diets might be impaired by the deficiency of indispensable amino acids along with some other nutrients in such diets. Previous studies have reported improvement in growth performance of broilers when they were fed VP diets supplemented with carnitine and taurine (Rabie and Szilagyi, 1998).
Energy utilization of broilers: The results of this study provide an indication of how efficiently broiler chickens utilize energy and other nutrients from VP and conventional diets. In the current study, the ME contents of the experimental diets were similar, but ME intake was reduced in groups on the VP diets. This reduction in ME intake may be due to a high fibre content and presence of anti-nutritive factors in these diets, as has been observed by some researchers (Warenham et al., 1994; Jan et al., 2008). Phytic acid, in particular can adversely affect energy utilization and the availability of other nutrients in poultry diets (Ravindran et al., 2005), although our diets were supplemented with microbial enzymes. The anti-nutritive properties of phytate can not be eliminated completely by phytase supplementation alone (Selle and Ravindran, 2007). Apart from this, increased activity of gut microflora on the dietary factors can lead to energy wastage (Choct et al., 1996) and availability and digestibility of other nutrients such as proteins and amino acids (Smits et al., 1997). Le Goff and Noblet (2001) also reported that most of the variation in digestibility of feed energy is related to the presence of dietary fibre. According to Black (2001) variation in gross chemical composition between grains may greatly affect the digestibility of feed energy. Metabolizable energy intake was high in birds on the conventional diets, possibly as a result of higher feed intake and other nutrient digestibility of these diets (Jan et al., 2008). Because fishmeal supplementation to VP diets might enhance the digestibility of amino acids of the feeds as a result of synergistic action or combination of vegetable and animal proteins in the conventional diets. As it is reported that fishmeal contains all sort of essential amino acids, particularly lysine and methionine as required to the poultry which may promote the higher uptake of feeds and improved feed efficiency of the birds as well (Solangi et al., 2002).
In this study, broilers on the conventional diets used energy better as shown by higher NEp with no significant differences in HP. This group of birds also attained heavier body weight than the VP groups. This improvement in NEp and performance of birds is evidence of more efficient utilization of energy on these diets. Olukosi et al. (2008) reported that an improvement in energy utilization may be due to improvement in nutrient and energy availability. Heat Production (HP) of birds did not differ significantly between treatments, although birds in the conventional diet groups had a numerically higher level of HP than the VP diet groups. This increase in HP on the conventional diets may be due to higher feed intake, in particular protein intake (Johnson, 2007). The conventional diets also resulted in increased whole body energy, fat and protein contents, a trend supported by Boekholt et al. (1994) who observed that when protein is not limiting in the diets of broilers, extra energy available in the diet is used for both fat and protein accretion. On the other hand, the rate of deposition of energy, fat and protein was reduced in birds on the VP diets. This could be caused by a higher fibre content, as previously reported by Uberoi et al. (1992).
In general, more energy is retained as protein than as fat, especially on the conventional diets. This also represents a more efficient use of energy and the carcass would be less fatty as a result (Hossain et al., 2012). Several researchers (Leeson and Summers, 1997) have reported that abdominal fat of birds increases with age whereas protein accretion decreases. This is simply related to the maturity of birds and is found commonly in most strains (Leeson, 1995). In the present study, the proportion of the retention of protein was found to be higher than that of fat as birds in the tested age group (0-21 day) are still in the actively growing phase of production (Bregendahl et al., 2002). The efficiency of utilization of metabolizable energy for energy, protein and fat retention was unaffected by dietary treatment, the ME being more efficiently used for energy deposition and less for protein and fat deposition. It is uncertain what the implications of this trend means, but it may explain the increase in fat deposition as birds become older.
CONCLUSION
The present study showed that conventional diets supported higher body growth and better FCR not only through higher feed intake but also through a more efficient utilization of dietary energy. The latter is supported by higher ME intake and higher NEp. Better growth response of birds on conventional diets is an outcome of improved feed efficiency plus efficient energy utilization of the broiler chickens. There is a need for the economic analysis of these sets of diets in a future study.
ACKNOWLEDGMENTS
The project was funded by the UNE, Australia and enzyme were supplied by Danisco Animal Nutrition, UK.
REFERENCES
- Alali, W.Q., C.L. Hofacre, G.F. Mathis and A.B. Batal, 2011. Effect of plant-based protein meal use in poultry feed on colonization and shedding of Salmonella Heidlberg in broiler birds. Agric. Food Anal. Bacteriol., 1: 45-53.
