
Yasser Abd El-Galil Ahmed
Department of Histology, Faculty of Veterinary Medicine, South Valley University, Qena, Egypt
Gamal Kamel
Department of Anatomy and Histology, Faculty of Veterinary Medicine, Assiut University, Assiut, Egypt
Ahmed Abo El-Magd Ahmad
Department of Anatomy and Histology, Faculty of Veterinary Medicine, Assiut University, Assiut, Egypt
Asian Journal of Poultry Science
Year: 2011 | Volume: 5 | Issue: 2 | Page No.: 56-67







ABSTRACT
The current study aimed to maximize the information on histology of Japanese quail digestive tract. Specimens from proventriculus and gizzard of post-hatching Japanese quail were embedded in paraffin and processed for microscopic examination. Grimelius silver impregnation technique was applied to localize the argyrophil endocrine cells. Quail stomach wall was presented by mucosa, submucosa, muscularis and serosa. Mucosa of the proventriculus formed plicae and sulci, which were lined by Periodic Acid Schiff reagent (PAS) and alcian blue positive simple columnar epithelium. The lamina propria connective tissue contained lymphocytic infiltration and lymphatic aggregates. Compound tubuloalveolar glandular lobules were located within muscularis mucosa. Each lobule consisted of uncoiled tubules radiating from a central cavity into which they discharge. The tubules contained negatively stained PAS and alcian blue secretory oxyntico-peptic cells. Many lobules opened together upon raised papillae into the lumen of the proventriculus via common large duct. The gizzard mucosa is covered by PAS, alcian blue and trichrome positive keratinized layer and formed of folds lead to tubular glands in the lamina propria. The gizzard glands were lined mostly with cuboidal cells. No muscularis mucosa was seen in the gizzard. The transitional areas between the proventriculus and the gizzard and between the gizzard and the duodenum had features similar to both portions before and after junctions. Argyrophil endocrine cells were abundant in the proventriculus and absent in the gizzard. Some morphometrical and histo-morphometrical growth kinetics during different stages of development were reported. These observations should be considered when studying quail digestion physiology.
PDF Abstract XML References Citation
Received: March 26, 2011;
Accepted: July 06, 2011;
Published: July 26, 2011
How to cite this article
Yasser Abd El-Galil Ahmed, Gamal Kamel and Ahmed Abo El-Magd Ahmad, 2011. Histomorphological Studies on the Stomach of the Japanese Quail. Asian Journal of Poultry Science, 5: 56-67.
DOI: 10.3923/ajpsaj.2011.56.67
URL: https://scialert.net/abstract/?doi=ajpsaj.2011.56.67
DOI: 10.3923/ajpsaj.2011.56.67
URL: https://scialert.net/abstract/?doi=ajpsaj.2011.56.67
INTRODUCTION
The avian stomach is formed of two distinct parts; the glandular portion, gastric proventriculus or true stomach (ventricular glandularis) which is caudal to the esophagus and the muscular portion, gastric ventriculus or gizzard (ventriculus muscularis) which is located caudal to the proventriculus. The two parts of the stomach are connected together by intermediate zone. The proventriculus secretes HCL and pepsin which is needed for protein digestion. The gizzard is much larger and more muscular than proventriculus and it is important for grinding the food. This grinding action of the gizzard prepares the food for further digestion in the intestine, where the first part of the small intestine, duodenum, is connected with the gizzard at gizzard- duodenal junction (Maya and Lucy, 2000). A third portion of avian stomach, called pyloric stomach, has also been described in aquatic birds and it usually contains feathers and bones because it is believed to act as a filter to prevent passing of these things to the intestine (Duke, 1997).
The proventriculus varies in size between different species. In graminivorous, it is of smaller size than carnivores where the proventriculus is quite larger and more distensible (Degen et al., 1994). The wall organization of the proventriculus of chickens, the most avian model studied by researchers, is according to the general pattern that specified most of the digestive organs; tunica mucosa, tunica submucosa, tunica muscularis and tunica serosa (Selvan et al., 2008). The proventricular folded mucosal surface is not smooth but it is covered by several projections or papillae. The proventricular glands which form the most thickness of the proventricular wall, open at the apex of these papilla by ducts elaborating pepsinogen, hydrochloric acid and mucus discharge into the stomach lumen (Liman et al., 2010). The surface lining epithelium of the proventriculus is of a simple columnar type (Rahman et al., 2003) however the glandular epithelium is formed by only one principle exocrine cells of a simple cuboidal type; oxyntico-peptic cells and some gastric argyrophil endocrine cells (Langlois, 2003).
