
Riza Annisa Fauziah
Department of Plant Protection, Faculty of Agriculture, Universitas Gadjah Mada, Flora Street No. 1 Bulaksumur, Yogyakarta 55281, Indonesia
Tri Joko
Department of Plant Protection, Faculty of Agriculture, Universitas Gadjah Mada, Flora Street No. 1 Bulaksumur, Yogyakarta 55281, Indonesia
LiveDNA: 62.16906
Asian Journal of Plant Sciences
Year: 2021 | Volume: 20 | Issue: 2 | Page No.: 281-290







ABSTRACT
Background and Objective: Many pathogens have been reported to cause serious problems to orchid crops. The use of the resistant plant is considered as the most economical way and environmentally friendly in disease control. This study aimed to investigate and characterize Pto genes in several orchid species for potential disease management strategies. Materials and Methods: In this research, a Pto-type disease resistance gene was detected and characterized using degenerate Polymerase Chain Reaction (PCR) primers in several orchid species and database mining. A fragment of ~500 bp was amplified and sequenced. The identity of Serine/Threonine Kinases (STKs) from orchid was confirmed by comparisons of DNA and amino acid sequences with the GenBank database using BLASTX and BLASTP. The multiple alignment and phylogenetic analysis were performed using the Molecular Evolutionary Genetic Analysis (MEGA) program. Results: Alignment analysis showed that the 13 orchid species have conserved motives and domains included in the Pto section and phylogenetic analysis revealed that these species belong to class VIII protein kinase superfamily of the main class of plant resistance genes. Conclusion: The present study suggested that the orchid Pto-RGCs has provided insight into the common evolutionary histories of this resistance gene in orchid that represent a valuable sequence resource for potential use in genetic mapping that could assist in the development of disease resistance.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Riza Annisa Fauziah and Tri Joko, 2021. Characterization of Pto-like Protein Kinase Disease Resistance Genes in Orchid. Asian Journal of Plant Sciences, 20: 281-290.
DOI: 10.3923/ajps.2021.281.290
URL: https://scialert.net/abstract/?doi=ajps.2021.281.290
DOI: 10.3923/ajps.2021.281.290
URL: https://scialert.net/abstract/?doi=ajps.2021.281.290
INTRODUCTION
Orchidaceae is a plant family with spectacular flowers and high esthetic value. This family is one of the most diverse plant families that highly attract ornamental plant lovers1. However, orchid productivity can decrease because of the threat of diseases. Bacteria, fungi and viruses are commonly known pathogens associated with orchids and can cause various diseases, including bacterial soft rot, bacterial brown rot2,3, fungal root rot, black rot, Fusarium wilt, collar rot, anthracnose, leaf spot, petal blight4,5, Odontoglossum ringspot and Cymbidium mosaic6,7.
Plants can recognize and activate their defenses against pathogens8. Plant resistance is genetically controlled by one gene (monogenic) and many genes (multigenic). gene-for-gene interactions in plant resistance against diseases involve Resistance (R) genes in plants and avirulent (Avr) genes in interconnected pathogens. When interactions occur, a resistance response occurs and minimizes the spread of pathogens9,10. R-genes produce proteins that can detect the onset of infection and activate the resistance mechanism of plants against pathogenic infections11. These genes can strengthen and multiply defense signals and elicit a variety of responses, including the lignification of the cell wall and the production of antimicrobial metabolites, proteins related to pathogenesis, enzymes that protect oxidative stress and hormones related to stress, these responses also occur in the form of hypersensitive responses in death cells to limit the spread of pathogens in plant cells12,13.
The development of the presence of R-genes, including homologs and analogs, in plants can be used as one of the strategies in controlling various diseases in different plants. Advances in plant genome sequencing technology and biotechnology approaches, combined with increased knowledge about R-genes, can provide new insights into genetic breeding and help identify and apply efficient strategies in controlling plant diseases14,15. Eight classes of conserved domains, namely, NBS-LRR-TIR, NBS-LRR-CC, LRR-TrD, LRR-TrD-Kinase, TrD-CC, TIR-NBS-LRR-NLS-WRKY, LRR-TrD-PEST-ECS and enzymatic R-genes (protein kinase superfamily [PKs]), have been identified in R-genes. The NBS-LRR is the largest class that can be found in plant genomes16. One of the most studied R-genes associated with gene-for-gene interactions is the Pto gene, which is a form of the signaling pathway providing resistance to Pseudomonas syringae pv. in tomato plants17. A study on the Pto-type disease R-gene has shown that it provides resistance not only to P. syringae pv. tomato but also to other important pathogens18. Pto is an R-gene product that belongs to class VIII
of the R-gene class and is a member of PKs19. The Pto protein plays a role in early infection recognition and signal transduction to activate the defense mechanism when the plant is infected. The Pto gene encodes Serine/Threonine Kinase (STK) that physically interacts with Avr Pto and Avr PtoB proteins from P. syringae pv. tomato and expresses its endurance through hypersensitive responses20.
The presence of R-genes can be identified through a Polymerase Chain Reaction (PCR) with the degenerate primer targeting the conserved domain. This method has been successfully carried out in Solanum21, Phaseolus vulgaris22, Citrus23, Vitis vinifera24, Musa acuminata25, Fragaria ananassa26 and Capsicum27. Pto-like homologs have also been characterized in Phalaenopsis spp.28. The present study aimed to identify the presence of disease R-genes (Pto) in several orchid species through PCR amplification by using degenerate primers and to characterize Pto genes in several orchid species.
MATERIALS AND METHODS
Study area: Orchid plants were purchased from nurseries in Yogyakarta, Indonesia. Isolation of genomic DNA and PCR analyses were carried out at the Laboratory of Plant Pathology, Department of Plant Protection, Faculty of Agriculture, Universitas Gadjah Mada, Yogyakarta (Indonesia). The study was conducted during the May-November, 2019.
