
Mohamed Saied Ali Khalil
Department of Plant Pathology, Agriculture and Biology Research Institute, National Research Centre, Dokki, Giza, Egypt
Amany Abd El-Mohsen Ramadan
Department of Botany, Agriculture and Biology Research Institute, National Research Centre, Dokki, Giza, Egypt
LiveDNA: 20.31087
ORCID: 0000-0002-4573-2729
Sahar A. EL-Sayed
Institute of Plant Pathology, Agricultural Research Centre, Giza, Egypt
Ahmed M. El-Taher
Department of Agriculture and Botany, Faculty of Agriculture, Al-Azhar University, Cairo, Egypt
Asian Journal of Plant Sciences
Year: 2023 | Volume: 22 | Issue: 1 | Page No.: 25-36







ABSTRACT
Background and Objective: Pea (Pisum sativum L.) was attacked by many foliar diseases. Downy mildew, powdery mildew and rust are the most common diseases that affect the quantity and quality of this crop. To control these diseases, certain natural organic acids, e.g., citric, benzoic, salicylic, boric, sorbic acids and also coumarin were foliar sprayed to examine the possibility of avoiding the adverse effect of biotic stress. Materials and Methods: The experiment was conducted at Kafr-El-Dwar district, Beheira Governorate, Egypt during 2 successive summer seasons of 2019 and 2020 to evaluate pea plants foliar spray with some organic acids and coumarin at the rate of 0.25 and 0.50% against some foliar diseases. Results: The results proved that citric and benzoic acids at 0.50% induced the highest reduction of downy, powdery mildew and rust, while coumarin recorded the lowest reduction at the same concentration and other tested materials came moderately. Moreover, all tested substances gave clear increases in crop parameters and alleviated the harmful effects of biotic (diseases) stress. Acceleration in the recovery in terms of enhancing photosynthetic pigments, indole acetic acid, phenolic, some compatible solutes and antioxidant content was recorded. Also, a positive variation in anatomical structure, e.g., the leaflet blade thickness, upper and lower epidermal layers, vascular bundles, midrib zone, palisade and spongy tissues was noticed. Conclusion: Conclusively, foliar spraying with tested materials alleviated the harmful effects of biotic stress and accelerated the recovery and improved pea plants productivity.
PDF Abstract XML References Citation
Copyright: © 2023. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Mohamed Saied Ali Khalil, Amany Abd El-Mohsen Ramadan, Sahar A. EL-Sayed and Ahmed M. El-Taher, 2023. Effectiveness of Natural Antioxidants on Physiological, Anatomical Changes and Controlling Downy, Powdery Mildew and Rust Diseases in Pea Plants. Asian Journal of Plant Sciences, 22: 25-36.
DOI: 10.3923/ajps.2023.25.36
URL: https://scialert.net/abstract/?doi=ajps.2023.25.36
DOI: 10.3923/ajps.2023.25.36
URL: https://scialert.net/abstract/?doi=ajps.2023.25.36
INTRODUCTION
Pea (Pisum sativum L.) is a widely grown vegetable crop for human use. Peas are regarded a matter because of their great nutritional value, which includes vitamins, protein and carbohydrates1. Numerous diseases harm pea output quantity and quality. The most important of these diseases are downy mildew, powdery mildew and rust. Peronospora viciae, which is found in many parts of the world, causes downy mildew, one of the deadliest diseases afflicting pea plants. As a systemic disease of this pathogenic organism destroys seedlings, while local infections affect leaves and pods. Mycelia and oospores of the organism P. viciae can be found in seed coatings. On pea plants, different types of downy mildew disease symptoms can be noticed. Throughout the crop life cycle, systemic, local and pod infections can be identified as three different symptom types2.
Pea powdery mildew disease result of Erysiphe pisi is an air-borne disease of around the world conveyance. It is one critical disease that infection the pea plant. The disease can reduce the yield by 25-50%, at the same time it reduces total yield biomass, number of seeds per pod, number of pods per plant, plant height and number of nodes. The malady to influences green pea quality3, Uromyces fabae is responsible for pea rut disease, which causes partial defoliation for all the green parts of the plant including pods of susceptible varieties in warm humid regions, responsible for huge economic losses.
Pea plants under biotic and abiotic stressful conditions, signalling intermediates (Reactive Oxygen Species (ROS)) overproduced and cause oxidative stress4. These particles are known to be mindful of damage of cellular membranes throughout lipid peroxidation and capacity to destruct chlorophyll, proteins and DNA5. To cope with osmotic stress, plants accumulate proline, Glycine Betaine (GB), soluble proteins and soluble carbohydrates4. The molecular basis of non-host resistance presumably inducible responses of a large array of proteins and other organic molecules produced which represents constitutive barriers before infection or during pathogen attack6,7. Constitutive defences tools are morphological and cell walls, structural barriers, e.g., trichomes, thorns epidermis layer, etc., chemical compounds, e.g., phenolics, metabolites, nitrogen compounds, etc., enzymes and proteins8,9.
Not only by defending the plant against invasion but also by strengthening plant strength and rigidity, which confers tolerance or resistance to biotic stress. Chemical resistance inducers are commonly utilised as bioactive agents to combat soil-borne and foliar plant diseases10. In this concern, phenolic compounds as a chemical inducer (vanillic, garlic, salicylic, cinnamic, P-coumaric, benzoic acids and coumarin) are considered protecting particles facing biotic and abiotic stress11,12. Many plant phenolic compounds have antibacterial properties, act as signal molecules or function as precursors to lignin as structural polymers13. The degree of plant resistance to diseases was proportional to the rise in phenolic content14. Chemical inducers encourage specific enzymes catalyzing biosynthetic responses to form resistance compounds such as polyphenols, pathogenesis-related proteins that enhance the resistance of the plant to pathogen15,16.
Salicylic acid could be a phenolic endogenous plant development bio controller that works as an antioxidant compound that contributes to the administration of physiological processes which is considered one of the defence mechanisms against biotic stress17. Also, foliar spray with salicylic acid swelled thickness of both midvein and lamina of flyers of Egyptian lupine ‘Giza 2’ due to extend initiated in thickness of palisade and light tissues to extend initiated measure in midvein bundle18. The valuable effects of salicylic acid on plant growth and productivity of cowpea plants are well-recorded19.
Benzoic acid is known to provide plants with abiotic stress tolerance20. Abdallah et al.21 reported that benzoic acid (carboxyl) is a natural antioxidant synthesized by plants and diffuses in the rhizosphere area to enable the assimilation of mineral nutrients. The increments in nutrient uptake helped plants in sandy soil to resist the nutrients shortage adverse effects and consequently improved yield quantity and quality.
Citric acid is a regular component in many vase solution preparations that regulate the pH that reduce bacterial creation and improve the water conductance in cut flowers xylem22. Citrate and malate are intermediate organic acids in the Krebs cycle which is participating in oxidative phosphorylation that produces cellular energy23.
Boric acid is a feebly acidic hydrate of boric oxide with gentle clean, antifungal and antiviral properties. Boron is a fundamental plant micronutrient taken up through the roots generally within the frame of boric acid24.
Sorbic acid may be a common natural compound (unsaturated greasy acid) that has antimicrobial movement against moulds, yeasts and organisms25.
The objective of this work was to evaluate the protective role of natural organic compound to biotic stress through increment in biochemical constituents and improvement anatomy measurements and consequently the productivity of pea plants.
MATERIALS AND METHODS
Field trials: Throughout cultivation season (2019/2020) field trials were carried out at Kafr-El-Dwar district, Beheira Governorate, Egypt to evaluate foliar spray with some organic acids and coumarin against some foliar diseases on pea plants, i.e., downy mildew, powdery mildew and rust. The forecasting weather throughout the experimental periods from November, 2019 until March, 2020 (C.F. https://www. timeanddate.com/weather/Egypt/Cairo/historic?month=12&year=2019) was recorded.
Field-tested materials: Pea seeds cv. Master B were get from Vegetables Crop Research Department, Horticultural Research Institute, Agricultural Research Center, Egypt. Organic acids, i.e., salicylic, sorbic, citric, boric and benzoic acids in addition to coumarin were obtained from Al-Gomhoria Company Ltd., for chemicals and medicinal instruments, Cairo, Egypt.