The gizzard wall also have the four layers; mucosa, submucosa, muscular layer and serosa. One of the characteristic features of the gizzard is the presence of an internal lining or koilin membrane, a secretory layer from the gizzard glands, above the tunica mucosa, known as the cuticle. The gizzard mucosa shows low folds (plicae ventricularis) lined by simple columnar to cuboidal epithelium. Gizzard tubular glands are lined by cuboidal epithelium which is lower at the base and higher at the upper portion of the glands (Rossi et al., 2005). Muscular layer contains two pairs of smooth muscle fibers termed by thin and thick muscle pairs. The avian stomach is currently used as an indicator for a bird performance in many nutritional experiments (Aderolu et al., 2007; Esuga et al., 2008; Iheukwumere et al., 2008).
Although, the histological structure of the stomach in different avian species has widely been studied by many authors (Bezuidenhout and van Aswegen, 1990; Gabella, 1985; Imai et al., 1991) the available literature on the histology of the stomach of post-hatching Japanese quail are scanty. Therefore the aim of the current study was to give more information on the histomorphological structure of the Japanese quail stomach from one day until 45 days after hatching.
MATERIALS AND METHODS
Birds and feeding: Fifty male Japanese quail were raised for this study in Elgazeria poultry farm in Qena in the May 2002. The birds after hatching were feed on turkey starter 28% protein; soybean meal sorghum was the added to the ration as an external source of protein. After 14 days, the ration was poultry starter with 21% protein until the end of the experiment. Minerals, vitamins and antibiotic were added to the ration. Temperature was 38°C during the first week, but decreased to 24-30°C to the end of the experiment. Regarding the light program, the birds after hatching till the first 3 days were exposed to 24 h light/day and decreased gradually to reach 12 h/day after 14 days and to the end of the experiment.
Sample processing for light microscopy: Five apparently healthy birds at different ages (1, 15, 30, 45 days) at least were collected and body weight was recorded. The birds were anesthetized with ethyl ether inhalation. The birds were perfused via heart with 10% buffered neutral formalin or Boin's fluid. Entire gastrointestinal tract was dissected and small pieces from the proventriculus, proventricular-gizzard junction, gizzard and gizzard-duodenal junction were quickly immersed in fixative for 1-3 days. The specimens were dehydrated in ascending grades of ethanol, cleared in methyl benzoate, embedded in paraffin wax and step serial sections (5-7 micron thickness) were cut using sledging microtome. Slides were stained with hematoxylin and eosin for general examination, Masson's trichrome for detection of connective tissue, Periodic Acid Schiff reagents (PAS) for identification of acidic mucous, alcian blue for identification of neutral mucous contents and Grimelius’ silver nitrate impregnation technique for demonstration of argyrophil endocrine cells.
Morphometrical and histo-morphometrical analysis: Different gross and histological parameters of the growth rate kinetics of the stomach of 1-day to 45-day post-hatching mail Japanese quail were estimated. The gross parameters included the stomach: body weight ratio, stomach length, proventricular length and diameter and gizzard diameter and thickness. Histological growth parameter included proventricular mucosal fold length, proventricular surface epithelium height, proventricular glandular epithelium height, gizzard keratinized layer thickness, gizzard gland thickness and gizzard glandular epithelium height. In addition, the total count of the argyrophil endocrine cells/ section in different parts of the samples examined was investigated.
Statistical analysis: Results were presented as Mean±SE (n≥3). Statistical differences between groups were evaluated using a one way ANOVA; p-values<0.05 were considered significant.