Plant materials and DNA extraction: The plant materials used were 13 orchid species, namely, Maxillaria tenuifolia, Vanda helvola, Oncidium millennium Gold, Paphiopedilum javanicum, Cymbidium artopurpureum, Coelogyne trinervis, Dendrobium phalaenopsis, Ascocentrum miniatum, Bulbophyllum lepidum, Rhynchostylis retusa, Eria hyacinthoides, Phaius tankervilleae var. alba and Phalaenopsis amabilis. Healthy young leaves (0.2 g) were prepared for DNA extraction based on the cetyltrimethylammonium bromide (CTAB) method, which was modified with the addition of PVP and β-mercaptoethanol29. Then, 0.2 g of young orchid leaves was crushed with a 1.5 mL of extraction buffer solution and incubated at 65°C for 20 min. The sample was transferred and centrifuged at a speed of 10,000 rpm for 10 min. Afterward, 700 μL of supernatant was transferred to a new 1.5 mL microtube and 600 μL of CIAA solution was added. The solution was mixed, shaken for 3-5 min and centrifuged at 10,000 rpm for 10 min. A total of ±600 μL supernatant was transferred to a new 1.5 mL microtube and added with ethanol to obtain a volume of 1.5 mL. The mixture was shaken for 10 min until pellets appeared and then centrifuged at 10,000 rpm for 10 min. Ethanol was discharged and isopropanol was added until a volume of 1.5 mL was reached. The mixture was centrifuged at a speed of 10,000 rpm for 10 min. Isopropanol was discharged and DNA was dried. Aquabidest (40 μL) was added to dissolve the DNA. The quality and quantity of the DNA were observed using Nanodrop (Maestro Nano, USA)30.
PCR amplification using Pto degenerate primers: PCR amplification was carried out using a genomic DNA with a pair of degenerate primers, forward primer (PtoF: 5’-ggagg atttggtaargtntarar-3’) and reverse primer (PtoR: 5’-accaca ccaaatgartanacrtc-3’)28. The reaction was done in 25 μL reaction volume using a GoTaq Green Kit (Promega) containing 2.5 μL of DNA template, 1.5 μL of forward and reverse primer (10 μM), 12.5 μL of GoTaq Green master mix and 7 μL of nuclease-free water. As the initial denaturation, the tubes were placed in a PCR machine (Biorad T100, Germany)31.
Amplification was performed in accordance with previously described methods28 with slight modification as follows: initial denaturation at 95°C for 3 min, followed by 35 amplification cycles. Each amplification cycle consisted of denaturation at 95°C for 30 sec, annealing at 45.9°C for 1 min, extension at 72°C for 1 min and a final extension at 72°C for 10 min. Furthermore, a qualitative test was performed using electrophoresis techniques and visualization was carried out with a UV Trans-Illuminator (Biorad)32.
Nucleotide sequence determination of Pto-fragment: The PCR results were sent to a DNA sequencing service provider (First BASE, Malaysia) to determine nucleotide sequences by using Big Dye terminator sequencing kits (Applied Biosystems). Sequencing products were separated with ABI 3730 automatic sequencers (Applied Biosystems)33.
Sequence editing and similarity analysis: The sequencing results were analyzed with bioinformatics techniques by using MEGA 7.0 software and compared with other Pto accessions in GenBank (https://www.ncbi.nlm.nih.gov/) by using the BLAST program. BLAST analysis was carried out at https://BLAST.ncbi.nlm.nih.gov/BLAST.cgi. BLAST results were used to infer the general identity of the acquired amplicons and similarity analysis was performed34.
Collection of other Pto-like PKs by mining databases: A comparison was made by using the Pto gene collection from the GenBank database (http://www.ncbi.nlm.nih.gov/) as comparison genes. The domains in the Pto sequence was examined by performing multiple sequence alignment based on the corresponding regions of the Pto-type disease resistance PK of Solanum pimpinellifolium (GenBank: A49332) and the Fen protein of S. pimpinellifolium (GenBank: AAC48932). In this alignment, the amino acid of the orchid began at number 67 of the amino acid of Pto disease resistance PK and Fen protein. The collections were also used for phylogenetic analysis. The Pto-type disease resistance PK and four paralogs (Fen, Lpimpth2, Lpimpth3 and Lpimpth4), which were parts of the Pto in tomato plant, of S. pimpinellifolium and other accessions from NCBI were used. The genes from the CC-NBS-LRR class, which is class II of the R-gene, namely, PKs from Arabidopsis thaliana and Pto-type disease resistance PK in tomato plants were also added. The data from PK members in A. thaliana was added because at least 44 subfamilies of 610 PK members in A. thaliana with different motives from their domain have been identified as such, the candidate of Pto R-genes in orchids can be easily identified35.
Phylogenetic analysis: Phylogenetic analysis was carried out to evaluate the genetic relationship between the Pto candidates of the 13 orchid species and Pto of other plants obtained from the NCBI collection. A phylogenetic tree was made using a model from the analysis of the Bayesian information criterion scores via maximum likelihood and bootstrap (1,000 replicates) analysis in MEGA software36,37.
RESULTS AND DISCUSSION
Identification of Pto-like genes in orchid: With technological advancements, millions of DNA sequences whose quality is highly dependent on the speed of work procedures and the technology used can be analyzed. This study used degenerate primers and based on the amino acid regions conserved from subdomains I and IX, which are identical to STKs, Pto, Fen and Ptil genes from tomato R-genes and MHK and APK1 from A. thaliana R-genes. The results of the PCR electropherogram with a 100 bp DNA marker are shown in Fig. 1. The electrophoresis results revealed that DNA bands were generated in the expected size of ~500 bp of the DNA marker. This result was in accordance with that of previously reported by Elina et al.28 that characterized Pto gene from 20 Phalaenopsis orchid genotypes.