Field experiments: Evaluation of the suppressing effectiveness for foliar spraying pea with organic acids, i.e., citric, benzoic, salicylic, sorbic and boric acids in addition to coumarin at 0.25 and 0.50% concentrations against diseases powdery mildew, downy mildew and rust were performed under natural infection conditions in the field. At the growing season both diseases, powdery mildew and downy mildew started to appear in December and were intensively recorded in January, while, rust disease started to appear and was recorded at the end of February.
The field was divided into plots 3×3 m2 and each plot consisted of five rows with 15 hills/row on the eastern side. Pea Master B cultivar seeds were sown as two seed/hill, two edges of each hill 20 cm between holes. The seeds were sown on 15 November, 2019. The organic acids and coumarin mentioned before were applied twice at concentrations of 0.25 and 0.50%, where the first one was applied at 30 days after sowing and the second one was at 30 days after the first application. The experimental design was a completely randomized block design with 3 replicates for each treatment. The disease severity was calculated 70, 80 and 90 days after sowing for above mentioned studied diseases and it was determined under natural field conditions based on a score chart of 0-5 (0-no infection, 1-1-10, 2-10.1-15, 3-15.1-25, 4-25.1-50 and 5->50% according to Elgamal and Khalil26 using the following equation:
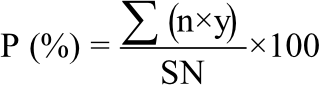 |
| P | = | Disease severity |
| n | = | Number of infected leaves in each category |
| y | = | Numerical values of each category |
| S | = | The highest rating value |
| N | = | Total number of the infected leaves |
On the other hand, the efficacy of each treatment was calculated as follows:
 |
Growth measurements: The averages of the morphological measurements in terms of plant height (cm), number of branches/plant, number of pods/plant, the weight of pod (g), hundred seed weight (g), length of the pod (cm), seed yield weight/plot (kg) and seed yield (ton/feddan) were recorded at harvest (90 days after sowing).
Physiological studies
Chemical analysis: Some biochemical constituents, in fresh leaves, including photosynthetic pigments (chlorophyll a, chlorophyll b, carotenoids and total pigments) were measured using the method of Lichtenthaler and Buschmann27. Total Soluble Sugars (TSS) were extracted according to Gomez et al.28 and determined according to Albalasmeh et al.29. The antioxidant activity (DPPH radical scavenging) was determined using the method of Liyana-Pathirana and Shahidi30. Indole acetic acid content was extracted and analyzed by the method of Gusmiaty et al.31. Phenolic content was measured as described by Maurya and Singh32. Total Soluble Protein (TSP) was determined according to the methods of Bonjoch and Tamayo33. Proline content was extracted and calculated according to the methodology of Tamayo and Bonjoch34. Free Amino Acids (Free AA) were extracted as outlined by Kalsoom et al.35 and determined with the ninhydrin reagent method reported by Verslues36.
Anatomical studies: For the anatomical study, samples were taken from the middle part of the leaflet blade. Samples were fixed and killed in FAA (formalin:glacial acetic acid:alcohol 5:5:90) for 48 hrs. Samples were washed in 50% ethyl alcohol, dehydrated and cleared in tertiary butyl alcohol series, embedded in paraffin wax of 54-56°C m.p. Vertical sections were cut at 15 μm by a rotary microtome, adhered by Haupt's adhesive and stained with the crystal violet-erythrosin combination, cleared in carbol xylene and mounted in Canada balsam, sections performed by the freehand made technique37. The observation and documented using an upright light microscope [Carl Zeiss then photoed by eyepiece digital camera (HIROCAM 5)]. Measurements were done, using a micrometre eyepiece and an average of 5 readings were calculated.
Statistical analysis: The obtained results were statistically analyzed38. Least Significant Difference (LSD) at 0.05 level of probability was applied for means comparison.
RESULTS
Efficacy of foliar spraying with different organic acids and coumarin against downy mildew, powdery mildew and rust on pea under field condition: Table 1 revealed that, all tested treatments significantly reduced the incidence of previously mentioned foliar diseases. Among the used all the tested treatments, citric and benzoic acids recorded the highest reduction in diseases severity of all tested diseases followed by salicylic and sorbic acids, respectively while coumarin caused recorded the lowest effect.
The highest reduction of downy mildew was obtained with citric and benzoic acids at 0.5% concentration by 72.63 and 71.40%, respectively, while coumarin recorded the lowest reduction by (46.43%) at the same concentration. Meanwhile, other treatments showed a moderate effect. The same results were obtained for the two other diseases with some differences in the reading values. The applied treatments have demonstrated their effect against fungal activities and were convenient in overcoming these diseases.
Changes in pea crop parameters under field conditions:Five organic acids i.e., citric, benzoic, salicylic, sorbic and boric acids in addition to coumarin at the concentrations of 0.25 and 0.50% were tested to study their effect on yield components of pea plants.
Table 2 demonstrate that all tried inducers gave a clear increase of crop parameters. This effect was increased with increasing inducers concentrations. Among the used inducers, citric acid caused the highest yield parameters followed by benzoic, salicylic and sorbic acids, respectively while coumarin caused the lowest. Where significant differences were found in some measures as a result of treatments such as plant height (cm), number of branches/plant, number of pods/plant, seed yield weight/plot (kg) and seed yield ton/feddan, while there were no significant differences between them, in the other treatments such as weight of pod (g), hundred seed weight, length of the pod (cm).
Changes in photosynthetic pigments of pea plants: Table 3 represents the influence of chlorophyll a, chlorophyll b, carotenoids and total pigment contents of a pea when treated with organic acids (salicylic, sorbic, citric, boric and benzoic) and coumarin.
All spraying treatments of the previous materials, generally, significantly (p<0.05) increased photosynthetic pigments compared to the control treatment. The highest (p<0.05) values were recorded when plants were sprayed with citric acid followed by benzoic and salicylic acids over that recorded for the other treatments. The maximum increases were obtained with citric acid foliar spraying at 0.50 mg L–1.