RESULTS
Histology of the stomach of the post-hatching male Japanese quail
Proventriculus: The proventriculus of post-hatching quail was spindle shaped organ located between the esophagus and gizzard. The proventricular wall had all the four tunics of digestive organs; tunica mucosa, tunica submucosa, tunica muscularis and outermost tunica serosa (Fig. 1a). The surface mucosa of the proventriculus was invaginated at regular intervals forming mucosal plicae and sulci (Fig. 1b). The surface lining epithelium was simple columnar with vesicular nucleus located near to the basement membrane and foamy acidophilic cytoplasm. The cells showed PAS- and alcian blue positive mucin granules occupying the supra nuclear area of the cells (Fig. 1b and c). Under the lamina epithelialis, the lamina propria was formed of loose connective tissue with lymphocytic infiltration and lymphatic nodules of different sizes were frequently demonstrated; especially close to the mucosal papillae (Fig. 1a). Compound tubuloalveolar proventricular glands formed the greatest thickness of the proventricular wall (Fig. 1a). The glands composed of rounded, oval, hexagonal or polymorphic lobules separated from each other by thin perilobular connective tissue sheath containing fibroblasts and few smooth muscle fibers (Fig. 1a). The wall of each lobule was formed of numerous secretory alveoli or tubules open together into a wide central cavity, from which a wide duct originates (Fig. 1d). Ducts from several lobules joined together to form a short main duct which connected to the apex of the raised mucosal papillae and open onto the narrow lumen of the proventriculus (Fig. 1c). The duct system of the proventricular glands was lined with tall columnar epithelium with oval or vesicular nuclei which located sometimes at different levels giving the epithelium pseudostratified appearance. The duct lining epithelium was faintly stained with PAS and alcian blue (Fig. 1c). The secretory cells, oxyntico-peptic cells, of the glandular tubules or alveoli at one day were basophilic and varied in size and shape (Fig. 1d, e). They were cuboidal, columnar, triangular, polyhedral or inverted flask in outline (Fig. 1e). Their nuclei were nearly rounded and located near to the basement membrane. The secretory cells oriented obliquely to the long axis of the glandular tubules of the proventriculus and were separated from each other by relatively narrow spaces giving the epithelial cells a serrated appearance (Fig. 1e). The secretory cells were negatively stained with PAS and alcian blue techniques (Fig. 3b).
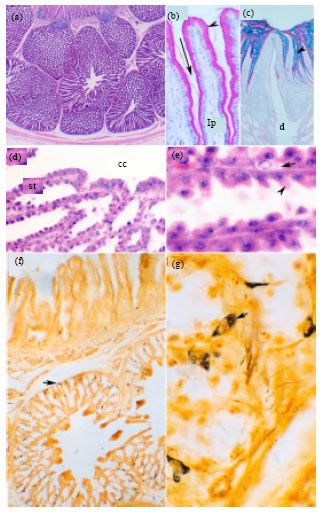 | |
| Fig. 1 (a-g): | Histology of the Quail proventriculus, Paraffin sections from the proventricular wall of a 45- (a, c, d, e), 30- (b) and 1- (f, g) day post-hatching quail. Sections stained with H and E (a, d, e), PAS (b), PAS-alcian blue (c) and Grimelius silver nitrate impregnation (g, h). Light micrographs showing the outline structure of the proventricular wall (a), PAS positive cells in the mucosal folds (b), PAS- alcian blue positive materials of the proventricular surface epithelium (c), glandular epithelium (d, e) and argyrophil endocrine cells (g, h). In a, arrow indicates lymphoid follicles underneath the epithelium, arrowhead indicating connective tissue separating proventricular glands (pg), which containing central cavity (cc). The glands located within the muscularis mucosa (mm). In b, mucosal folds formed of plicae (arrowhead) and sulci (arrow) with PAS-positive cells underneath by connective tissue lamina propria (lp). In c, note PAS-alcian blue positive cells and large duct opening at the proventricular lumen. In d, note the secretory tubules with serrated appearance (st) and central cavity of the proventricular glands (cc). In e, arrowhead indicates glandular epithelium and arrow indicates endocrine cells of the proventricular secretory tubules. In g and h, arrows indicate the distribution of argyrophil endocrine cells (a) and details of these cells (h). Magnifications are x25 (a), x400 (b, d, h), x100 (c, g), x1000 (e) |