Multiple sequence alignment of Pto-like genes: The sequencing results were evaluated in terms of their similarity to the conserved domain in the NCBI conserved domain database.
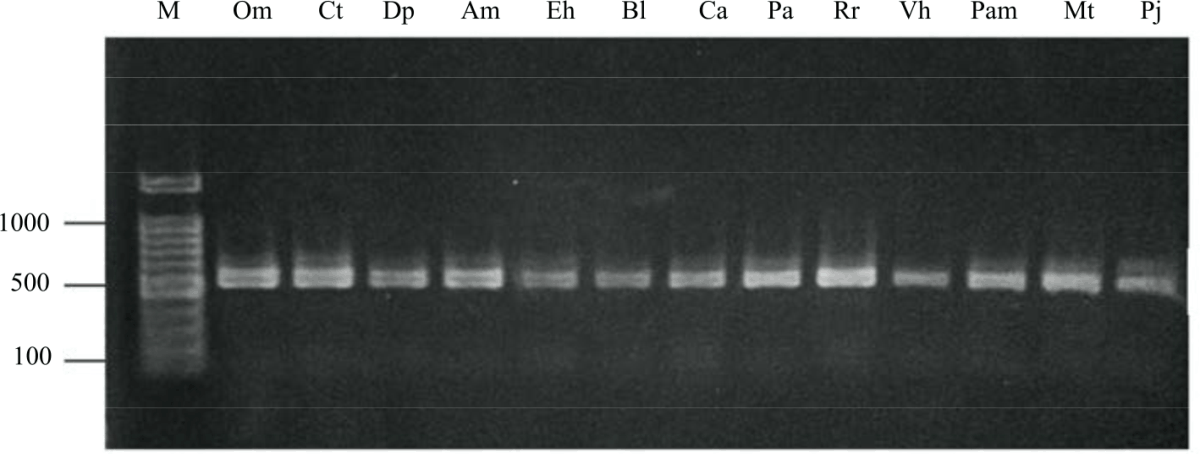 |
| Fig. 1: | Amplification of Pto-like gene from 13 orchid species by using the degenerate PtoF/PtoR primers The PCR product was analyzed through electrophoresis on 1% agarose gel in 1× TBE and amplified at the expected size of ∼500 bp. Om: Oncidium millennium Gold, Ct: Coelogyne trinervis, Dp: Dendrobium phalaenopsis, Am: Ascocentrum miniatum, Eh: Eria hyacinthoides, Bl: Bulbophyllum lepidum, Ca: Cymbidium artopureum, Pa: Phaius tankervilleae var. alba, Rr: Rhynchostylis retusa, Vh: Vanda helvola, Pam: Phalaenopsis amabilis, Mt: Maxillaria tenuifolia, Pj: Paphiopedilum javanicum, M: 100 bp DNA marker (Promega) |
The presence of the conserved domain was analyzed to assure that the amino acid of orchids is a part of Pto. The results showed that the high identity similarity of the translated amino acids was in the STK domain of the STK subfamily, namely, interleukin-1 receptor-associated kinases, which are a part of the Pto catalytic domain.
Multiple sequence alignment was performed to predict the amino acid sequences of the 13 orchid species by using the Pto gene obtained from BLAST as well as the Pto disease resistance PK and Fen protein in S. pimpinellifolium. The alignment revealed that several features of the Pto protein in plant serine/threonine kinases were highly conserved in the corresponding region of all orchid Pto-RGCs, such as the STK subdomains internal to the degenerate primer sequences. The indicated T and S revealed catalytic domain of threonine and serine (Fig. 2a). The alignment also showed the deletion of two amino acids and the insertion of three amino acids with regards to all orchid Pto-like sequences (Fig. 2b). The other important part in Pto sequence analysis was the activation domain and its internal P+1 loop site of protein kinases, which is responsible for the specific binding of Avr Pto, which were also conserved in the Pto-like sequences from orchid (Fig. 2c).
The Pto sequence analysis showed that several conserved subdomains are important for catalytic activity and some active amino acid residues are essential for Pto function27. The catalytic domain is in the form of a PK consisting of threonine and serine, which participate in the activation of resistance response to pathogenic bacteria38. The active site of the STK type enzyme39 is also present in the Pto catalytic domain, which is characterized by the presence of a DFG amino acid residue motif and terminated by the PE motif. The DFG and PE motifs are separated by 25 amino acid residues in the middle.
| Table 1: Deletion and insertion analysis of the Pto-like gene of Solanum pimpinellifolium with the corresponding Pto-type gene from 13 orchid species | ||
| Species | Deletion 133-134 | Insertion 158-160 |
| Pto Solanum pimpinellifolium | TM | |
| Vanda helvola | GSA | |
| Cymbidium artopurpureum | GAE | |
| Bulbophyllum lepidum | GSA | |
| Rhynchostylis retusa | GSA | |
| Phaius tankervilleae var. alba | GPK | |
| Ascocentrum miniatum | GSA | |
| Coelogyne trinervis | GKR | |
| Eria hyacinthoides | GSA | |
| Dendrobium phalaenopsis | GSA | |
| Oncidium millennium Gold | GAE | |
| Maxillaria tenuifolia | GSA | |
| Paphiopedilum javanicum | GSA | |
| Phalaenopsis amabilis | GSA | |
| G: Glycine, S: Serine, A: Alanine, E: Glutamic, R: Agrinine, K: Lysine, P: Proline | ||
The active site has a P+1 loop domain, which is marked by the G[S/T][F/L]GY[L/I]DPE amino acid residue motif at the end of the activation domain or activation site. The P+1 loop domain is associated with Avr Pto and serine and threonine residues are autophosphorylation sites. When the P+1 loop domain interacts with Avr Pto, serine and threonine residues undergo autophosphorylation and cause signal transduction to activate plant resistance mechanisms40.