| Table 1: | Efficacy evaluation of foliar spray with different organic acids against some foliar diseases of pea |
| Disease severity (%) | |||||||
| Treatments | Concentration (%) | Downy mildew | Reduction (%) | Powdery mildew | Reduction (%) | Rust | Reduction (%) |
| Control | 0.00 | 55.83±1.36 | 0.00 | 39.23±5.28 | 0.00 | 28.20±1.31 | 0.00 |
| Salicylic acid | 0.25 | 24.70±0.75 | 55.75 | 15.43±0.63 | 60.67 | 8.37±0.49 | 70.32 |
| 0.50 | 18.08±1.46 | 66.33 | 9.63±0.92 | 75.45 | 5.43±0.58 | 80.74 | |
| Sorbic acid | 0.25 | 19.20±0.46 | 65.61 | 11.67±0.54 | 70.25 | 6.20±1.21 | 78.01 |
| 0.50 | 15.97±0.62 | 71.40 | 8.87±0.81 | 77.39 | 4.00±0.44 | 85.86 | |
| Citric acid | 0.25 | 20.83±1.23 | 62.69 | 11.90±0.83 | 69.67 | 6.16±0.56 | 78.16 |
| 0.50 | 15.28±0.64 | 72.63 | 6.81±0.76 | 82.64 | 4.40±0.51 | 84.40 | |
| Boric acid | 0.25 | 27.86±2.26 | 50.09 | 19.30±0.53 | 50.80 | 12.23±0.59 | 56.63 |
| 0.50 | 21.82±0.95 | 60.92 | 15.97±1.24 | 59.29 | 8.33±0.41 | 70.46 | |
| Benzoic acid | 0.25 | 19.44±1.40 | 65.18 | 17.80±0.25 | 54.63 | 11.67±0.90 | 58.62 |
| 0.50 | 17.69±1.22 | 68.31 | 14.67±0.49 | 62.61 | 6.33±0.32 | 77.55 | |
| Coumarin | 0.25 | 32.80±1.48 | 41.25 | 23.03±1.11 | 41.29 | 14.03±0.52 | 50.25 |
| 0.50 | 29.91±0.73 | 46.43 | 18.80±0.81 | 52.08 | 12.60±1.21 | 55.32 | |
| LSD 5% | 4.83 | - | NS | - | 3.65 | - | |
| SE: Standard error, LSD: Least significant difference at p<0.05 and NS: Not significant | |||||||
| Table 2: | Effect of spraying pea plants with different organic acids and coumarin on crop parameters under field conditions (Mean±SE) |
| Average of crop parameters | |||||||||
| Treatments | Concentration (%) | Plant height | Number of branches/plant | Number of pods/plant | Weight of pod (g) | 100-seed weight (g) | Length of pod (cm) | Seed yield weight/plot (kg) | Seed yield (t/fed) |
| Control | 0 | 64.00±1.57 | 8.50±1.21 | 10.33±0.88 | 7.20±0.81 | 46.00±1.15 | 9.30±0.42 | 8.80±0.49 | 3.91±0.15 |
| Salicylic acid | 0.25 | 73.40±0.83 | 15.50±1.37 | 11.33±0.88 | 9.80±0.62 | 48.80±1.37 | 11.20±0.47 | 11.00±0.60 | 4.22±0.24 |
| 0.5 | 77.50±2.25 | 21.50±1.27 | 18.00±1.17 | 11.50±0.93 | 69.20±1.42 | 12.30±0.47 | 12.30±0.44 | 4.58±0.31 | |
| Sorbic acid | 0.25 | 95.00±1.66 | 27.00±0.64 | 16.00±0.58 | 9.80±0.64 | 51.00±1.00 | 9.70±0.61 | 9.50±0.36 | 4.89±0.13 |
| 0.5 | 102.0±1.25 | 35.33±1.30 | 25.67±1.54 | 7.50±0.66 | 53.60±1.59 | 10.10±0.49 | 10.30±0.69 | 5.10±0.34 | |
| Citric acid | 0.25 | 95.30±1.24 | 11.33±1.45 | 12.33±0.88 | 10.30±0.93 | 54.80±0.62 | 11.00±0.35 | 11.50±0.75 | 5.11±0.23 |
| 0.5 | 97.20±1.22 | 25.00±1.57 | 20.00±1.15 | 11.50±1.14 | 60.80±1.17 | 12.50±0.76 | 12.00±0.87 | 5.47±0.15 | |
| Boric acid | 0.25 | 65.33±1.68 | 17.33±0.99 | 13.33±0.88 | 8.10±0.44 | 43.50±1.32 | 9.50±0.32 | 9.70±0.59 | 4.31±0.23 |
| 0.5 | 75.67±1.59 | 32.00±1.49 | 22.67±0.88 | 10.20±0.66 | 48.00±1.01 | 11.33±0.96 | 10.80±0.38 | 4.80±0.20 | |
| Benzoic acid | 0.25 | 66.67±1.30 | 9.67±0.96 | 8.50±0.67 | 8.20±0.99 | 45.00±0.61 | 10.20±0.61 | 9.20±0.32 | 4.09±0.20 |
| 0.5 | 75.67±7.78 | 19.00±0.85 | 13.00±1.00 | 9.00±0.64 | 51.60±1.17 | 11.33±0.38 | 10.00±0.51 | 4.44±0.30 | |
| Coumarin | 0.25 | 80.00±2.11 | 21.50±1.32 | 19.50±1.72 | 9.70±0.79 | 52.80±1.61 | 12.00±0.74 | 10.90±0.64 | 4.85±0.16 |
| 0.5 | 83.33±7.57 | 17.33±1.09 | 13.00±0.58 | 10.00±0.95 | 54.40±1.53 | 13.20±0.54 | 10.70±0.47 | 4.76±0.14 | |
| LSD 5% | 11.37 | 4.69 | 3.63 | NS | NS | NS | 2.11 | 0.79 | |
| SE: Standard error, LSD 5%: Least significant difference at p<0.05 and NS: Not significant | |||||||||
Changes in biochemical constituents: The changes in Total Soluble Sugar (TSS), Total Soluble Protein (TSP), proline, Free Amino Acid (FAA), total phenols, Indol Acetic Acid (IAA) and total antioxidants (DPPH %) were explored in Table 4.
It is clear that, all tested materials (chemical resistance inducers) (salicylic, sorbic, citric, boric, benzoic acids and coumarin) at both concentrations (0.25 and 0.50%) induced a significant (p<0.05) increase in all tested parameters compared with untreated plants. The highest value was observed in response to the highest concentration in general. It was observed that the maximum increases in the most biochemical constituents of pea plants were observed when the plants were sprayed with citric acid.
Effect of foliar spraying organic acids and coumarin on the anatomical characteristics of pea (Pisum sativum L.) leaves: Microscopically counts and measurements of specific histological characters in transverse sections through the blade of mature foliage leaf developed a the median portion of the leaflet of pea plant sprayed with 0.50% of citric acid and those of untreated plant is assumed in Table 5 and Fig. 1a-e.
As well, microphotographs explain these treatments are shown in the Fig. 1a-e. It is clear from Table 5 and Fig. 1b that, foliar application with citric acid at 0.50% recorded the highest increase in the thickness of upper and lower epidermal layers of pea leaflet by +92.08 and +70.38%, respectively, but foliar application with coumarin at the at 0.50% led to decrease in the thickness of the upper epidermal layer of pea leaflet by -6.59 and -15.31%, respectively less than the untreated plants. Also, the highest increase is recorded in the length and width of the vascular bundle by +50.82 and +52.83%, respectively more than the control (Fig. 1a) when a foliar application with citric acid at 0.50% (Fig. 1b), in this regard the length and width of vascular bundle of are slightly increased by +3.23 and 3.42%, respectively over the control when a foliar application with boric acid at 0.50% (Fig. 1c). Although the foliar application with coumarin at the same concentration led to a sharp decrement in the length of the vascular bundle by-46.59%, the led to a slight reduction in the width of the vascular bundle by -2.06% (Fig. 1d) less than the untreated plants (Fig. 1a).