In h and e stained paraffin sections, the endocrine cells were lightly stained and located between the basement membrane and the secretory cells of the proventricular glands (Fig. 1e). Grimelius silver nitrate impregnation of Bouin's fixed paraffin sections demonstrated abundant argyrophil endocrine cells, located singly between the lamina epithelialis and between the glandular epithelium and its basement membrane. They were more abundant at the peripheral part of the glands than the central part (Fig. 1f). The argyrophil endocrine cells were rounded, oval, triangular or branched cells with few cytoplasmic processes and sometimes appeared elongated in shape (Fig. 1g). Their cytoplasm demonstrated few argyrophilic brown to brown-black granules located at the peripheral part of the cell and the nucleus was rounded and eccentric. The cytoplasmic nuclear ratio was relatively small. The muscularis mucosa of the proventriculus was formed of two small muscle fibers layers; inner isolated muscular bands arranged in a longitudinal manner and outer band arranged in a circular manner. The proventricular glands located between the inner and outer layers of the muscularis mucosa. The submucosa was poorly developed and represented by a narrow layer of loose connective tissue containing blood vessels. The muscular layer was formed of thick inner circular and thin outer longitudinal smooth muscle fibers. The proventriculus was covered from outside by serous peritoneal membrane which was formed of a single layer of flattened mesothelial cells rested on a submesothelial connective tissue layer.
Gizzard: The gizzard was small spheroid organ located caudal to the proventriculus. The gizzard wall was represented by mucosa containing branched tubular glands, submucosa, muscular layer and serosa (Fig. 2a). The surface mucosa showed clear ridges and depressions (Fig. 2a). The gizzard surface lining epithelium was of simple columnar or cuboidal type with rounded to oval basal nucleus and basophilic cytoplasm (Fig. 2b). The surface epithelium showed sequences of degenerative changes with sloughing the dead cells (Fig. 2c). The mucosal epithelium was PAS and alcian blue positive especially at the apical portion of the cells. The mucosa was covered with a thick keratinized laminated layer formed by the secretion of the glandular tubules and the degenerated surface epithelial cells which react positively to the acid fuchsin after using trichome stain. Branched tubular glands was located in the lamina propria and opened in the mucosal crypts by short neck (Fig. 2a-d). The glands located within the mucosal ridges were less branched and had a straight regular appearance while those locating in the depressions of the mucosal surfaces were more branched. The lumina of the tubular glands were narrow and filled with homogenous lightly stained acidophilic materials with h and e (Fig. 2b-d). The secretory material within the lumen of the glandular tubules showed positive reaction with PAS and acid fuchsin stains. The tubular glands were lined with low columnar or cuboidal cells with rounded vesicular basal nucleus and darkly stained basophilic cytoplasm. No muscularis mucosa was present making no partitions between lamina propria and submucosa. The muscular layer was well developed forming the main bulk of the gizzard wall and represented by smooth muscle fibers arranged mostly in a circular manner, however some fibers arranged as an outer longitudinal and inner oblique layers (Fig. 2a). Beneath the muscular layer, there was a tendinous layer of collagen bundles arranged parallel to each other as stained with trichrome method. Argyrophil endocrine cells were completely absent from the gizzard.
Proventricular-gizzard junction: The proventricular-gizzard junction was a short area between the proventriculus and the gizzard. At this area, the proventricular glands terminated abruptly (Fig. 3a). Near the junctional area, the proventricular glands demonstrated PAS- positive secretion within the duct system (Fig. 3b). The alcian blue- positive materials decreased, while the PAS-positive substances predominated the secretion covering the surface (Fig. 3b). The proventricular plicae and sulci appeared shorten.