Sequencing analysis can show that proteins encoded by R-genes provide several conserved domains, including Nucleic Binding Sites (NBS), Leucine Rich Repeats (LRRs), transmembrane domains, Coiled Coil Domains (CCs) and STKs41.
The amino acids of Pto-type disease resistance PK of S. pimpinellifolium and the Fen protein of S. pimpinellifolium were subjected to sequence alignment to examine the conserved domain in Pto candidates that could encode resistance genes in orchids. The results (Table 1) showed the amino acid deletions in the 133-134 sequence and the insertion of three amino acids in the 158-160 sequence.
 |
| Fig. 2(a-c): | Multiple amino acid sequence alignment of the 13 orchid species and the Pto protein, together with the corresponding region of the Pto resistance protein and the Fen protein of Solanum pimpinellifolium (a) Conserved amino acids in plant serine/threonine kinases and Pto autophosphorylation sites (b) Deletion and insertion, (c) Activation domain and P+1 loop site of protein kinases |
 |
| Fig. 3: | Phylogenetic tree based on the alignment of the deduced amino acid sequences of the Pto-like genes from 13 orchid species and its four paralogs (Fen, LpimPth2, pimPth3 and LpimPth4) The tree was constructed using the maximum likelihood method with Tamura 3-Parameter+Gamma distribution+Invariable best fit model provided in MEGA 7.0. Stability of the tree was assessed with 1,000 bootstrap replications |
| Table 2: Autophosphorylation site analysis of Pto-like gene from Solanum | ||||||||||||||||||
| Autophosphorylation sites | ||||||||||||||||||
| Species | T84 | T157 | T184 | T193 | T198 | T202 | T207 | S75 | S90 | S99 | S129 | S135 | S137 | S170 | S189 | S201 | ||
| Pto Solanum pimpinellifolium | T | T | T | T | T | T | T | S | S | S | S | S | S | S | S | S | ||
| Vanda helvola | T | T | A | P | T | T | S | S | S | S | S | N | N | S | S | S | ||
| Cymbidium artopurpureum | T | T | A | P | T | T | S | S | S | T | S | P | S | T | S | S | ||
| Bulbophyllum lepidum | T | T | A | P | T | T | S | S | S | S | S | N | N | S | S | S | ||
| Rhynchostylis retusa | T | T | A | P | T | T | S | S | S | S | S | N | N | S | S | S | ||
| Phaius tankervilleae var. alba | T | T | P | P | T | T | S | S | S | S | P | H | N | P | S | S | ||
| Ascocentrum miniatum | T | T | A | P | T | T | S | S | S | S | S | N | N | S | S | S | ||
| Coelogyne trinervis | T | T | A | P | T | T | S | S | S | S | S | N | N | T | S | S | ||
| Eria hyacinthoides | T | T | A | P | T | T | S | S | S | S | S | N | N | S | S | S | ||
| Dendrobium phalaenopsis | T | T | A | P | T | T | S | S | S | S | S | N | N | S | S | S | ||
| Oncidium millennium Gold | T | T | A | P | T | T | S | S | S | A | S | P | S | T | S | S | ||
| Maxillaria tenuifolia | T | T | A | P | T | T | S | S | S | S | S | N | N | S | S | S | ||
| Paphiopedilum javanicum | T | T | A | P | T | T | S | S | S | S | S | N | N | T | S | N | ||
| Phalaenopsis amabilis | T | T | A | P | T | T | S | S | S | S | S | N | N | S | S | S | ||
| T: Threonine, A: Alanine, P: Proline, S: Serine, N: Asparagines, H: Histidine | ||||||||||||||||||
This result indicated that the Pto gene candidates in the orchids evolved, resulting in diversity. These insertion and deletion also occur in other Pto plants, such as chili in the sequence of 133 and 134 and 159-161, respectively27.
The conserved serine and threonine autophosphorylation sites of Pto-type disease resistance PK and the Fen protein were present in several amino acid sequences of orchids (Table 2). Serine autophosphorylation sites can be found in the amino acid sequences of 75, 90, 99, 129, 135, 137, 170, 189 and 201. However, in C. artopurpureum, amino acids differed in the sequences of 99, 135 and 170, which were replaced by T, P and T, respectively. In P. tankervilleae var. Alba, differences in amino acids were also found in the sequences of 129, 135, 137 and 170, which were replaced by P, H, N and P, respectively. In O. millennium Gold, the amino acid sequences of 99, 135 and 170 were replaced by A, P and T. In P. javanicum, the amino acid sequences of 135, 137, 170 and 201 were replaced by N, N, T and N. In C. trinervis, the amino acid sequences of 135, 137 and 170 were replaced by N, N and T. In E. hyacinthoides, P. amabilis, V. helvola, R. retusa, M. tenuifolia, D. phalaenopsis, B lepidum and A. miniatum, the differences were in the amino acid sequences of 135 and 137 that were both replaced by N. Among all the sequences, S75, S90 and S189 were the strongest conservation areas.
 |
| Fig. 4: | Phylogenetic tree construction of the deduced amino acid sequences of the Pto-like genes from 13 orchid species and other Pto-like genes retrieved from Genbank The tree was constructed using the maximum likelihood method with Jones, Taylor and Thornton+ Gamma (JTT+G) best fit model assessed with 1,000 bootstrap replications |
This autophosphorylation site has a regulatory function and is important for kinase activity that induces plant defenses, including HR mechanisms during interactions with Avr Pto. The P+1 loop and activation domains are also conserved in the amino acids of Pto in orchids that have been marked. They are responsible for Avr Pto-specific binding. All areas that can be conserved likely cause orchid resistance to pathogens. The results revealed that the 13 orchid species have conserved motifs and domains that are part of Pto, so these plants are included in the Pto section.