| Table 3: | Effect of foliar spraying with different organic acids and coumarin on photosynthetic pigments of pea plants (Mean±SE) |
| Treatments | Concentration (%) | Chlorophyll a | Chlorophyll b | Carotenoids | Total pigments |
| Control | 8.67±0.23 | 1.71±0.07 | 2.57±0.21 | 12.95±0.52 | |
| Salicylic acid | 0.25 | 10.87±0.16 | 2.35±0.04 | 2.31±0.04 | 15.53±0.16 |
| 0.50 | 12.30±0.07 | 4.63±0.20 | 2.20±0.02 | 19.13±0.28 | |
| Sorbic acid | 0.25 | 10.61±0.45 | 1.70±0.11 | 2.78±0.27 | 15.09±0.83 |
| 0.50 | 12.04±0.42 | 2.96±0.51 | 2.92±0.10 | 17.92±1.04 | |
| Citric acid | 0.25 | 16.02±0.34 | 4.61±0.17 | 3.88±0.24 | 24.51±0.74 |
| 0.50 | 17.60±0.57 | 4.49±0.22 | 4.01±0.01 | 27.01±0.36 | |
| Boric acid | 0.25 | 9.70±0.11 | 3.96±0.04 | 2.80±0.07 | 16.46±0.08 |
| 0.50 | 11.39±0.19 | 3.95±0.13 | 3.05±0.01 | 18.39±0.05 | |
| Benzoic acid | 0.25 | 13.88±0.40 | 3.94±0.02 | 3.19±0.01 | 21.01±0.43 |
| 0.50 | 14.63±0.20 | 4.14±0.09 | 4.20±0.01 | 22.97±0.30 | |
| Coumarin | 0.25 | 9.47±0.29 | 2.31±0.11 | 3.48 ±0.11 | 15.26±0.29 |
| 0.50 | 10.25±0.15 | 3.46±0.01 | 3.92±0.05 | 17.63±0.10 | |
| LSD 5% | 1.12 | 0.62 | 0.60 | 1.93 | |
| SE: Standard error, LSD 5%: Least significant difference at p<0.05 and NS: Not significant | |||||
| Table 4: | Effect of foliar spraying with different concentrations of organic acids and coumarin on some compatible solutes, total phenols and indoles of pea plants (Mean±SE) | |||||||
| Compatible solutes (mg g–1 dry weight) | ||||||||
| Treatments | Concentration (%) | TSS | TSP | Proline | FAA | Phenol (mg g–1 dry weight) | IAA (mg g–1 dry weight) | DPPH (%) |
| Control | 291.92±4.88 | 189.20±11.22 | 76.79±0.50 | 965.76±17.60 | 123.21±3.99 | 28.72±0.58 | 29.85±1.01 | |
| Salicylic acid | 0.25 | 345.74±2.50 | 209.84±14.77 | 98.45±0.11 | 1164.53±73.71 | 130.58±1.08 | 31.08±0.93 | 35.45±1.25 |
| 0.50 | 423.48±3.13 | 224.89±1.31 | 116.33±0.95 | 1576.59±32.39 | 142.60±0.46 | 51.21±2.44 | 45.42±0.51 | |
| Sorbic acid | 0.25 | 426.04±4.62 | 174.91±3.54 | 113.12±0.44 | 1760.87±0.94 | 159.07±4.28 | 36.16±0.67 | 38.62±2.43 |
| 0.50 | 409.39±4.67 | 236.97±9.11 | 111.00±0.35 | 2124.71±84.92 | 155.02±0.17 | 43.46±0.35 | 35.75±1.81 | |
| Citric acid | 0.25 | 299.73±3.60 | 169.06±1.58 | 86.26±0.02 | 1528.02±19.81 | 153.03±0.21 | 30.56±0.87 | 26.82±0.47 |
| 0.50 | 373.86±5.60 | 244.35±1.86 | 127.31±0.29 | 2186.82±62.51 | 162.85±1.81 | 42.75±2.02 | 48.04±1.45 | |
| Boric acid | 0.25 | 233.91±8.66 | 162.71±1.18 | 96.98±0.58 | 1134.82±16.05 | 146.62±1.96 | 35.89±1.10 | 27.50±0.28 |
| 0.50 | 368.03±3.18 | 206.91±9.19 | 96.71±0.48 | 1729.81±31.05 | 156.28±0.71 | 41.94±4.76 | 39.88±1.54 | |
| Benzoic acid | 0.25 | 314.81±1.76 | 156.80±0.70 | 88.97±0.01 | 1884.09±12.38 | 160.41±6.57 | 28.14±0.58 | 27.37±0.55 |
| 0.50 | 440.99±8.46 | 187.07±8.24 | 105.39±0.25 | 2100.28±46.53 | 180.12±3.93 | 46.67±0.56 | 30.58±1.07 | |
| Coumarin | 0.25 | 257.54±5.94 | 174.47±0.06 | 81.47±0.85 | 1287.90±60.59 | 153.45±2.45 | 35.21±0.57 | 25.13±0.08 |
| 0.50 | 330.16±4.88 | 251.31±2.63 | 94.75±0.53 | 1404.73±6.15 | 157.87±4.47 | 39.47±0.99 | 46.44±1.35 | |
| LSD 5% | 89.98 | 20.10 | 1.91 | 210.20 | 13.06 | 5.74 | 4.65 | |
| SE: Standard error, LSD 5%: Least significant difference at p<0.05, TSS: Total soluble sugar, TSP: Total soluble protein, FAA: Free amino acid, IAA: Indol acetic acid and DPPH: Total antioxidants (DPPH) | ||||||||
| Table 5: | Effect of foliar spraying of organic acids and coumarin on the anatomical characteristics in pea plant leaves | |||||||
| Characters | ||||||||
| Epidermis thickness (μm) | Vascular bundle thickness (μm) | |||||||
| Upper epidermal layer (μm) | Lower epidermal layer (μm) | Length (μm) | Width (μm) | |||||
| Treatments | Absolute value | %±of control | Absolute value | %±of control | Absolute value | %±of control | Absolute value | %±of control |
| Control | 14.27 | 0.00 | 13.71 | 0.00 | 244.61 | 0.00 | 192.340 | |
| Salicylic acid | 18.36 | +28.66 | 16.9 | +23.26 | 258.23 | +5.56 | 194.97 | +1.36 |
| Sorbic acid | 22.76 | +59.49 | 21.2 | +54.63 | 247.87 | +1.33 | 195.51 | +1.64 |
| Citric acid | 27.41 | +92.08 | 23.36 | +70.38 | 368.95 | +50.83 | 293.98 | +52.84 |
| Boric acid | 17.46 | +22.35 | 19.66 | +43.39 | 252.52 | +3.23 | 198.93 | +3.42 |
| Benzoic acid | 20.76 | +45.48 | 17.41 | +26.98 | 260.77 | +6.6 | 199.18 | +3.55 |
| Coumarin | 13.33 | -6.59 | 11.61 | -15.31 | 130.63 | -46.59 | 188.48 | -2.06 |
| Characters | ||||||
| Mesophyllic tissue (μm) | ||||||
| Palisade tissue | Spongy tissue | Midrib zone thickness (μm) | ||||
| Treatments | Absolute value | %±of control | Absolute value | %±of control | Absolute value (μm) | %±of control |
| Control | 57.35 | 0.00 | 136.59 | 0.00 | 410.03 | 0.00 |
| Salicylic acid | 98.1 | +71.05 | 222.67 | +63.02 | 419.87 | +2.39 |
| Sorbic acid | 105.6 | +84.13 | 235.04 | +72.07 | 549.76 | +34.07 |
| Citric acid | 128.65 | +124.32 | 289.73 | +112.11 | 632.09 | +54.15 |
| Boric acid | 88.08 | +53.58 | 219.4 | +60.62 | 477.68 | +16.49 |
| Benzoic acid | 95.41 | +66.36 | 227.29 | +66.4 | 546.66 | +33.32 |
| Coumarin | 51.64 | -9.95 | 128.61 | -5.84 | 402.15 | -1.92 |
The alteration occurred on the thickness of leaflet blade either increasing or reduction corresponding to changes in the thickness of both of upper and lower epidermal layers, palisade and spongy tissues. In Table 5 and Fig.1b, it was clear that foliar application with citric acid at 0.50% recorded the highest increase in the thickness of mesophyllic tissue either palisade or spongy tissue by +124.32 and +112.11%, respectively more than the untreated plants (Fig. 1a). But the foliar application with coumarin at 0.50% recorded the decrement in the thickness of palisade and spongy tissues by -9.95 and -5.84%, respectively less than the untreated plants.
Although the foliar application with sorbic acid at 0.50% (Fig. 1e) led to an increase in the thickness of the midrib zone by +34.07% more than the control (Fig. 1a).