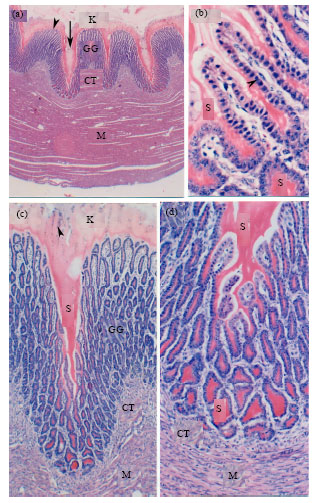 | |
| Fig. 2 (a-d): | Histology of the Quail Gizzard, Paraffin sections from the gizzard wall of a 1-day post-hatching quail stained with H and E showing the outline of the gizzard wall (a), gizzard glands (b, d), gizzard mucosa (a, c). In a, note keratinized layer (K), mucosal ridge (arrow head) and crypt (arrow), gizzard glands (GG), connective tissue lamina propria-submucosa (CT) and heavy muscular layer (M). In b, note gizzard gland acidophilic secretion (S), gizzard gland epithelium (arrowhead). In c and d note keratinized layer (K) containing degenerated cells (arrowheads), gizzard glands (GG) and connective tissue lamina propria-submucosa (CT) underneath by smooth muscle fibers (M). Magnification is 25x (a), 400x (b), 100x (c) and 200x (d) |
The connective tissue lamina propria was occupied by short branched tubular glands lined with cuboidal epithelium. The muscularis mucosa was formed of relatively thick layer of smooth muscle fibers arranged longitudinally. The muscular layer was formed of smooth muscle fibers arranged into inner thick circular layer and outer thin longitudinal layers. The muscularis mucosa and the inner layer of tunica muscularis merged with the gizzard muscle, while the outer layer disappeared near the gizzard. The serosa was similar to that of the proventriculus.
Gizzard-duodenal junction: The Gizzard-duodenal junction is the transitional zone between the gizzard and the duodenum (Fig. 3c). In this zone, Gizzard keratinized layer decreased in thickness.
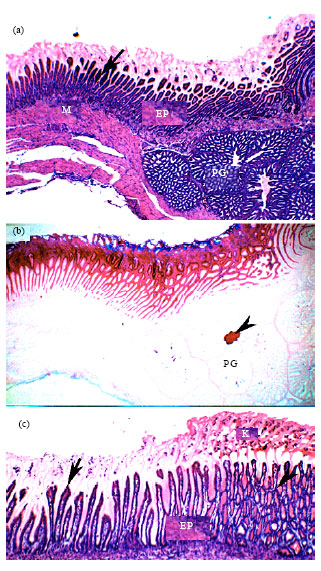 | |
| Fig. 3 (a-c): | Histology of the Quail proventricular-gizzard-duodenal junctions, Paraffin sections from the proventricular-gizzard (a, b) and gizzard-duodenal (c) junctions of a 1-day post-hatching quail stained with H and E (a, c) and PAS-alcian blue (B). In A, note the changes in the keratinized layer (arrow), the epithelial lining (EP), the Proventricular Glands (PG) and the muscular layer (M). In b, note the predominant PAS and decreased alcian blue staining of the epithelium and the keratinized layer (arrow), the PAS-positive material in PG central cavity. In C, note the transition between gizzard (arrowhead) epithelium (EP) and duodenal (arrow) and the changes in the keratinized layer (K). Magnification, is 25x (a, b) and 100x (c) |
The gizzard pits were increased in depth and the mucosa appeared with villous appearance and the mucosal glands become shorter. The glands were lined with columnar epithelium with rounded vesicular basal nucleus and lightly acidophilic cytoplasm. Lamina propria was highly infiltrated with lymphocytes and lymphocytic infiltration. Muscularis mucosa was formed of circularly arranged smooth muscle fibers. The gizzard musculature was greatly decreased in thickness.