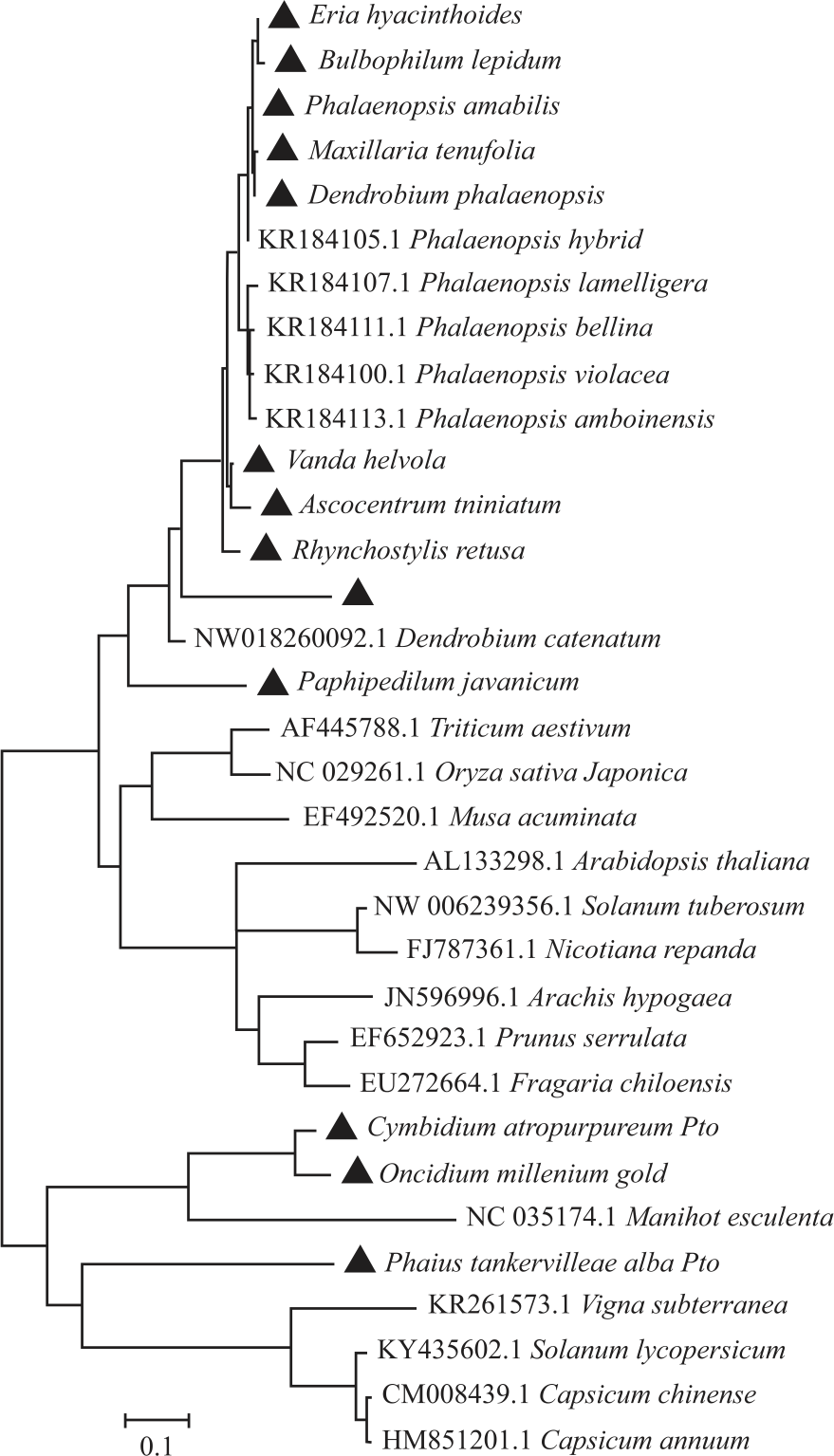
Phylogenetic relationships of the serine/threonine kinase sequences in orchid: Phylogenetic analysis results clearly showed that the 13 orchid species were included in the Pto like-kinase class because of their high similarity to Pto-type disease resistance PK and four paralogs of S. pimpinellifolium (Fen, Lpimpth2, Lpimpth3 and Lpimpth4) with CC-NBS-LRR of A. thaliana as the outgroup. These results indicated that the 13 orchid species were included in class VIII PK encoded by STK (Fig. 3).
Further phylogenetic determination revealed that Pto-type disease resistance genes in E. hyacinthoides, D. phalaenopsis, B. lepidum, P. amabilis, V. helvola, A. miniatum, R. retusa, C. trinervis and P. javanicum have high similarity to those in Phalaenopsis hybrid cultivar, Phalaenopsis lamelligera, Phalaenopsis amboinensis, Phalaenopsis bellina, Phalaenopsis violacea and Dendrobium catenatum. Pto R-genes in O. millennium Gold and C. artopurpureum have high similarity to those in Manihot esculenta and Pto-type disease R-genes in P. tankervilleae var. alba have high similarity to those in Vigna subterranean, Solanum lycopersicum, Capsicum chinense and Capsicum annum (Fig. 4).
