
Ahmad Muarifin
Laboratory of Genetics and Breeding, Faculty of Biology, Universitas Gadjah Mada, Jl. Teknika Selatan, Sekip Utara, 55281, Yogyakarta, Indonesia
Ayudha Bahana Ilham Perdamaian
Laboratory of Genetics and Breeding, Faculty of Biology, Universitas Gadjah Mada, Jl. Teknika Selatan, Sekip Utara, 55281, Yogyakarta, Indonesia
LiveDNA: 62.19319
Dian Sartika
Laboratory of Genetics and Breeding, Faculty of Biology, Universitas Gadjah Mada, Jl. Teknika Selatan, Sekip Utara, 55281, Yogyakarta, Indonesia
Budi Setiadi Daryono
Laboratory of Genetics and Breeding, Faculty of Biology, Universitas Gadjah Mada, Jl. Teknika Selatan, Sekip Utara, 55281, Yogyakarta, Indonesia
LiveDNA: 62.25765
Asian Journal of Plant Sciences
Year: 2021 | Volume: 20 | Issue: 2 | Page No.: 263-270







ABSTRACT
Background and Objective: Peanut (Arachis hypogaea L.) is one of the most important crop species in Indonesia. Peanut production can be increased by genetic improvement through plant polyploidization. This research aimed to study the optimal concentration of periwinkle extract application for peanut chromosome polyploidization. Materials and Methods: Six different periwinkle leaf extract concentrations (0.01, 0.05, 0.1, 0.15, 0.2 and 0.25%) were used to induce peanut polyploidy by 12 and 24 hrs of induction. These were compared to negative (water) and positive (0.1% colchicines) controls by observing the morphological character, Squash slice and flow cytometry. Results: All treated peanuts showed improved phenotypes (i.e., higher leaf number, main stem height and pod number) than the negative control. Squash slice and flow cytometry showed that the chromosome number was doubled after 24 hrs of treatment with 0.15% periwinkle extract. Conclusion: Catharanthine content of periwinkle leaf was successful to induce polyploidization. Based on study, the optimal formula for peanut polyploidization was 0.15% of periwinkle extract after 24 hrs incubation. Treated peanut had better morphological performance i.e., higher leaf number, stem height and pod number.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Ahmad Muarifin, Ayudha Bahana Ilham Perdamaian, Dian Sartika and Budi Setiadi Daryono, 2021. Induced Polyploidy in Arachis hypogaea L. var. Talam using Catharanthus roseus Phenolic Extract. Asian Journal of Plant Sciences, 20: 263-270.
DOI: 10.3923/ajps.2021.263.270
URL: https://scialert.net/abstract/?doi=ajps.2021.263.270
DOI: 10.3923/ajps.2021.263.270
URL: https://scialert.net/abstract/?doi=ajps.2021.263.270
INTRODUCTION
Peanut (Arachis hypogaea L.) is widely used in Indonesia as a complementary food item. Although the peanut was introduced to Indonesia, it was widely adopted and grown as main food item. Recently, 34 varieties of peanut from Spain and Valencia were cultivated. Although Indonesia introduced and produced its own new peanut varieties, the old Kelinci and Gadjah variants remain popular. However, the more recent peanut varieties, such as Talam, have better agricultural performance. According to the Indonesian ministry of Agriculture, the Talam variant can yield 3.2 t ha–1 of peanut seed 90 days after planting. Furthermore, Talam was a Spanish type of peanut that is resistant to the pathogen Aspergillus flavus which the main treat for peanut1. Talam was produced by selective breeding from Giraffe (drought and acidic soil tolerant variant) to the ICGV 91283 variant (also resistant to A. flavus) at the ninth generation (i.e., F9)2.
The high demand for peanuts in Indonesia was unmatched by the supply. However, peanut production can be increased by genetics improvement through plant polyploidization. Traditionally, plant polyploidization can be conducted using commercially available colchicines powder from autumn crocus (Colchicum autumnale Linn.) extract3. This alkaloid compound causes uneven chromosome migration during cell division by altering the microtubule formation4,5. Recently, the source of colchicines was substituted by catharanthine from the periwinkle plant (Catharanthus roseus [L]. G. Don.).
The periwinkle plant is native to Madagascar. It was introduced to other tropical places including Indonesia. Periwinkle plants contain a small amount of catharanthine, which is an anti-mitotic agent similar to the colchicines in autumn crocus. Catharanthine and vindoline are sources for vinblastine and vincristine synthesis5,6.