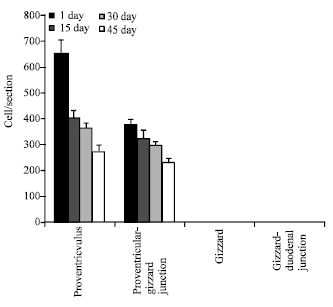 | |
| Fig. 4: | Distribution and frequency of the argyrophil endocrine cells in the quail stomach, argyrophil endocrine cells were count using paraffin sections from proventriculus, proventricular-gizzard junction, gizzard and gizzard-duodenal junction and stained with Grimelius silver impregnation. Each column represents Mean±SD. In comparison between proventriculus and other parts at each stage of development, p-value was <0.05 |
| Table 1: | Morphometrical changes in the growth rate of quail stomach |
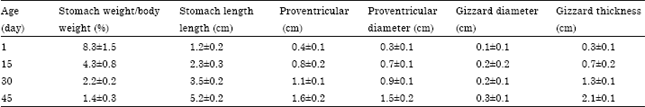 | |
| Values are as Mean±SD | |
Morphometrical analysis of the male Japanese quail stomach after hatching: Many gross growth parameters (Table 1) were measured during different ages and the results were compared between each 2 successive ages. With the advancing age, it was found that the stomach/body weight ratio showed significantly decreased, stomach length significantly increased, proventricular length significantly increased each second age except between 30 and 45 days, proventriculus diameter showed a significant increase between each 2 successive ages and gizzard diameter did not significantly increase but the gizzard length did.
Histo-morphometrical studies (Table 2) were evaluated between each 2 successive ages. Mucosal fold thickness significantly increased, height of proventricular-surface epithelium significantly increased, height of proventricular glandular epithelium significantly increased at 15 days but not after and thickness of the gizzard keratinized layer significantly increased but not at 45 days and so did the gizzard gland thickness, however, the height of the glandular epithelium showed a significant increase between each 2 successive ages.
In addition, the number of argyrophil endocrine cells/section in different specimen examined during each growth age was counted and expressed as Mean±SD (Fig. 4).
| Table 2: | Histo-morphometrical changes in the growth rate of quail stomach |
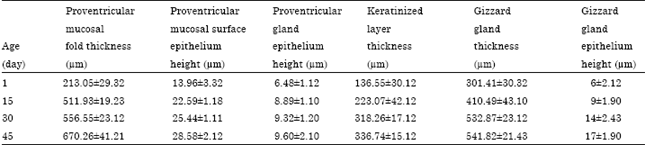 | |
| Values are as Mean±SD | |
The Number of argyrophil endocrine cells of the proventriculus of 1-day post-hatching quail was the greatest; 650±50 cells/ section, decreased to 270±250 cells/ section at 45 days post-hatching. No argyrophil endocrine cell was found in the gizzard during any stage of development.
DISCUSSION
The present study aimed to document the morphology of stomach of the post-hatching male Japanese quail. To achieve this aim, paraffin sections prepared from proventriculus, proventricular-gizzard junction, gizzard and gizzard-duodenal junction were stained with different histological stains and examined with light microscopy. It was shown that the stomach of quail was formed of two parts; proventricular and gizzard and the wall had four layers of digestive organs; mucosa, submucosa, muscularis and serosa. The proventriculus mucosa showed plicae and sulci lined by simple columnar containing PAS and alcian blue positive mucin granules (neutral and acid mucin, respectively) as reported by Hodges (1974) in domestic fowl. The presence of neutral and acid mucin acts as a barrier to protect the proventricular mucosal surface (Mogil’naia et al., 1978). In agreement with Prasad and Kakade 1990 in duck (1990) and Rahman et al. (2003) in chickens, the current study showed that lymphocytes and lymphatic nodules were seen in the lamina propria of the quail proventricular surface mucosa. That is likely, indicate a sort of participation of the proventriculus in the quail immune response. The compound tubuloalveolar proventricular glands formed the greatest thickness of the proventricular wall supporting results of others (Imai et al., 1991; Okamoto and Yamada, 1981; Prasad and Kakade, 1990). The glands located between the inner and outer layers of the muscularis mucosa. The location of the proventricular glands is a matter of controversy between the authors. Some authors reported that the proventricular glands present in the lamina propria of duck proventriculus (Calhoun, 1954) in similar to our results, however many studies suggested the glands in other species to be submucosal (Farner, 1960) contradicting our results. These differences in the location of the glands may due to species variation but may be also due to developmental stages differences. The glands consisted of lobules, each of which had numerous tubules radiating from a central cavity into which they discharge. As mentioned in an earlier work, the secretion from many central cavities connected to the raised mucosal papillae with a large duct (Imai et al., 1991). The secretory cells, oxyntico-peptic cells are separated from each other by narrow spaces giving the cells serrated appearance, due to the presence of cell junctions at the lower lateral portions of the cells but not at the upper ones. The cells stained negatively for PAS and alcian blue indicating these cells may not of a mucous secretory function, but is certainly secret HCL and pepsin analogues to that of mammalian stomach secretion (Hodges, 1974). Many argyrophil endocrine cells were seen among the osyntico-peptioc cells similar to reports from other studies (Okamoto and Yamada, 1981). One of the argyrophil endocrine cell products is the glucagon which is known as a gastric acid secretion power inhibitor peptide (Kirkegaard et al., 1982). Thus, it is not surprising that the proventriculus, especially that from one day post-hatching quail have the greatest number to protect its mucosa from the effect of HCL.