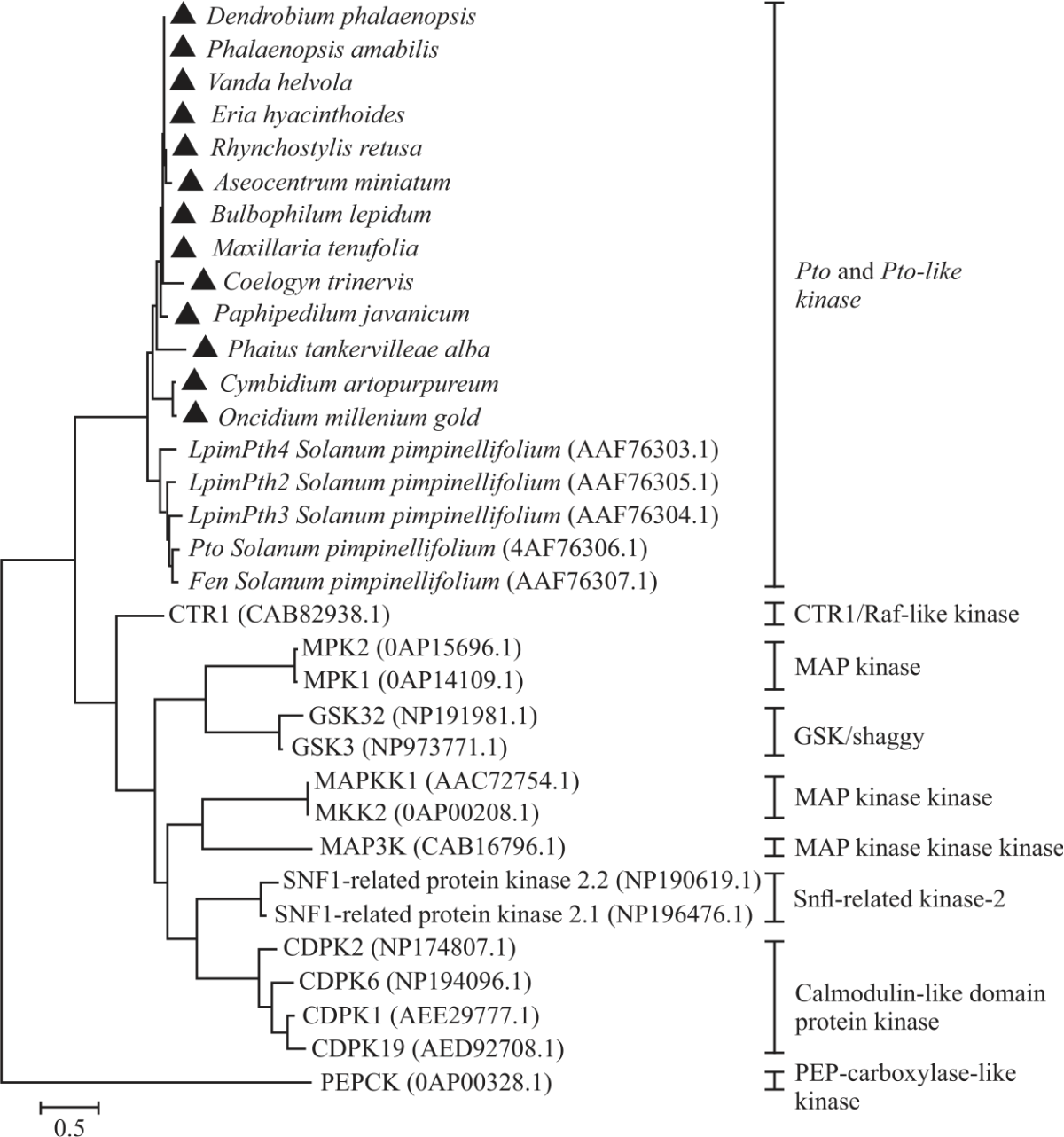 |
| Fig. 5: | Phylogenetic tree construction of the deduced amino acid sequences of the Pto-like genes from 13 orchid species, Pto disease resistance protein kinase and its four paralogs (Fen, LpimPth2, pimPth3, and LpimPth4) The tree was constructed using the maximum likelihood method with Jones, Taylor and Thornton+ Gamma (JTT+G) best fit model assessed with 1,000 bootstrap replications |
The result suggested that Pto-like kinases in orchid were divided into two clusters in which some of them were more closely related to monocotyledonous while the other have closely related to dicotyledonous plant species. It is interesting to note that O. millennium, C. artopurpureum and P. tankervilleae were more closely related to Pto sequences from dicot families than to Orchidaceae suggesting that the origin of this type of sequence may have preceded he divergence of monocot and dicot plants. Previously reported research on banana Pto-RGCs25 also revealed similar finding that the banana Pto-RGCs were more closely related to Pto-RGCs from other plant species than each other.
To elucidate the evolutionary history, the STK sequences (including Pto-like genes) from orchid and its four paralogues (Fen, LpimPth2, LpimPth3 and LpimPth4) were included in the phylogenetic analysis. As expected, the Pto-like genes from orchid and its four paralogues were grouped together in one branch, which represents the Pto and Pto-like kinase subfamily, supported by the highest bootstrap value. Phylogenetic analysis showed that the 13 orchid species could be classified as PK members of A. thaliana and be part of the Pto-like kinase. The other subfamilies were calmodulin-like protein kinase, PEP carboxylase-like kinase, Snf1-related kinase-2, Map kinase, MAP kinase kinase kinase and GSK/shaggy that can represent the diversity of PK members (Fig. 5). Phylogenetic analysis of Pto and Pto-like sequences have suggested that these sequences form a unique group of kinases in plants. In agreement with this finding, the orchid Pto-like kinase formed a highly supported group with the Pto disease resistance protein suggesting that these sequences share a common evolutionary origin with the tomato Pto protein and possibly a similar function in disease resistance.
This study has uncovered a set of orchid Pto-like sequences and provided the first insights about their amino acid sequence structure and evolution. Phylogenetic analysis indicated that this type of genes may have evolved before the divergence of monocotyledonous and dicotyledonous plants. The presence of several conserved amino acids in the orchid Pto-like sequences that are crucial for Pto function and the fact that these sequences were phylogenetically closely related to Pto, make of them a valuable sequence resource for plant-pathogen interaction studies in orchid. Furthermore, current results could be used as a basis for controlling diseases in orchids through the breeding of resistant varieties.