Polyploid plants tend to have higher performance (growth rate, size, resistance etc.) than diploid plants7. Artificial polyploidization using colchicines from autumn crocus was reported to be successful in various plant species8-12. Whether catharanthine extract from periwinkle acts on peanut plants remains unknown.
Almost all varieties of peanuts are already polyploid (2n = 4x = 40) and are hybrids between different species13; including in Indonesia. Further duplication of the chromosome set (octoploid) is hypothesized to increase the harvest performance of peanuts. In this study, the cytological and phenotypic characters of the Talam peanut were investigated after the application of catharanthine from the periwinkle extract.
MATERIALS AND METHODS
Polyploidy induction: This research was conducted from October, 2017 to May, 2019 at several locations. Seed polyploidization was conducted in the Laboratory of Genetics and Breeding (Faculty of Biology, UGM). Karyotyping and ploidy analysis was conducted at the Laboratory of Clinical Pathology (Faculty of Medicine, Public Health and Nurse, Gadjah Mada University). Peanut seeds were provided by the Department of Forestry and Plantation office (Balai Pengembangan Perbenihan dan Percontohan Kehutanan dan Perkebunan; BP3KP), Yogyakarta Special Province. The peanuts were sorted by weight to minimize false positives. Peanuts were homogenized by weight (0.45-0.53 g) and were divided into eight groups each consisting of five seeds.
Six different concentrations of periwinkle (Catharanthus roseus [L]. G. Don.) leaf extracts (i.e., 0.01, 0.05, 0.1, 0.15, 0.2 and 0.25%) were used to assess peanut polyploidy after 12 or 24 hrs of submerging. 0.1% Colchicin BioChemica (PanReac AppliChem GmbH; German) was used as the positive control whereas dH2O was used as the negative control.
The seeds were germinated and cultivated in the greenhouse until harvest. The peanuts were provided with adequate water and fertilizers following standard peanut cultivation protocols.
Peanut growing: Germination of peanuts was conducted in a greenhouse at Inter-University Center (Pusat Antar Universitas; PAU; UGM). The seedlings were placed in small polybag (10×15 cm) until 3 weeks of age. The plants were then transferred into a big polybag (50×40 cm) and moved outside of the greenhouse for optimal capture of sunlight.
Phenotyping: At the harvesting age, the height of the main stem of plants and the leaf number was measured. The pods of each plant were collected and counted.
Qualitative and quantitative analysis of peanut: After harvest, the young roots and leaves of plants from each treatment group were collected for qualitative (squash) and quantitative (flow cytometry) analyzes. For the qualitative analysis, small sections of root were pre-fixed using 0.03% colchicine prior to fixation using 45% glacial acetic acid. The fixated leaves were softened using 1N HCl were subsequently stained using 1% aceto orcein. The leaf material was mounted and observed under a light microscope.
For the quantitative analysis, the midrib of each leaf was cut from leaves and discarded. The leaf material then placed into sterile microtubes and treated with 45% glacial acetic acid at 4°C for 10 min before washing three times with ddH2O. This was followed by softening by HCl 1 N at 60°C for 15 min before washing three times again with ddH2O. Otto buffer was added into each tube and was subsequently ripped by scalpel. Gradual filtration through 100, 50 and 10 μm filter paper was performed to eliminate cell debris. The filtrate was stained using propidium iodide (Pi) to visualize the nuclei. The filtrate was then transferred into the tube for the flow cytometry Section (Dickinson FACSCalibur). Nuclei were identified as a thickening of light intensity. The light was captured by a detector and then the curves and peaks were processed.
Data analysis: All acquired data from periwinkle treatment were compared to the positive and negative controls using Duncan multiple range test and LSD using SPSS.’’
RESULTS AND DISCUSSION
Phenotype of peanut: In groups of peanuts treated with periwinkle extract, Fig. 1 shows that the average plant height was higher than control. The tallest plant (21.5 cm) was archived by 24 hrs treatment of 0.1% periwinkle extract (Fig. 1). In general, all treated seeds had higher yields than both positive and negative controls. Only the 0.01% treatment group did not show statistical difference to the control.
Based on recent results (Fig. 2), the second highest leaf number (24.33 pieces) was obtained with 12 hrs of 0.15% periwinkle extract treatment. The highest number was 12 hrs of 0.1% catharanthine. The table implies that all treated group yield more leaf number than control group.