The current study revealed that the quail gizzard was relatively small spheroid organ located between the proventriculus and the small intestine. In agreement to other reports, the current study showed that the gizzard wall made by mucosa, submucosa, muscularis and serosa (Bailey et al., 1997; Gabella, 1985). The gizzard mucosa covered by a thick keratinized laminated layer of koilin membrane which is formed of proteinous material similar to keratin and stained positive for PAS and alcian blue indicating the presence of neutral and acidic mucin within its contents; in agreement with reports of other authors (Selvan et al., 2008). This layer with the strong muscular layer of the gizzard is concerning with grinding the food, but also it may protect the underlying tissues from mechanical or chemical abrasions. The mucosa had gastric pits or crypts lead to gizzard tubular glands lined by cuboidal epithelium and stained positively to both PAS and alcian blue indicating the presence of both neutral and acid mucin, similar to the proventriculus. The gizzard tubular glands located in the lamina propria and extend to the submucosa in agreement with (Akester, 1986). The glands lined mostly by chief cells which stained basophilic, similar to in fowl (Toner, 1964). These cells are suggested to secrete photolytic enzyme, pepsin. The present study revealed that no muscularis mucosa was present in quail gizzard; that may allow the tubular gizzard glands to penetrate the lamina propria more deeply and to become more active in secreting the keratinized layer.
This study showed the debarkation of the junctions of the stomach. The proventricular-gizzard junction characterized by abruptly termination of the proventricular glands, shoterining of the proventricular mucosal folds and thickened muscular layer. The gizzard duodenal junction showed different morphology from the gizzard and the duodenum. It was distinguished from the gizzard by the presence of villous like mucosa and decreased thickness of the keratinized layer and length of the gizzard glands in addition to the lining tall columnar epithelium and the presence of muscularis mucosa. On the other hand, this zone can be readily distinguished from the duodenum by the absence of duodenal villi and duodenal glands. Also, neither goblet cells nor argyrophil endocrine cells were seen in this area.
Different growth parameters in the developing post-hatched quail have been tabulated. These results should be considered as an average baseline when studying some pathological diseases affecting quail digestive system in relating to age.
The current study explained in details the morphological features of the male quail stomach. Further studies are recommended to maximize our knowledge about quail biology in the purpose of use this bird as an experimental model in studying the physiology of avian digestion in health and diseases.
REFERENCES
- Aderolu, A.Z., E.A. Iyayi and A.A. Onilude, 2007. Performance, organ relative weight, serum and haematology parameters in broiler finisher fed biodegraded brewers dried grain. Pak. J. Nutr., 6: 204-208.
CrossRefDirect Link - Akester, A.R., 1986. Structure of the glandular layer and koilin membrane in the gizzard of the adult domestic fowl (Gallus gallus domesticus). J. Anat., 147: 1-25.
PubMedDirect Link - Bailey, T.A., E.P. Mensah-Brown, J.H. Samour, J. Naldo, P. Lawrence and A. Garner, 1997. Comparative morphology of the alimentary tract and its glandular derivatives of captive bustards. J. Anat., 191: 387-398.
CrossRefDirect Link - Bezuidenhout, A.J. and G. van Aswegen, 1990. A light microscopic and immunocytochemical study of the gastrointestinal tract of the ostrich (Struthio camelus L.). Onderstepoort J. Vet. Res., 57: 37-48.