Direct Link - Boekholt, H.A., P. Van der Grinten, V.V.A.M. Scheurs, M.J.N. Los and C.P. Leffering, 1994. Effect of dietary energy restriction on retention of protein, fat and energy in broiler chickens. Br. Poult. Sci., 35: 603-614.
CrossRefDirect Link - Bregendahl, K., J.L. Sell and D.R. Zimmerman, 2002. Effect of low-protein diets on growth performance and body composition of broiler chicks. Poult. Sci., 81: 1156-1167.
Direct Link - Black, J.L., 2001. Variation in nutritional value of cereal grains across livestock species. Proc. Aust. Poult. Sci. Symp., 13: 22-29.
Direct Link - Choct, M., R.J. Hughes, J. Wang, M.R. Bedford, A.J. Morgan and G. Annison, 1996. Increased small intestinal fermentation is partly responsible for the anti-nutritive activity of non-starch polysaccharides in chickens. Br. Poult. Sci., 37: 609-621.
CrossRefPubMedDirect Link - Hughes, R.J. and M. Choct, 1999. Chemical and physical characteristics of grains related to variability in energy and amino acid availability in poultry. Aust. J. Agric. Res., 50: 689-702.
Direct Link - Johnson, D.E., 2007. Contributions of animal nutrition research to nutritional principles: Energetic. J. Nutr., 137: 698-701.
Direct Link - Jan, B., L. Olga and A. Romana, 2008. Energy utilization and nutritive value of maize grain cultivars for Broiler chickens. J. Central. Europ. Agric., 9: 209-211.
Direct Link - Jackson, S., J.D. Summers and S. Leeson, 1982. Effect of dietary protein and energy on broiler performance and production costs. Poult. Sci., 61: 2232-2240.
PubMedDirect Link - Lopez, G. and S. Leeson, 2005. Utilization of metabolizable energy by young broilers and birds of intermediate growth rate. Poult. Sci., 84: 1069-1076.
CrossRefDirect Link - Le Goff, G. and J. Noblet, 2001. Comparative digestibility of dietary energy and nutrients in growing pigs and adult sows. J. Anim. Sci., 79: 2418-2427.
Direct Link - NRC., 1994. Nutrient Requirements of Poultry. 9th Edn., National Academy Press, Washington, DC., USA.
Direct Link - Olukosi, O.A., A.J. Cowieson and O. Adeola, 2008. Energy utilization and growth performance of broilers receiving diets supplemented with enzymes containing carbohydrase or phytase activity individually or in combination. Br. J. Nutr., 99: 682-690.
PubMed - Radhakrishnan, L., R. Kadirvel and R. Vijayakumar, 2001. Replacement of fishmeal with vegetable protein sources in broiler diets. Indian J. Poult. Sci., 36: 300-304.
Direct Link - Rabie, M.H. and M. Szilagyi, 1998. Effects of L-carnitine supplementation of diets differing in energy levels on performance, abdominal fat content and yield and composition of edible meat of broilers. Br. J. Nutr., 80: 391-400.
Direct Link - Ravindran, V., J.S. Sands, M. Hruby and G.G. Partridge, 2005. Influence of phyzyme tm XP phytase on nutrient utilization in broilers fed diets containing graded levels of phytate. Proceedings of the 17th Australian Poultry Science Symposium, February, 7-9 2005, Sydney, New South Wales, Australia, pp: 238-241.
- Sakomura, N.K., R. Silva, H.P. Couto, C. Coon and C.R. Pacheco, 2003. Modeling metabolizable energy utilization in broiler breeder pullets. Poult. Sci., 82: 419-427.
Direct Link - Selle, P.H. and V. Ravindran, 2007. Microbial phytase in poultry nutrition. Anim. Feed. Sci. Technol., 135: 1-41.
CrossRefDirect Link - Smits, C.H.N., A. Veldman, M.W.A. Verstegen and A.C. Beynen, 1997. Dietary carboxymethylcellulose with high instead of low viscosity reduces macronutrient digestion in broiler chickens. J. Nutr., 127: 483-487.
Direct Link - Short, F.J., P. Gorten, J. Wiseman and K.N. Boorman, 1996. Determination of titanium dioxide added as an inert marker in chicken digestibility studies. Anim. Feed Sci. Technol., 59: 215-221.
CrossRefDirect Link
sajedul korim Reply
i find it very instructive and informative for poultry nutrition , specially poultry science. very good piece of article
Hossain
thanks a lot for your nice comments
A. Hasan Reply
very nice article for poultry enterprise
MAHossain
Thanks for your remarks.