Based on recent results (Fig. 3), the periwinkle extract treated plants yielded more pods than the negative control, but only the 0.2 and 2.5% treatments were statically higher than the positive control. Negative control peanut (without colchicines or catharanthine) yielded 17 pods (Fig. 3a), positive control peanut (0.1% colchicines) yielded 25 pod (Fig. 3b), treated peanut with 0.01% periwinkle extract yielded 25 pods (Fig. 3c), treated peanut with 0.05% periwinkle extract yielded 19 pods (Fig. 3d), treated peanut with 0.1% periwinkle extract yielded 19 pods (Fig. 3e), treated peanut with 0.15% periwinkle extract yielded 27 pods (Fig. 3f), 0.2% periwinkle extract yielded 30 pods (Fig. 3g): 0.25% of periwinkle extract yielded 33 pods (Fig. 3h). The highest pod number, 33 pods, was obtained under 0.25% of periwinkle extract treatment while the lowest pod number, 17 pods, was obtained at negative control.
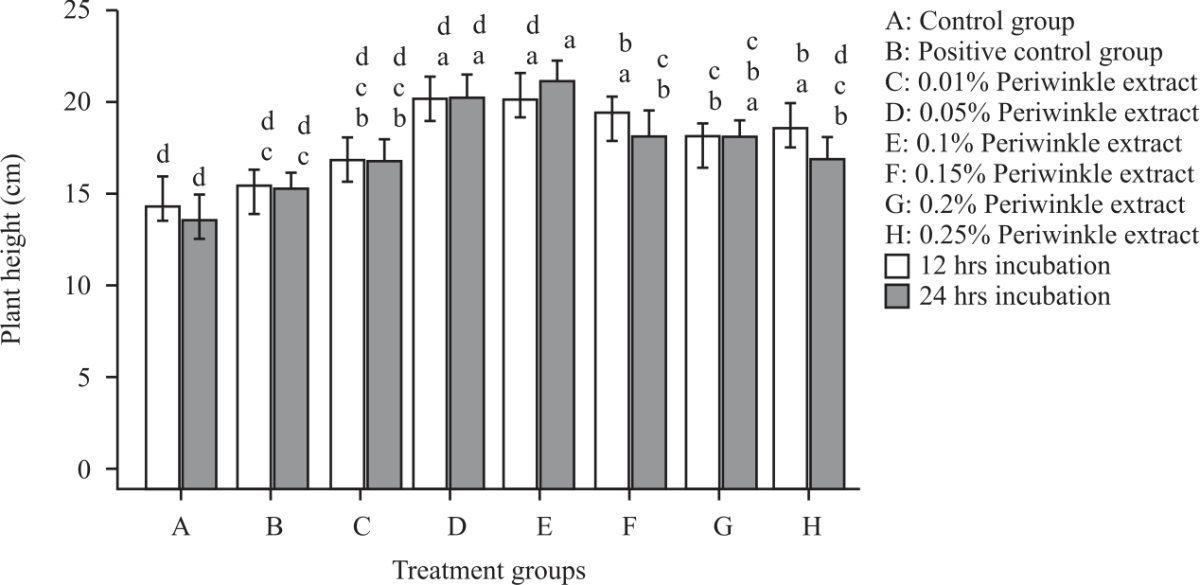 |