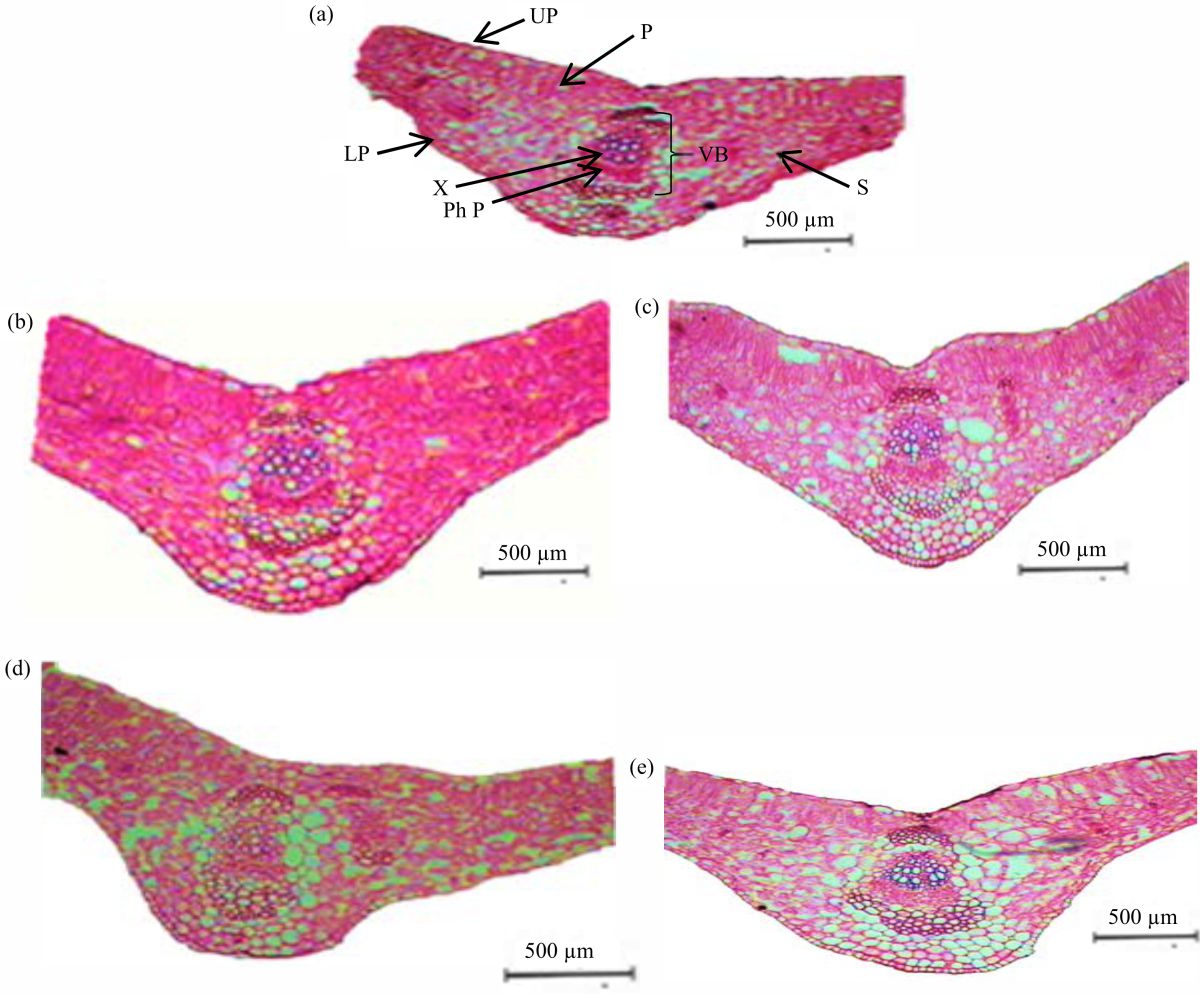 |
| Fig. 1(a-e): | Changes in transverse sections of the terminal leaflet blade of the fourth upper compound leaf on the mainstem of pea (Pisum sativum L.) plants grown under downy mildew, powdery mildew and rust diseases and application of the studied organic acids and coumarin at 0.50%, (a) Untreated plant leaves, (b) Plant leaves treated with citric acid, (c) Plant leaves treated with boric acid, (d) Plant leaves treated with coumarin and (e) Plant leaves treated with sorbic acid L: Lower epidermis, P: Palisade tissue, Ph: Phloem, S: Spongy tissue, U: Upper epidermis, Vb: Vascular bundle, X: Xylem vessels |
DISCUSSION
In Egypt, it is necessary to note that the conditions of the climate in these periods were characterized by high humidity and low temperatures which prevailed causing the emergence of fungal diseases on pea plants. During the experimentation season temperature and humidity were in ranges of 13-16°C and 69-74%, respectively and rainfall recorded 23.1-59.2 mm in the same season. Ash et al.39 showed that in early fall and late summer, under favourable weather conditions, humid nights, suitably warm, this leads to the emergence of fungal diseases.
Pea (Pisum sativum L.) was attacked by many foliar diseases. Downy mildew, powdery mildew and rust are the most common of these diseases. To control these diseases, certain chemical inducers have been utilized. Among the all tested treatments, citric and benzoic acids (Table 1) recorded the highest reduction in diseases severity for all tested diseases followed by salicylic and sorbic acids, respectively while coumarin recorded the lowest effect. The highest reduction of downy mildew was obtained with citric and benzoic acids at 0.5% concentration, while coumarin recorded the lowest reduction at the same concentration. Meanwhile, other treatments showed a moderate effect. The same results were obtained for the two other diseases with some differences in the reading values. In the present study, the efficiency of used chemical inducers could be attributed to their dynamic action in enhancing resistance and eliciting natural immunity of the host plants that may promote their defence mechanisms to fight the pathogens40-42. In this regard, various studies reported that salicylic acid proved to have stimulation activity in resisting disease. It has been reported that foliar application of cotton plants with potassium citrate and salicylic acid induced a significant rise in plant development and yield advantages as well as augmentation of the leaf chemical composition43. Moreover, Dalie et al.44 found that organic acids affected the plasmic membrane throughout changing its electrochemical properties thus increasing its permeability. With low pH values, a maximal concentration of protons is resulted and cause the augmentation in acid spreading across the plasmic membrane and cytoplasm. The results obtained herein are in agreement with Khalil et al.45, who explained that salicylic acid, potassium sorbate and ascorbic acid spraying treatments on pea plants suppressed the incidence of disease by about 12.4 and 17.1%. Also, Elgamal and Khalil26 studied that the effect of the foliar spray against powdery mildew on luffa in vitro and under field conditions with organic acids and essential oils, the results showed that all treatments significantly reduced powdery mildew of luffa caused by P. xanthii compared to untreated plants. In this regard, Ismail and Afifiorcid46 indicated that use some organic acids for control of rust disease on a bean plant. They added that salicylic, citric and fulvic acids were the most effective treatments in the reduction of rust under field conditions. It also led to an increase in growth parameters, total carbohydrates, chlorophyll and protein in bean plants over control. Fulvic, citric and salicylic acids were the most effective treatments in this regard. On the other hand, the highest activities of defence-related enzymes i.e., peroxidase, polyphenol oxidase and catalase were recorded in treated bean plants with organic acids.
All tested inducers gave a clear increase of crop parameters (Table 2). These results are in agreement with Ismail and Afifiorcid46, who indicated that, use some organic acids for the control of rust disease on bean plants. Results showed that salicylic, citric and fulvic acids were the most effective treatments in the reduction of rust under field conditions. It also led to an increase in growth parameters, total carbohydrates, chlorophyll and protein in bean plants over control. On the other hand, the highest activities of defence-related enzymes i.e., peroxidase, polyphenol oxidase and catalase were recorded in treated bean plants with organic acids. Similar results were recorded by Ramadan et al.47 on flax plants when treated with benzoic acid and coumarin and Abdallah et al.48 on wheat plants using salicylic acid.
All photosynthetic pigments (Table 3) showed generally a significant increase in pea plants when treated with organic acids (salicylic, sorbic, citric, boric and benzoic) and coumarin. Abd El-Gawad and Bondok17 found that total chlorophyll significantly decreased in leaves of infected tomato plants. They attributed the reduction in total chlorophyll of tomato leaves due to tomato mosaic virus infection to be a significance of the released transported toxins that induce the liberation of ROS49. Also, a high level of lipid peroxidation mediating cell damage in tomato tissues decreased its biomass and chlorophyll content15. The enhancement in these parameters of pigments induced by citric acid spraying is in line with Darandeh and Hadavi50 since the foliar application of citric acid at 0.075 and 0.15% w/v increased the chlorophyll contents of Lilium plants. Also, Hu et al.51 found that, foliar spraying of citric acid (0.2, 2 and 20 mM) ameliorate the adverse effect on chlorophyll content of heat-stressed tall fescue (Lolium arundinaceum) plants. Concerning the effect of salicylic acid on photosynthetic pigments, the maximum increase was observed in response to the highest concentration (0.50 mg L–1). Vicente and Plasencia52 stated that SA is an important regulator of photosynthesis due to its effects on leaf and chloroplast structure, stomata closure, chlorophyll and carotenoid contents. Khodary53 attributed the increase in photosynthetic capacity effect, with SA treatments on Zea maize plants under salinity stress, to its stimulatory effects on Rubisco activity and pigment contents. Moreover, SA is an antioxidant compound intense in the chloroplast and reported to protect the photosynthetic apparatus in drought stress conditions throughout scavenging the excessive ROS54. The primitive influence photosynthetic pigments induced by benzoic acid treatments are in harmony with Anjum et al.55 on soybean and Ramadan et al.47 on flax plants. They explained that the improvement effect of benzoic acid on photosynthetic pigments resulted from the increase in gas exchange, stomatal conductance, transpiration and photosynthetic rates. Coumarin also affected photosynthetic pigments, Saleh et al.56 on Vicia faba plants and Ramadan et al.47 on flax plants found that application of coumarin significantly increased its content. Khairy and Roh57 recorded a similar trend of improvement in chlorophyll contents of salinity stressed tobacco plants when treated with benzoic acid and p-coumaric acid. It is well established that boron has a direct role in photosynthesis as reviewed58 and its deficiency reduces chlorophyll and soluble protein contents of leaves, which results in Hill reaction's inhibition and net photosynthetic rate59.