CONCLUSION
PCR with PtoF/PtoR degenerate primers produced the expected size of 500 bp for the 13 orchid species (E. hyacinthoides, P. amabilis, V. helvola, P. javanicum, C. trinervis, R. retusa, O. millennium Gold, C. artopurpureum, M. tenuifolia, D. phalaenopsis, B. lepidum, P. tankervilleae var. Alba and A. miniatum). Alignment results revealed that all 13 orchid species have conserved motifs and domains that are included in the Pto section. Phylogenetic analysis showed that the Pto-like kinase gene in the 13 orchid species belongs to class VIII of PKs and is a product of the R-gene.
SIGNIFICANCE STATEMENT
This research described the identification of orchid STKs, their structure and evolution and uncovers a set of Pto-like candidates. These findings will contribute to expanding our knowledge of the roles of Pto-like genes in orchid-pathogen interactions. Furthermore, the phylogenetic analysis revealed that the orchids Pto-RGCs were clustered with Pto suggesting a common evolutionary origin with this R gene. Thus, a new theory on genetic breeding applications that will provide foundation for the development of disease resistance may be arrived at.
ACKNOWLEDGMENT
This study is financially supported by Universitas Gadjah Mada under RTA Program 2019 with contract No. 3220/UN1/ DITLIT/DIT-LIT/LT/2019.
REFERENCES
- Cai, J., X. Liu, K. Vanneste, S. Proost, W.C. Tsai and K.W. Liu et al., 2015. The genome sequence of the orchid Phalaenopsis equestris. Nat. Genet., 47: 65-72.
CrossRefDirect Link - Keith, L.M., K.T. Sewake and F.T. Zee, 2005. Isolation and characterization of Burkholderia gladioli from orchids in hawaii. Plant Dis., 89: 1273-1278.
CrossRefDirect Link - Joko, T., A. Subandi, N. Kusumandari, A. Wibowo and A. Priyatmojo, 2014. Activities of plant cell wall-degrading enzymes by bacterial soft rot of orchid. Arch. Phytopathol. Plant Prot., 47: 1239-1250.
CrossRefDirect Link - Smitamana, P. and R.J. McGovern, 2018. Diseases of Orchid. In: Handbook of Florists' Crops Diseases. McGovern, R. and W. Elmer (Eds.), Springer International Publishing, Cham, ISBN: 978-3-319-39670-5 pp: 633-662.
CrossRefDirect Link - Zettler, F.W., N.J. Ko, G.C. Wisler, S. Elliot Mark and S.M. Wong, 1990. Viruses of orchids and their control. Plant Dis., 74: 621-626.
CrossRefDirect Link - Mahfut, T. Joko and B.S. Daryono, 2016. Molecular characterization of Odontoglossum ringspot virus (ORSV) in Java and Bali, Indonesia. Asian J. Plant Pathol., 10: 9-14.
CrossRefDirect Link - Joko, T., M. Umehara, T. Murata, H. Etoh, K. Izumori and S. Tsuyumu, 2018. Hyperinduction of pectate lyase in Dickeya chrysanthemi EC16 by plant-derived sugars. J. Plant Interact., 13: 141-150.
CrossRefDirect Link - Flor, H.H., 1971. Current status of the gene-for-gene concept. Annu. Rev. Phytopathol., 9: 275-296.
CrossRef - Chisholm, S.T., G. Coaker, B. Day and B.J. Staskawicz, 2006. Host-microbe interaction: Shaping the evolution of the plant immune response. Cell, 124: 803-814.
CrossRefDirect Link - Han, X., L. Li, L. Cui, J. Xing, L. Tang and M. Cao, 2011. Isolation of candidate disease resistance genes from enrichment library of Oryza minuta based on conserved domains. Afr. J. Biotechnol., 10: 14738-14745.
CrossRefDirect Link - Islam, M.S., M.S. Haque, M.M. Islam, E.M. Emdad and A. Halim et al., 2012. Tools to kill: Genome of one of the most destructive plant pathogenic fungi Macrophomina phaseolina. BMC Genom., Vol. 13.
CrossRef - Solekha, R., F.A. Susanto, T. Joko, T.R. Nuringtyas and Y.A. Purwestri, 2019. Phenylalanine Ammonia Lyase (PAL) contributes to the resistance of black rice against Xanthomonas oryzae pv. oryzae. J. Plant Pathol., (In Press).
CrossRefDirect Link - Pandolfi, V., J. Neto, M. Silva, L. Amorim and A.W. Nogueira et al., 2017. Resistance (R) genes: Applications and prospects for plant biotechnology and breeding. Curr. Protein Pept. Sci., 18: 323-334.
CrossRefDirect Link - Sutrisno, F.A. Susanto, P. Wijayanti, M.D. Retnoningrum, T.R. Nuringtyas, T. Joko and Y.A. Purwestri, 2018. Screening of resistant Indonesian black rice cultivars against bacterial leaf blight. Euphytica, Vol. 214, No. 11.
CrossRefDirect Link - Quirin, E.A., H. Mann, R.S. Meyer, A. Traini, M.L. Chiusano, A. Litt and J.M. Bradeen, 2012. Evolutionary meta analysis of solanaceous resistance gene and solanum resistance gene analog sequences and a practical framework for cross-spesies comparisons. Mol. Plant Microbe-Interact., 25: 603-612.
CrossRefDirect Link - Gururani, M.A., J. Venkatesh, C.P. Upadhyaya, A. Nookaraju, S.K. Pandey and S.W. Park, 2012. Plant disease resistance genes: Current status and future directions. Physiol. Mol. Plant Pathol., 78: 51-65.
CrossRefDirect Link - Martin, G.B., S.H. Brommonschenkel, J. Chunwongse, A. Frary and M.W. Ganal et al., 1993. Map based cloning of a protein kinase gene conferring disease resis-tance in tomato. Science, 262: 1432-1436.
CrossRefDirect Link - Sekhwal, M., P. Li, I. Lam, X. Wang, S. Cloutier and F. You, 2015. Disease resistance gene analogs (RGAs) in plants. Int. J. Mol. Sci., 16: 19248-19290.