| Fig. 1: | Average of plant height (n = 3) at three weeks of age for each treatment group Light bars indicate 12 hrs of treatment and the dark bars indicate 24 hrs of treatment. Bars represented by the same letters are not statistically significant (LSD, p<0.05) |
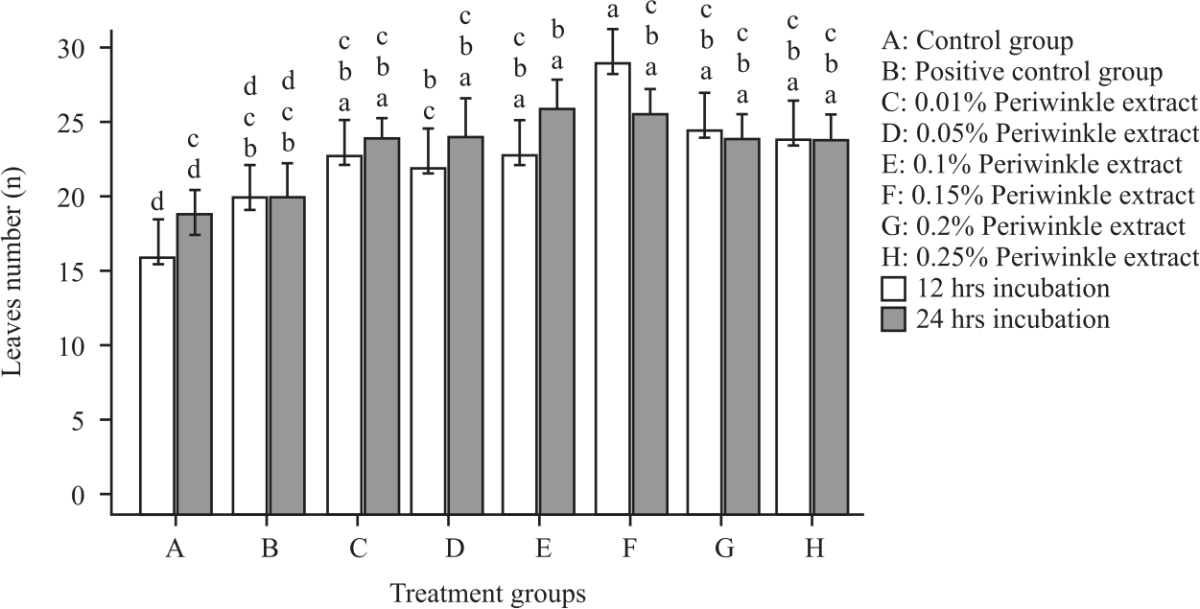 |
| Fig. 2: | Average number of peanut plant leaves (n = 3) at three weeks of age Light bars indicate 12 hrs of treatment and the dark bars indicate 24 hrs of treatment. Bars represented by the same letters are not statistically significant (LSD, p<0.05) |
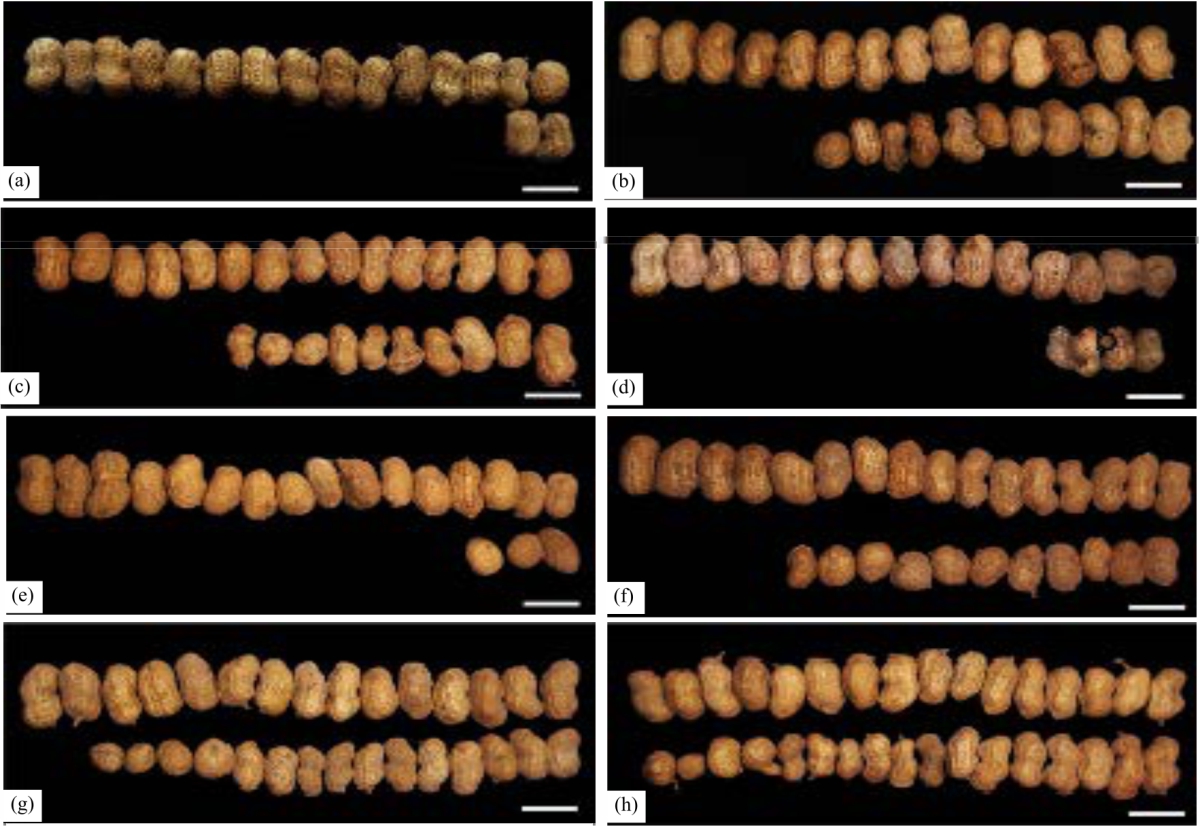 |
| Fig. 3: | Average pod number and the weight of pods and the seed of treated peanut compared to the positive and negative controls |
Here, all treated peanut plants produced better phenotypes than the negative control. In contrast, the polyploidization experiments conducted previously on Miscanthus14,15, Glycyrrhiza9, rubber12, willow16, Dendrobium17, Stevia18 and kiwi19 plants all yielded higher output for the polypoid forms compared to their diploid base forms. Those plants were treated using conventional colchicines and there was limited reference to periwinkle extracts.