It is clear that all tested materials (salicylic, sorbic, citric, boric, benzoic acids and coumarin) at both concentrations (0.25 and 0.50%) induced a significant increase in all determined parameters (Table 4) compared with untreated plants. Concerning the effect of salicylic acid on the tested biochemical constituents, similar results were shown by Abdallah et al.48 on wheat plants who reported that plant resistance to stress improved by the significant increase in TSS, proline, FAA and phenolic contents. In this concern, Tawfik et al.60 decided that increasing TSS, proline and free amino acids improved plant cells tolerance to salinity stress throughout increasing osmotic pressure in the cytoplasm as well as relative water contents essential for plant growth. In addition, Ashraf and Foolad61 stated that proline induces vital roles in preventing damaging effects of osmotic stresses expressed in forms of osmotic adjustment, stabilization and protection of enzymes, proteins and membranes. Also, Ahmadi et al.62 stated that proline content increased in grape buds under cold stress when treated with SA. In this concern, Jagesh et al.63 demonstrated that the osmotic adjustment in stressed plants resulted from the accumulation of high concentrations of osmotic compatible solutes, e.g., proline, glycinebetaine, soluble sugars and free AA. They also noticed that these substances express their important role in the adaptation of cells to various adverse environmental conditions via raising cytoplasm’s osmotic pressure, stabilizing proteins and membranes and maintaining the relatively high-water content obligatory for plant growth and cellular functions. Amino acids are primary metabolites that play essential roles in plant immunity against a wide range of pathogens. The variation in plant tissues amino acid quantity may determine the chance of environment for the pathogenic attackers like fungi, bacteria and viruses. This results finally either in strengthening plant defence to resist pathogenic attack effectively or surrender before vigorous infection64. The defences to biotic stress include morphological and structural barriers, chemical compounds and proteins and enzymes. These confer tolerance or resistance to biotic stresses by protecting products and by giving them strength and rigidity. The improvement effect of the used tested inducers on pea growth parameters (Table 2) may be attributed to the enhancement in the biosynthesis of photosynthetic pigments (Table 3) and/or its antioxidant effect (Table 4). These results are in the line with those recorded on maize using SA65 and Anjum et al.55 on soybean using benzoic acid. Ramadan et al.47 found an increase in total soluble sugar, total soluble protein, proline, free amino acids, total phenols, IAA contents and DPPH% of flax plants when sprayed with benzoic acid or coumarin. Also, they added that the increase in endogenous IAA content due to foliar spraying of the same materials resulted in enhancement of growth rate which stimulate cell division and/or enlargement21. In addition, amino acids play a vital role in enhancing secondary metabolites which alleviated the harmful effects of stress on plants66. Khairy et al.57 found that osmoprotectants (total soluble and insoluble sugar, totally soluble and insoluble protein) and antioxidant (total phenols) of Vicia faba plant' leaves significantly increased with coumarin application.
The results in Table 5 and Fig. 1a-e indicated that foliar application with 0.50% citric acid on pea plants led to the highest increase in the thickness of upper, lower epidermal layers, length and width of the vascular bundle and mesophyllic tissue either palisade or spongy tissue. Also, foliar application with boric acid at 0.50% produced a slight increase in the length and width of the vascular bundle more than the control. Although the foliar application with coumarin at the same concentration led to a decrease in the thickness of the upper epidermal layer and a sharp decrement in the length of the vascular bundle, a slight reduction in the width of the vascular bundle was recorded compared to the untreated plants. In this regard, Ali et al.67 on maize (Zea mays L.) reported that treatment with citric acid led to increasing upper and lower epidermal layers, length of the vascular bundle and mesophyll tissue, but the width of the vascular bundle was similar to the control. Concerning foliar application with salicylic acid, the results showed an increase in thickness in both palisade and spongy tissues. Moreover, the foliar application with sorbic acid at the same concentration led to an increase in the thickness of the midrib zone more than the control. These results are in agreement with Gomaa et al.18 who reported that foliar spraying with salicylic acid increased the thickness of all studied layers containing both palisade and spongy tissues. Some investigators confirmed the present findings using salicylic acid on other field crop plants, for instance, Farouk and Osman68 on Phaseolus vulgaris L.; Cárcamo et al.69 on Zea mays L., Nour et al.70 on bean and Gomaa et al.18 on Lupinus termis L. They found that salicylic acid application increased the thickness of the midvein and lamina of leaves.
CONCLUSION
The tested organic acids and coumarin at 0.25 and 0.50% are highly recommended to be applied twice for promoting vegetative growth, increasing productivity from pod and seeds, based on induced favourable changes in anatomical structures and endogenous secondary metabolites against powdery mildew, downy mildew and rust diseases that occurrence on pea plants under natural field conditions. The highest concentration (0.5%) of all tested materials were the most effective.
SIGNIFICANCE STATEMENT
From environmental and human health points of view, natural chemical inducers (natural organic antioxidants and phenolic compounds) proved to be of a great impact compared to pesticides which threaten globalization. The plant researchers should focus on the use of biological resources such as salicylic, sorbic, citric, boric, benzoic acids and coumarin to avoid the adverse effects of using other chemicals on the quantity and quality of the produced plants. This study represents guidance for researchers to uncover the critical areas of the positive impact for using similar substances which not only protect the plants from air-born diseases but also improve the morphological, physiological and anatomical structure. Thus, a natural material application for plant nutrition produce more healthy plants (resistant to biotic stresses and metabolic disorders).
REFERENCES
- Kumari, T. and S.C. Deka, 2021. Potential health benefits of garden pea seeds and pods: A review. Legume Sci., Vol. 3.
CrossRefDirect Link - Pandey, P., K.P.S. Kushwaha and V. Upadhyay, 2017. Evaluation of potential fungicides and bio agents for the management of pea downy mildew and yield under field condition. Int. J. Curr. Microbiol. Applied Sci., 6: 1381-1388.
CrossRefDirect Link - Fondevilla, S. and D. Rubiales, 2012. Powdery mildew control in pea. A review. Agron. Sustainable Dev., 32: 401-409.
CrossRefDirect Link - Latef, A.A.H.A. and H. Chaoxing, 2014. Does inoculation with Glomus mosseae improve salt tolerance in pepper plants? J. Plant Growth Regul., 33: 644-653.
CrossRefDirect Link - Mittova, V., M. Tal, M. Volokita and M. Guy, 2002. Salt stress induces up-regulation of an efficient chloroplast antioxidant system in the salt-tolerant wild tomato species Lycopersicon pennellii but not in the cultivated species. Physiol. Plant., 115: 393-400.
CrossRefDirect Link - Jones, J.D.G. and J.L. Dangl, 2006. The plant immune system. Nature, 444: 323-329.
CrossRefDirect Link - Király, L., B. Barna and Z. Király, 2007. Plant resistance to pathogen infection: Forms and mechanisms of innate and acquired resistance. J. Phytopathol., 155: 385-396.