CrossRefDirect Link - Chang, J.H., Y.S. Tai, A.J. Bernal, D.T. Lavelle, B.J. Staskawicz and R.W. Michelmore, 2002. Functional analyses of the Pto resistance gene family in tomato and the identification of a minor resistance determinant in a susceptible haplotype. Mol. Plant-Microbe Interact., 15: 281-291.
CrossRefDirect Link - Vleeshouwers, V.G.A.A., A. Martens, W. van Dooijeweert, L.T. Colon, F. Govers and S. Kamoun, 2001. Ancient diversification of the Pto kinase family preceded speciation in Solanum. Mol. Plant–Microbe Interact., 14: 996-1005.
CrossRefDirect Link - Vallad, G., M. Rivkin, C. Vallejos and P. McClean, 2001. Cloning and homology modelling of a Pto-like protein kinase family of common bean (Phaseolus vulgaris L.). Theor. Appl. Genet., 103: 1046-1058.
CrossRefDirect Link - Deng, Z. and F.G. Gmitter, 2003. Cloning and characterization of receptor kinase class disease resistance gene candidates in citrus. Theor. Appl. Genet., 108: 53-61.
CrossRefDirect Link - Gaspero, G.D. and G. Cipriani, 2003. Nucleotide binding site/leucine-rich repeats, Pto-like and receptor-like kinases related to disease resistance in grapevine. Mol. Genet. Genomics, 269: 612-623.
CrossRefDirect Link - Peraza-Echeverria, S., A. James-Kay, B. Canto-Canché and E. Castillo-Castro, 2007. Structural and phylogenetic analysis of Pto-type disease resistance gene candidates in banana. Mol. Genet. Genomics, 278: 443-453.
CrossRefDirect Link - Zamora, M.G.M., A.P. Castagnaro and J.C. Díaz Ricci, 2008. Genetic diversity of Pto-like serine/threonine kinase disease resistance genes in cultivated and wild strawberries. J. Mol. Evol., 67: 211-221.
CrossRefDirect Link - Venkatesh, J., M. Jahn and B.C. Kang, 2016. Genome-wide analysis and evolution of the Pto-Like protein kinase (PLPK) gene family in pepper. PLoS ONE, Vol. 11.
CrossRefDirect Link - Elina, J., D. Sukma, Giyanto and D. Sudarsono, 2017. Isolasi dan karakterisasi gen Pto Asal 20 aksesi anggrek phalaenopsis. J. Agron. Indonesia, 45: 204-2011.
CrossRefDirect Link - Sutanto, A., D. Sukma, C. Hermanto and S. Sudarsono, 2014. Isolation and characterization of resistance gene analogue (RGA) from fusarium resistant banana cultivars. Emir. J. Food Agric., 26: 508-518.
CrossRefDirect Link - Joko, T., H. Hirata and S. Tsuyumu, 2007. A sugar transporter (MfsX) is also required by Dickeya dadantii 3937 for in planta fitness. J. Gen. Plant Pathol., 73: 274-280.
CrossRefDirect Link - Widyaningsih, S., S.N.H. Utami, T. Joko and S. Subandiyah, 2019. Plant response and huanglongbing disease development against heat treatments on 'Siam Purworejo' (Citrus nobilis (Lour)) and 'Nambangan' (C. maxima (Burm.) Merr.) under field condition. Arch. Phytopathol. Plant Protect., 52: 259-276.
CrossRefDirect Link - Trianom, B., T. Arwiyanto and T. Joko, 2019. Morphological and molecular characterization of Sumatra disease of clove in Central Java, Indonesia. Trop. Life Sci. Res., 30: 107-118.
CrossRefDirect Link - Rahma, A.A., Suryanti, S. Somowiyarjo and T. Joko, 2020. Induced disease resistance and promotion of shallot growth by Bacillus velezensis B-27. Pak. J. Biol. Sci., 23: 1113-1121.
CrossRefDirect Link - Joko, T., A. Soffan and M.S. Rohman, 2019. A novel subspecies-specific primer targeting the gyrase B gene for the detection of Pectobacterium carotovorum subsp. brasiliense. Biodiversitas: J. Biol. Divers., 20: 3042-3048.
CrossRefDirect Link - Hardie, D.G., 1999. Plant protein serine/threonine kinases: Classification and functions. Annu. Rev. Plant Physiol. Plant Mol. Biol., 50: 97-131.
CrossRefDirect Link - Kumar, S., G. Stecher and K. Tamura, 2016. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol., 33: 1870-1874.
CrossRefDirect Link - Navitasari, L., T. Joko, R.H. Murti and T. Arwiyanto, 2020. Rhizobacterial community structure in grafted tomato plants infected by Ralstonia solanacearum. Biodiversitas, 21: 4888-4895.
CrossRefDirect Link - Lehti-Shiu, M.D. and S.H. Shiu, 2012. Diversity, classification and function of the plant protein kinase superfamily. Phil. Trans. R. Soc. B, 367: 2619-2639.
CrossRefDirect Link - Afzal, A.J., A.J. Wood and D.A. Lightfoot, 2008. Plant receptor-like serine threonine kinases: Roles in signaling and plant defense. Mol. Plant Microbe Interact., 21: 507-517.
CrossRefDirect Link - Parravicini, G., C. Gessler, C. Denance, P. Lasserre-Zuber and E. Vergne et al., 2011. Identification of serine/threonine kinase and nucleotide-binding site-leucine-rich repeat (NBS-LRR) genes in the fire blight resistance quantitative trait locus of apple cultivar ‘Evereste’. Mol. Plant Pathol., 12: 493-505.
CrossRefDirect Link - Hammond-Kosack, K.E. and D.J.G. Jones, 1997. Plant disease resistance genes. Ann. Rev. Plant Physiol. Plant Mol. Biol., 48: 575-607.
CrossRef