This result might form the higher number of chromosomes in cell rise the activity of beneficial genes. The results indicate that highest concentration of catharanthine (0.25%) not yield highest leaf number. This might be due to the toxicity from the catharanthine to the cell. This results not consisted with previous work with watermelon15 which at 0.25%. The results regarding the duration of treatment were not consistent across the treatment groups. The results were similar to previous studies which polyploid plant was larger than its diploid13,14.
A limitation of recent research was that the use of polybags is not optimal for the growth of peanut plants, as the gynophore of the peanut plants may not develop well.
Karyotype analysis of peanut: The root tips of plants from each treatment group were collected for cytological analysis. All samples underwent pre-fixation using 0.03% colchicines for 24 hrs at 14°C. This step was important to break the cytoskeleton in the cells thus the chromosome is distributed evenly in the cell. This makes for easier counting under the light microscope (Fig. 4).
Under the light microscope, the nuclei were observed and the chromosomes were counted. Of each group decided in most consistent result through perpetrates. During prometaphase, the chromosomes were easier to be observed because they are most highly condensed.
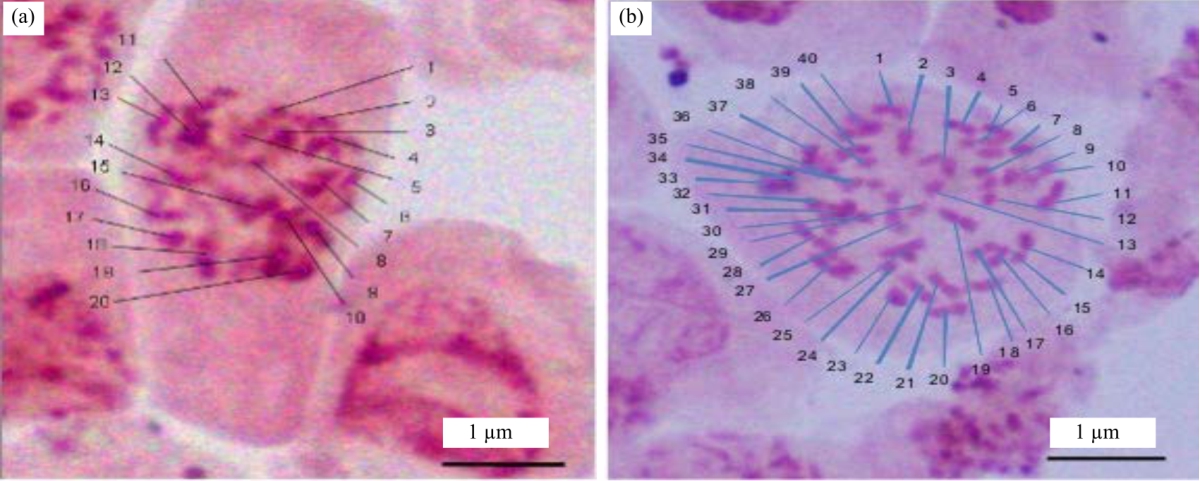 |
| Fig. 4(a-b): | (a) Prometaphase of negative control contains 20 pair of chromosomes and (b) 0.1% periwinkle extract treated peanut root cell contains 40 pair of chromosomes 400×magnifications was used |
Based on the microscope slice from the squashing method, 0.1% periwinkle extract treatment successfully yielded octoploid plants (2n = 8x) (Fig. 4a) which had a two-fold higher number of chromosomes compared to the negative control (2n = 4x) (Fig. 4b). Natural polyploidization was noticed in several crop plants including tetraploid potato (2n = 4x), hexaploids wheat (2n = 6x) and octoploid sugar cane (2n = 8x)20. However, peanut plants produced from the hybridization of two or more species can also undergo chromosome polarization, resulting in allotetraploid plants (2n = 4x = 2A+2B = 40)21.
Flow cytometry (FCM) analysis: Periwinkle extract-treated and control peanut leaves were subjected to Flow cytometry (FCM) quantitative analysis. To date, FCM is standard tool for the estimation of the amount of plant DNA16,17,22. Histograms of each treatment group are presented in Fig. 5. In histogram, cell numbers depicted at y-axis while fluorochrome intensity depicted at x-axis. The position of cell number peak at x-axis (FL2-Area) can determine the degree of ploidy. The peak of octoploid plant’s G2/M always occurs and located two-fold away from the peak of tetraploid plant’s G0/G1.