CrossRefDirect Link - Ferreira, R.B., S. Monterio, R. Freitas, C.N. Santos and Z. Chen et al., 2007. The role of plant defence proteins in fungal pathogenesis. Mol. Plant Pathol., 5: 677-700.
CrossRef - Dahal, D., D. Heintz, A. Van Dorsselaer, H.P. Braun and K. Wydra, 2009. Pathogenesis and stress related, as well as metabolic proteins are regulated in tomato stems infected with Ralstonia solanacearum. Plant Physiol. Biochem., 47: 838-846.
CrossRefDirect Link - Ragab, M.M.M., A.M.A. Ashour, M.M. Abdel-Kader, R. El-Mohamady and A. Abdel-Aziz, 2012. In vitro evaluation of some fungicides alternatives against Fusarium oxysporum the causal of wilt disease of pepper (Capsicum annum L.). Int. J. Agric. For., 2: 70-77.
CrossRefDirect Link - Makoi, J.H.J.R. and P.A. Ndakidemi, 2007. Biological, ecological and agronomic significance of plant phenolic compounds in rhizosphere of the symbiotic legumes. Afr. J. Biotechnol., 6: 1358-1368.
Direct Link - Waqas, M., A.L. Khan, M. Hamayun, R. Shahzad, Y.H. Kim, K.S. Choi and I.J. Lee, 2015. Endophytic infection alleviates biotic stress in sunflower through regulation of defence hormones, antioxidants and functional amino acids. Eur. J. Plant Pathol., 141: 803-824.
CrossRefDirect Link - Hammerschmidt, R., 2005. Phenols and plant-pathogen interactions: The saga continues. Physiol. Mol. Plant Pathol., 66: 77-78.
CrossRef - Nicholson, R.L. and R. Hammerschmidt, 1992. Phenolic compounds and their role in disease resistance. Annu. Rev. Phytopathol., 30: 369-389.
CrossRefDirect Link - El-Khallal, S.M., 2007. Induction and modulation of resistance in tomato plants against Fusarium wilt disease by bioagent fungi (arbuscular mycorrhiza) and/or hormonal elicitors (jasmonic acid & salicylic acid): 1-Changes in growth, some metabolic activities and endogenous hormones related to defence mechanism. Aust. J. Basic Applied Sci., 1: 691-705.
Direct Link - Mandal, S., N. Mallick and A. Mitra, 2009. Salicylic acid-induced resistance to Fusarium oxysporum f. sp. lycopersici in tomato. Plant Physiol. Biochem., 47: 642-649.
CrossRefDirect Link - Abd El-Gawad, H.G. and A.M. Bondok, 2015. Response of tomato plants to salicylic acid and chitosan under infection with tomato mosaic virus. Am.-Eurasian J. Agric. Environ. Sci., 15: 1520-1529.
Direct Link - Gomaa, E.F., R.M.A. Nassar and M.A. Madkour, 2015. Effect of foliar spray with salicylic acid on vegetative growth, stem and leaf anatomy, photosynthetic pigments and productivity of Egyptian lupine plant (Lupinus termis forssk.). Inter. J. Adv. Res., 3: 803-813.
Direct Link - Chandra, R.P., R. Bura, W.E. Mabee, A. Berlin, X. Pan and J.N. Saddler, 2007. Substrate pretreatment the key to effective enzymatic hydrolysis of lignocellulosics. Adv. Biochem. Eng. Biotechnol., 108: 67-93.
CrossRef - Senaratna, T., D. Merritt, K. Dixon, E. Bunn, D. Touchell and K. Sivasithamparam, 2003. Benzoic acid may act as the functional group in salicylic acid and derivatives in the induction of multiple stress tolerance in plants. Plant Growth Regul., 39: 77-81.
CrossRefDirect Link - Abdallah, M.M.S., H.M.S. El-Bassiouny, T.A.E. Elewa and T.N. El-Sebai, 2015. Effect of salicylic acid and benzoic acid on growth, yield and some biochemical aspects of quinoa plant grown in sandy soil. Int. J. ChemTech. Res., 8: 216-225.
Direct Link - van Doorn, W.G., 2010. Water Relations of Cut Flowers. In: Horticultural Reviews, Janick, J. (Ed.), Vol. 18, John Wiley & Sons, Inc., New Jersey, ISBN-13: 9780471573340, pp: 1-85.
CrossRefDirect Link - da Silva, J.A.T., 2003. The cut flower: Postharvest considerations. J. Biol. Sci., 3: 406-442.
CrossRefDirect Link - Becker, L., A. Scheffczyk, B. Förster, J. Oehlmann, J. Princz, J. Römbke and T. Moser, 2011. Effects of boric acid on various microbes, plants, and soil invertebrates. J. Soils Sediments, 11: 238-248.
CrossRefDirect Link - El-Mohamedy, R.S.R., H. Jabnoun-Khiareddine and M. Daami-Remadi, 2014. Control of root rot diseases of tomato plants caused by Fusarium solani, Rhizoctonia solani and Sclerotium rolfsii using different chemical plant resistance inducers. Tunisian J. Plant Prot., 9: 45-55.
Direct Link - Elgamal, N.G. and M.S.A. Khalil, 2020. First report of powdery mildew caused by Podosphaera xanthii on Luffa cylindrica in Egypt and its control. J. Plant Prot. Res., 60: 311-319.
CrossRefDirect Link - Lichtenthaler, K.H. and C. Buschmann, 2001. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protocols Food Anal. Chem., Vol. 1.
CrossRefDirect Link - Gomez, L., E. Rubio and M. Augé, 2002. A new procedure for extraction and measurement of soluble sugars in ligneous plants. J. Sci. Food Agric., 82: 360-369.
CrossRefDirect Link - Albalasmeh, A.A., A.A. Berhe and T.A. Ghezzehei, 2013. A new method for rapid determination of carbohydrate and total carbon concentrations using UV spectrophotometry. Carbohydr. Polym., 97: 253-261.
CrossRefDirect Link - Liyana-Pathirana, C.M. and F. Shahidi, 2005. Antioxidant activity of commercial soft and hard wheat (Triticum aestivum L.) as affected by gastric pH conditions. J. Agric. Food Chem., 53: 2433-2440.
CrossRefDirect Link - Gusmiaty, A.M. Restu and R.Y. Payangan, 2019. Production of IAA (Indole acetic acid) of the rhizosphere fungus in the Suren community forest stand. IOP Conf. Ser.: Earth Environ. Sci., Vol. 343.
CrossRefDirect Link - Maurya, S. and D. Singh, 2010. Quantitative analysis of total phenolic content in Adhatoda vasica Nees extracts. Int. J. PharmTech Res., 2: 2403-2406.
Direct Link - Bonjoch, N.P. and P.R. Tamayo, 2001. Protein Content Quantification by Bradford Method. In: Handbook of Plant Ecophysiology Techniques. Roger, M.J.R. (Ed.), Springer, Dordrecht, Netherlands, ISBN-13: 978-0-306-48057-7, pp: 283-295.
CrossRefDirect Link - Tamayo, P.R. and N.P. Bonjoch, 2001. Free Proline Quantification. In: Handbook of Plant Ecophysiology Techniques, Roger, M.J.R. (Ed.)., Springer, Dordrecht, ISBN: 978-0-306-48057-7, pp: 365-382.
CrossRefDirect Link - Kalsoom, U., I.J. Bennett and M.C. Boyce, 2016. A review of extraction and analysis: Methods for studying osmoregulants in plants. J. Chromatogr. Sep. Tech., Vol. 7.
CrossRefDirect Link - Verslues, P.E., 2010. Quantification of Water Stress-Induced Osmotic Adjustment and Proline Accumulation for Arabidopsis thaliana Molecular Genetic Studies. In: Plant Stress Tolerance, Sunkar, R. (Ed.), Humana Press. pp: 301-315.
CrossRefDirect Link - Freund, R.J., W.J. Wilson and D.L. Mohr, 2010. Statistical Methods. 3rd Edn., Academic Press, United States, Pages: 796.
CrossRefDirect Link - Ash, A.S., R.P. Ellis, G.C. Pope, J.Z. Ayanian and D.W. Bates et al., 2000. Using diagnoses to describe populations and predict costs. Health Care Financing Rev., 21: 7-28.