PubMedDirect Link - Rossi, J.R., S.M. Baraldi-Artoni, D. Oliveira, C. da Cruz, V.S. Franzo and A. Sagula, 2005. Morphology of glandular stomach (Ventriculus glandularis) and muscular stomach (Ventriculus muscularis) of the partrigde Rhynchotus rufescens. Ciecia Rural, 35: 1319-1324.
CrossRefDirect Link - Degen, A.A., G.E. Duke and J.K. Reynhout, 1994. Gastroduodenal motility and glandular stomach function in young ostriches. Auk, 111: 750-755.
Direct Link - Duke, G.E., 1997. Gastrointestinal physiology and nutrition in wild birds. Proc. Nutr. Soc., 56: 1049-1056.
CrossRefPubMedDirect Link - Esuga, P.M., A.A. Sekoni, J.J. Omage and G.S. Bawa, 2008. Evaluation of enzyme (maxigrain) supplementation of graded levels of palm kernel meal (PKM) on the performance of broiler chickens. Pak. J. Nutr., 7: 607-613.
CrossRefDirect Link - Gabella, G., 1985. Chicken gizzard. The muscle, the tendon and their attachment. Anat. Embryol. Berl, 171: 151-162.
PubMed - Iheukwumere, F.C., E.C. Ndubuisi, E.A. Mazi and M.U. Onyekwere, 2008. Performance, nutrient utilization and organ characteristics of broilers fed cassava leaf meal (Manihot esculenta Crantz). Pak. J. Nutr., 7: 13-16.
CrossRefDirect Link - Imai, M., T. Shibata, K. Moriguchi, M. Yamamoto and H. Hayama, 1991. Proventricular glands in fowl. Okajimas Folia Anatomica Japonica, 68: 155-160.
CrossRefPubMedDirect Link - Kirkegaard, P., A.J. Moody, J.J. Holst, F.B. Loud, P.S. Olsen and J. Christiansen, 1982. Glicentin inhibits gastric acid secretion in the rat. Nature, 297: 156-157.
CrossRef - Langlois, I., 2003. The anatomy, physiology and diseases of the avian proventriculus and ventriculus. Vet. Clin. North Am. Exot. Anim. Pract., 6: 85-111.
PubMed - Liman, N., E. Alan, Kucuk and G. Bayram, 2010. The differences between the localizations of MUC1, MUC5AC, MUC6 and osteopontin in quail proventriculus and gizzard may be a reflection of functional differences of stomach parts. J. Anat., 217: 7-66.
PubMed - Maya, S. and P. Lucy, 2000. Histology of gizzard-duodenal junction in Japanese quail (Coturnix coturnix japonica). Indian J. Poult. Sci., 35: 35-36.
Direct Link - Mogil’naia, G.M., M.G. Shubich, V.I. Dudetskii and L. Bogatyr, 1978. Comparative histochemical characteristics of the secretion of superficial gastric epitheliocytes. Arkh. Anat. Gistol. Embriol., 75: 43-51.
PubMed - Okamoto, T. and J. Yamada, 1981. Light and electron microscopic studies on the endocrine cells in the duck proventriculus. Nippon Juigaku Zasshi, 43: 863-870.
PubMed - Prasad, R.V. and K. Kakade, 1990. Histology and histochemistry of proventriculus of domestic duck (Anas platyrhynchos L.). Mysore J. Agric. Sci., 25: 506-511.
Direct Link - Rahman, M.L., M.R. Islam, M. Masuduzzaman and M.Z.I. Khan, 2003. Lymphoid tissues in the digestive tract of deshi chicken (Gallus domesticus) in Bangladesh. Pak. J. Biol. Sci., 6: 1145-1150.
CrossRefDirect Link - Selvan, P.S., S. Ushakumary and G. Ramesh, 2008. Studies on the histochemistry of the proventriculus and gizzard of post-hatch guinea fowl (Numida meleagris). Int. J. Poult. Sci., 7: 1112-1116.
CrossRefDirect Link - Toner, P.G., 1964. The fine structure of gizzard gland cells in the domestic fowl. J. Anat. Lond., 98: 77-85.
Direct Link
midhat abd elfatah elkasheif Reply
thanks u very much please i want link them to our best and over scentific services