The histogram of the control group (Fig. 5a) shows a single peak of cell numbers at y-axis at 100-200 scale of x-axis (FL2-Area) indicating most of cell at the interphase (G0/G1) and approximately 1251 cells were counted from the sample injected to the machines (CV 20.30%). Positive control peanut (Fig. 5b) also at interphase (G0/G1) and 3670 cells were counted (CV 18.88%) (Fig. 5b). The position of positive control’s peak was similar to the negative control’s peak. The flow cytometry (FCM) outputs indicate that both negative and positive controls had two sets of chromosomes (2n=4x).
Peanut seeds treated with 0.15% periwinkle extracts had two peaks at interphase (G0/G1) and mitotic phase (G2/M) (Fig. 5c). From the FCM data, 864 cells were counted at G0/G1 (CV 19.01) and 1111 cells were counted at G2/M (CV 10.96%). The G2/M’s peak was occurred and distant away from G0/G1 indicating that the chromosome set become octoploid (2n = 8x). However, the cell read was very low and the CV was >10%. Conversely, 0.2% periwinkle extract treated peanut had single peak which counted 2047 cells at interphase (G0/G1) (CV 20.93) (Fig. 5d). The peak of 0.2% periwinkle extract treated peanut had similar position to negative control indicated that plant had diploid (2n) at interphase (G0/G1). Furthermore, the 0.25% periwinkle extract treated peanut showed higher cell counts at the G2M phase, 712 than the G0/G1 phase (314) (CV 22.71%); however, a stationary peak was produced. Thus, this histogram cannot be interpreted as octoploid (2n = 8x) (Fig. 5e).
Based on the FCM data, the octoploid plant (2n = 8x) was clearly observed at 0.15% periwinkle extract. Not all treated peanut plants produced readable FCM data. This may be due to some errors in sample preparation. Nonetheless, the performance of peanut plants was better than the negative control. In this research, the widely available periwinkle is promising as a polyploidization agent. Further research is needed to confirm this effect on field plants. Beside catharanthine as anti-mitotic agents, other attempt using oryzalin to produce polyploid plant was successful in watermelon23.
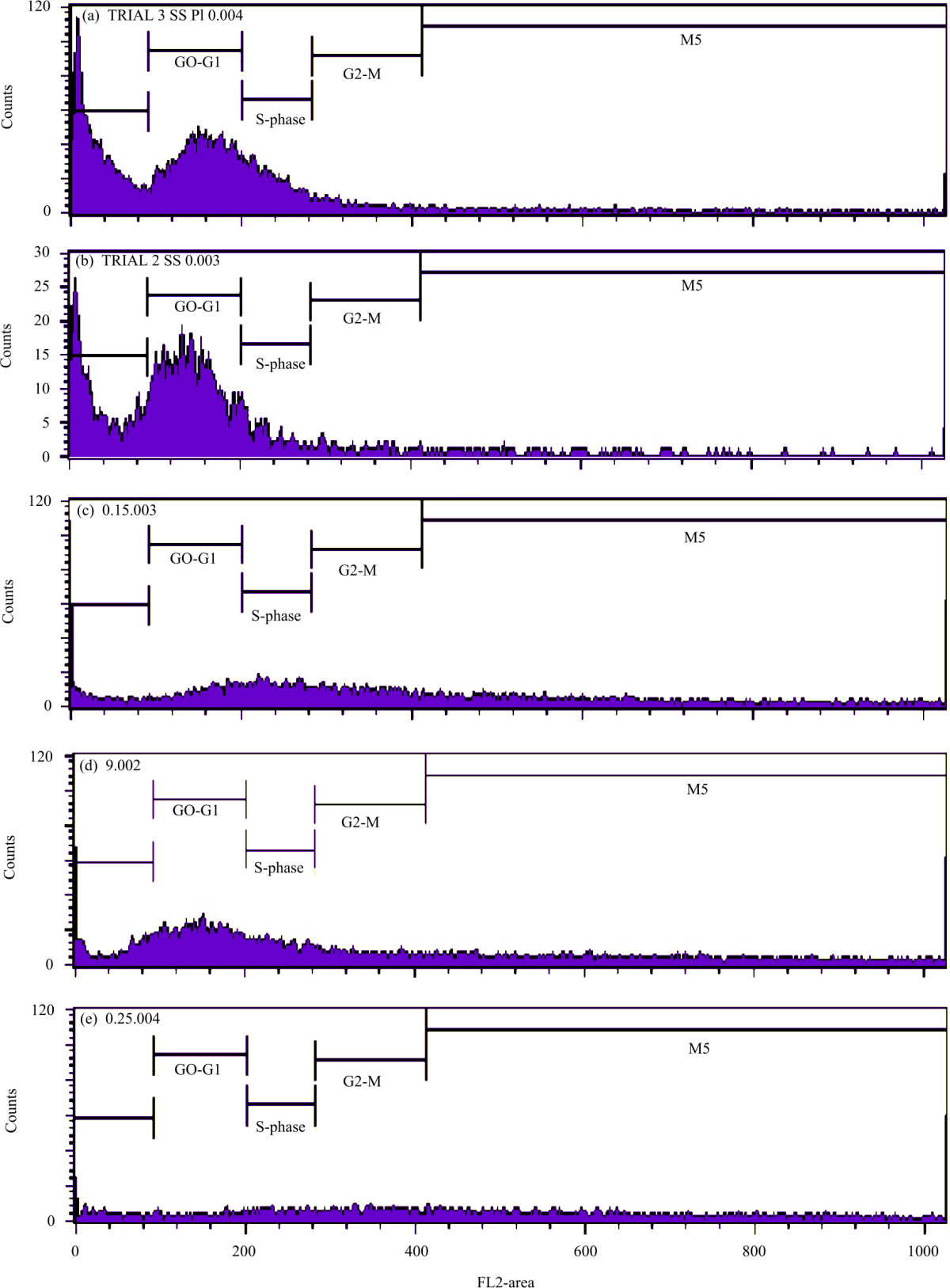 |