Direct Link - El-Khallal, S.M., 2007. Induction and modulation of resistance in tomato plants against Fusarium wilt disease by bioagent fungi (arbuscular mycorrhiza) and/or hormonal elicitors (Jasmonic acid and Salicylic acid): 2-Changes in the antioxidant enzymes, phenolic compounds and pathogen related-proteins. Aust. J. Basic Appl. Sci., 1: 717-732.
Direct Link - Lopez-Reyes, J.G., D. Spadaro, M.L. Gullino and A. Garibaldi, 2010. Efficacy of plant essential oils on postharvest control of rot caused by fungi on four cultivars of apples in vivo. Flavour Fragr. J., 25: 171-177.
CrossRefDirect Link - Madukwe, E.U., J.O. Ezeugwu and P.E. Eme, 2013. Nutrient composition and sensory evaluation of dry Moringa oleifera aqueous extract. Int. J. Basic Appl. Sci., 13: 100-102.
Direct Link - Dalie, D.K.D., A.M. Deschamps and F. Richard-Forget, 2010. Lactic acid bacteria-potential for control of mould growth and mycotoxins: A review. Food Control, 21: 370-380.
CrossRefDirect Link - Khalil, M.S.A., N.G. El-Gamal, N.S. El-Mougy and M.M. Abdel-Kader, 2021. Molecular identification of Peronospora viciae causing downy mildew on field pea pods and its management. Indian Phytopathol., 74: 597-604.
CrossRefDirect Link - Ismail, A.M. and M.M.I. Afifiorcid, 2019. Efficacy of some biotic and abiotic factors in controlling common bean rust disease caused by Uromyces appendiculatus. Egypt. J. Phytopathol., 47: 313-329.
CrossRefDirect Link - Ramadan, A.A., A.E.S.M. Younis, B.A. Bakry and H.M.S. El-Bassiouny, 2020. Biochemical and yield of flax in responses to some natural antioxidant substances under sandy soil conditions. Asian J. Plant Sci., 19: 261-272.
CrossRefDirect Link - Abdallah, M.M.S., A.A.E. Ramadan, H.M.S. El-Bassiouny and B.A. Bakry, 2020. Regulation of antioxidant system in wheat cultivars by using chitosan or salicylic acid to improve growth and yield under salinity stress. Asian J. Pl. Sci., 19: 114-126.
CrossRefDirect Link - Shimizu, N., N. Hosogi, G.S. Hyon, S. Jiang, K. Inoue and P. Park, 2006. Reactive oxygen species (ROS) generation and ROS-induced lipid peroxidation are associated with plasma membrane modifications in host cells in response to AK-toxin I from Alternaria alternata Japanese pear pathotype. J. Gen. Plant Pathol., 72: 6-15.
CrossRefDirect Link - Darandeh, N. and E. Hadavi, 2012. Effect of pre-harvest foliar application of citric acid and malic acid on chlorophyll content and post-harvest vase life of Lilium cv. Brunello. Front. Plant Sci., Vol. 2.
CrossRefDirect Link - Hu, L., Z. Zhang, Z. Xiang and Z. Yang, 2016. Exogenous application of citric acid ameliorates the adverse effect of heat stress in tall fescue (Lolium arundinaceum). Front. Plant Sci., Vol. 7.
CrossRefDirect Link - Vicente, M.R.S. and J. Plasencia, 2011. Salicylic acid beyond defence: Its role in plant growth and development. J. Exp. Bot., 62: 3321-3338.
CrossRefDirect Link - Khodary, S.E.A., 2004. Effect of salicylic acid on the growth, photosynthesis and carbohydrate metabolism in salt stressed maize plants. Int. J. Agric. Biol., 6: 5-8.
Direct Link - Bakry, B.A., D.M. El-Hariri, M.S. Sadak and H.M.S. El-Bassiouny, 2012. Drought stress mitigation by foliar application of salicylic acid in two linseed varieties grown under newly reclaimed sandy soil. J. Applied Sci. Res., 8: 3503-3514.
Direct Link - Anjum, S.A., Ehsanullah, L. Xue, L. Wang, M.F. Saleem and C.J. Huang, 2013. Exogenous benzoic acid (BZA) treatment can induce drought tolerance in soybean plants by improving gas-exchange and chlorophyll contents. Aust. J. Crop Sci., 7: 555-560.
Direct Link - Saleh, A.M., M.M.Y. Madany and L. González, 2015. The effect of coumarin application on early growth and some physiological parameters in Faba Bean (Vicia faba L.). J. Plant Growth Regul., 34: 233-241.
CrossRefDirect Link - Khairy, A.I.H. and K.S. Roh, 2016. Effect of salicylic acid, benzoic acid and p-Coumaric acid on growth, chlorophyll, proline and vitamin C of salinity-stressed tobacco (Nicotiana tabacum). Int. J. Plant Soil Sci., 9: 1-10.
CrossRef - Mouhtaridou, G.N., T.E. Sotiropoulos, K.N. Dimassi and I.N. Therios, 2004. Effects of boron on growth, and chlorophyll and mineral contents of shoots of the apple rootstock mm 106 cultured in vitro. Biol. Plant., 48: 617-619.
Direct Link - Dixit, D., N.K. Srivastava and S. Sharma, 2002. Boron deficiency induced changes in translocation of 14CO2-photosynthate into primary metabolites in relation to essential oil and curcumin accumulation in turmeric (Curcuma longa L.). Photosynthetica, 40: 109-113.
CrossRefDirect Link - Tawfik, M.M., E.A. Badr, O.M. Ibrahim, E.M.A. Elhamid and M.S. Sadak, 2017. Biomass and some physiological aspects of Spartina patens grown under salt affected environment in South Sinai. Int. J. Agric. Res, 12: 19-27.
CrossRefDirect Link - Ashraf, M. and M.R. Foolad, 2007. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot., 59: 206-216.
CrossRefDirect Link - Ahmadi, J., B. Abedy, M. Shoor, S. Eshghi and R.A. Fahliani, 2017. The effect of salicylic acid and potassium sulphate on the amount of soluble sugars, starch, proline and protein of grape buds under cold stress. Rev. QUID, Special Issue: 2548-2556.
Direct Link - Tiwari, J.K., A.D. Munshi, R. Kumar, R.N. Pandey, A. Arora, J.S. Bhat and A.K. Sureja, 2010. Effect of salt stress on cucumber: Na+-K+ ratio, osmolyte concentration, phenols and chlorophyll content. Acta Physiologiae Plantarum, 32: 103-114.
CrossRefDirect Link - Tegeder, M., 2014. Transporters involved in source to sink partitioning of amino acids and ureides: Opportunities for crop improvement. J. Exp. Bot., 65: 1865-1878.
CrossRefDirect Link - Tuna, A.L., C. Kaya, M. Dikilttas, I. Yokas, B. Burun and H. Altunlu, 2007. Comparative effects of various salicylic acid derivatives on key growth parameters and some enzyme activities in salinity stressed maize (Zea mays L.) plants. Pak. J. Bot., 39: 787-798.
Direct Link - Hildebrandt, T.M., A.N. Nesi, W.L. Araújo and H.P. Braun, 2015. Amino acid catabolism in plants. Mol. Plant, 8: 1563-1579.
CrossRefDirect Link - Farouk, S. and M.A. Osman, 2009. The effect of plant defense elicitors on common bean (Phaseolus vulgaris L.) growth and yield in absence or presence of spider mite (Tetranychus urticae Koch) infestation. J. Agric. Sci. Mansoura Univ., 34: 11399-11419.
Direct Link - Cárcamo, H., C. Herle and V. Hervet, 2012. Greenhouse studies of thiamethoxam effects on pea leaf weevil, Sitona lineatus. J. Insect Sci., Vol. 12.
CrossRefDirect Link - Nour, K.A.M., N.T.S. Mansour and G.S.A. Eisa, 2012. Effect of some antioxidants on some physiological and anatomical characters of snap bean plants. New York Sci. J., Vol. 5.
Direct Link