| Fig. 5(a-e): | Representative DNA histograms of cell count (Y axis) against FL2-A (FL2-Area: total DNA fluorescence) (X axis) for propidium iodide-stained peanuts cells, (a) Negative control, (b) Positive control, (c) 0.15% periwinkle extract treated group, (d) 0.2% periwinkle extract treated group and (e) 0.25% periwinkle extract treated group X axis indicate FL2-Area (Fluorescence detector number 2 Area) whereas Y axis, courts indicate the cell numbers. G0-G1: Growth phase 0-Growth phase 1 of interphase, S-phase: Synthesis phase, G2-M: Growth phase 2-Mitotic phase |
CONCLUSION
Based on the results of this research, any level of periwinkle extract application can boost the performance of peanut plants. The squash slice method and FCM showed that the chromosome number was doubled and the highest read was obtained after 24 hrs of treatment using 0.15% periwinkle extracts.
SIGNIFICANCE STATEMENT
The present study demonstrated that Madagascar periwinkle extractable to doubled peanut chromosomes. Polyploidization significantly improved shoot growth and peanut yield of the peanut plant. Accordingly, the results of this study may be useful for the development of peanut plant cultivars.
ACKNOWLEDGMENT
The authors want to acknowledge Universitas Gadjah Mada for Providing RTA research grant (No. 1655/UN1/ DITLIT/DIT-LIT/PT/2020). The authors want to thank Agus Setiawan, S.Si. for the valuable advice and technical assistance in chromosome preparation. The authors want to thank Mr. Farid for his assistance in operating the flow cytometry instrument in the laboratory.
REFERENCES
- Jayaprakash, A., R.R. Thanmalagan, A. Roy, A. Arunachalam and P.T.V. Lakshmi, 2019. Strategies to understand Aspergillus flavus resistance mechanism in Arachis hypogaea L. Curr. Plant Biol., Vol. 20.
CrossRefDirect Link - Purnomo, P. and N. Khotimah, 2019. Variations and phenetic analysis of peanut cultivars (Arachis hypogaea L.) based on morphological characteristics. J. Trop. Biodivers. Biotechnol., 4: 24-31.
CrossRefDirect Link - Castro, M., S. Castro and J. Loureiro, 2018. Production of synthetic tetraploids as a tool for polyploid research. Web Ecol., 18: 129-141.
CrossRefDirect Link - Spasevska, I., A. Ayoub, P. Winter, J. Preto, G. Wong, C. Dumontet and J. Tuszynski, 2017. Modeling the Colchicum autumnale tubulin and a comparison of its interaction with colchicine to human tubulin. Int. J. Mol. Sci., Vol. 18.
CrossRefDirect Link - S.J. Pyle and K.R. Reuhl, 2010. Cytoskeletal elements in neurotoxicity*. Compr. Toxicol., Sec. Ed., 13: 71-87.
CrossRefDirect Link - Ishikawa, H., D.A. Colby and D.L. Boger, 2008. Direct coupling of catharanthine and vindoline to provide vinblastine: Total synthesis of (+)- and ent-(−)-vinblastine. J. Am. Chem. Soc., 130: 420-421.
CrossRefDirect Link - Corneillie, S., N. De Storme, R.V. Acker, J.U. Fangel and M. De Bruyne et al., 2019. Polyploidy affects plant growth and alters cell wall composition. Plant Physiol., 179: 74-87.
CrossRefDirect Link - Glowacka, K., S. Jezowski and Z. Kaczmarek, 2009. Polyploidization of Miscanthus sinensis and Miscanthus x giganteus by plant colchicine treatment. Ind. Crops Prod., 30: 444-446.
CrossRefDirect Link - Moghbel, N., M.K. Borujeni and F. Bernard, 2015. Colchicine effect on the DNA content and stomata size of Glycyrrhiza glabra var. glandulifera and Carthamus tinctorius L. cultured in vitro. J. Genet. Eng. Biotechnol., 13: 1-6.
CrossRefDirect Link - Munzbergora, Z., 2017. Colchicine application significantly affects plant performance in the second generation of synthetic polyploids and its effects vary between populations. Ann. Bot., 120: 329-339.
CrossRefDirect Link - Głowacka, K., S. Jeżowski and Z. Kaczmarek, 2010. In vitro induction of polyploidy by colchicine treatment of shoots and preliminary characterisation of induced polyploids in two Miscanthus species. Ind. Crops Prod., 32: 88-96.
CrossRefDirect Link - Luo, Z., B.J. Iaffaldano and K. Cornish. 2018. Colchicine-induced polyploidy has the potential to improve rubber yield in Taraxacum kok-saghyz. Ind. Crops Prod., 112: 75-81.
CrossRefDirect Link - Bertioli, D.J., J. Jenkins, J. Clevenger, O. Dudchenko and D. Gao et al., 2019. The genome sequence of segmental allotetraploid peanut Arachis hypogaea. Nat. Genet., 51: 877-884.
CrossRefDirect Link - Zhuang, W., H. Chen, M. Yang, J. Wang and M.K. Pandey et al., 2019. The genome of cultivated peanut provides insight into legume karyotypes, polyploid evolution and crop domestication. Nat. Genet., 51: 865-876.
CrossRefDirect Link - Bae, S.J., M.M. Islam, H.Y. Kim and K.B. Lim, 2020. Induction of tetraploidy in watermelon with oryzalin treatments. Hortic. Sci. Technol., 38: 385-393.
CrossRefDirect Link - Guo, W., J. Hou, T. Yin and Y. Chen, 2016. An analytical toolkit for polyploid willow discrimination. Sci. Rep., Vol. 6.
CrossRefDirect Link - Grosso, V., A. Farina, D. Giorgi, L. Nardi, G. Diretto and S. Lucretti, 2018. A high-throughput flow cytometry system for early screening of in vitro made polyploids in Dendrobium hybrids. Plant Cell Tiss. Organ. Cult., 132: 57-70.
CrossRefDirect Link - Zhang, H., S. An, J. Hu, Z. Lin, X. Liu, H. Bao and R. Chen, 2018. Induction, identification and characterization of polyploidy in Stevia rebaudiana Bertoni. Plant Biotechnol., 35: 81-86.
CrossRefDirect Link - Wu, J.H., A.R. Ferguson, B.G. Murray, Y. Jia, P.M. Datson and J. Zhang, 2012. Induced polyploidy dramatically increases the size and alters the shape of fruit in Actinidia chinensis. Ann. Bot., 109: 169-179.
CrossRefDirect Link - Sattler, M.C., C.R. Carvalho and W.R. Clarindo, 2016. The polyploidy and its key role in plant breeding. Planta, 243: 281-296.
CrossRefDirect Link - Yin, D., C. Ji, X. Ma, H. Li and W. Zhang et al., 2018. Genome of an allotetraploid wild peanut Arachis monticola: A de novo assembly. Giga Sci., Vol. 7.
CrossRefDirect Link - Varela-Álvarez, E., J. Loureiro, C. Paulino and E.A. Serrão, 2018. Polyploid lineages in the genus Porphyra. Sci. Rep., Vol. 8.
CrossRefDirect Link - Bae, S.J., M.M. Islam, H.Y. Kim and K.B. Lim, 2020. Induction of tetraploidy in watermelon with oryzalin treatments. Hortic. Sci. Technol., 38: 385-393.
CrossRefDirect Link