
Ahemd Farahat Sahab
Department of Plant Pathology, National Research Centre, Dokki, Giza, Egypt
Ayman Anter Saber Abdullah
Department of Field Crop Research, National Research Centre, Dokki, Giza, Egypt
LiveDNA: 20.33544
Ghada Mansour Samaha
Department of Field Crop Research, National Research Centre, Dokki, Giza, Egypt
El-Sead Husein Ziedan
Department of Plant Pathology, National Research Centre, Dokki, Giza, Egypt
Asian Journal of Plant Sciences
Year: 2021 | Volume: 20 | Issue: 2 | Page No.: 220-231







ABSTRACT
Background and Objective: Sesame diseases, which are mostly caused by fungal pathogens, are important production restrictions in Egypt and climatic change is exacerbating their impact. The objectives of this study were to identify high yield lines, to find out the phytochemical components in the ethanol extracts of genotypes and to determine some markers related to genes controlling the Fusarium wilt resistance trait. Materials and Methods: Four elite sesame lines and commercial variety Shandaweel were evaluated based on seed yield ha–1 over two seasons. And they tested for seed-borne fungi. Then these genotypes were examined using R-ISSR. Results: Significant variations (p<0.05) found among genotypes for seed yield ha–1. The highest yield was obtained from Shandaweel. The line C3.8 recorded the low fungal incidence and achieved the highest germination and its seed has chemical compositions that have antifungal activity and absent in Shandaweel. R-ISSR generated more markers linked to Fusarium wilt resistance compared with RAPD and ISSR when used alone. Line C3.8 contains defense response genes against fungi diseases, which produced bioactive chemical compounds that have conferred resistance. Conclusion: Line C3.8 will be used as a parent in other breeding programs to form new varieties resistant to Fusarium wilt in sesame. And, this line can use as a basis for generating new broad-spectrum antimicrobial formulations.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Ahemd Farahat Sahab, Ayman Anter Saber Abdullah, Ghada Mansour Samaha and El-Sead Husein Ziedan, 2021. Evaluation of Sesame Genotypes for Yield, Phytochemical Characteristics and Fusarium Wilt Resistance by R-ISSR. Asian Journal of Plant Sciences, 20: 220-231.
DOI: 10.3923/ajps.2021.220.231
URL: https://scialert.net/abstract/?doi=ajps.2021.220.231
DOI: 10.3923/ajps.2021.220.231
URL: https://scialert.net/abstract/?doi=ajps.2021.220.231
INTRODUCTION
Sesame (Sesamum indicum L.) crop is recognized as the talent of oil seeds owing to its high value oil quality and excess of nutritive substances1. Conventional, sesame seeds are used for a variety of purposes including making margarine, sweets, cakes and bread. It acts as one of the ingredients for the manufacture of soaps, perfumes, pharmaceutical products and insecticides2. In addition, sesame plant has many characterizes including adaptability to a domain of soil types, drought tolerant, oil content reached to 63%, oil has a phenolic group is responsible for antioxidant activities, source of super quality oil, resistance against oxidation3-5. Despite these important characteristics, the area planted with sesame in Egypt is still insufficient compared to other crops, which contributes to low production capacity and instability6. The cultivated area of sesame crop reached 34,000 hectares and produced 44,000 t and the unit productivity is 1294.1 kg ha–1 7. Sesame Fusarium wilt which is one of the main worldwide diseases in sesame, resulting in 15-30% losses of yield8. Moreover, climate change is likely to increase demand for new crop varieties more resistance to plant diseases. Because, plant diseases have been caused significant crop losses in agriculture ranged from 20-40% of crop yields worldwide and are one of the most obstacles to achieving global food security in the 21st century9-11. In addition, human used chemicals substances (fungicides) to address plant fungi diseases, but the excessive usage of these substances caused their accumulation, which affected the crop productivity and soil degradation. And thus increase the harmful effect of climate changes.
Plant diseases are caused by kinds of organisms, namely fungi, bacteria and viruses12,13. Fortunately, the seed kernels of sesame oil extracted with ethanol by GC-MS revealed the presence of many bioactive compounds, including, steroids, terpenoids, phenolic compounds, fatty acids and different types of ester14-16. And these compounds sets several biochemical reactions to the production of phytoalexin, which are antibiotic polyphenolic compositions17. The development of crop varieties that combine resistance to plant pathogens and high yield is an essential method of disease control as well as addressing the adverse effects of climate change18,19. By preventing the spread of fungal diseases, reducing the use of pesticides, reducing production costs and preserving the pollution environment.
To accelerate these tasks, molecular markers will use to identify polymorphism represented by differences in DNA sequences20. Of these markers, RAPD and ISSR markers have been used to detect markers associated with resistance to Fusarium wilt disease in many crops such as chickpea and faba bean21,22. Moreover, R-ISSR markers are useful to detect new genomic loci because they represent two different kinds of information about genomic sequences23. The current study was based on four axes including: evaluation of distinct lines based on the seed yield, evaluation based on fungal diseases resistant, the detection of some antibiotic polyphenolic compositions in genotypes and determine molecular markers linked resistance to Fusarium wilt to identify high yielding lines and to find out the phytochemical components in the ethanol extracts of genotypes by GC-MS analysis and determine molecular markers linked resistance to Fusarium wilt in new sesame lines.
MATERIALS AND METHODS
Study area: Genotypes evaluated based on seed yield ha–1 at Agricultural Production and Research Station, National Research Centre, Al-Nubaria district, El-Behera Governorate for two years (2017 and 2018) in sandy soil. Irrigation drip was applied. The research was conducted in May until September of each year.
Breeding materials: Breeding materials used in this investigation were 4 elite derived lines of sesame in F9 and F10 generations, namely; C1.5, C1.6, C3.8 and C6.3 obtained via pedigree selection from Cairo University and commercial variety, Shandaweel, (C) obtained Agriculture Research centre. The characterization of their parents and C is presented in Table 1.
Experiment design: Genotypes laid down in randomized complete blocks design with three replicates. Plots consist of 2 rows 5 m long and spaced 0.60 m apart with a 10 cm plant distanced. Seed yield ha–1 of the genotypes was taken off from net area (6 m2) of the three replications. Genotypes were grown according to the standard agricultural practice. Data of each trait were subjected to a regular analysis of variance of RCBD according to John24. The p<0.05 was considered statistically significant.
Diseases studies
Testing procedure: Seeds of all lines and C were analyzed for their association of seed-borne fungi by agar plate method25. One hundred seeds of sesame lines and C were sterilized with 1% sodium hypochlorite solution for 3 min and then placed at the rate of 10 seeds per Petri plate containing 20 mL of potato dextrose agar medium (PDA). The Petri plates were incubated for seven days in the incubator at 27±2°C and the seeds were examined individually under a dissecting microscope.
| Table 1: Origin breeding status and description for parents and C | |||||
| Genotypes | Breeding status | Seed source* | Specific characters | ||
| P1 (HM19) | F8-hybrid pop | Cairo Univ.* | Early maturity, non branching, first capsule set low, 3 capsules/axil, high resistant against Fusarium oxysporum | ||
| P2 (EUL90) | Mutant line | Cairo Univ.* | Early maturity, non-branching, first capsule set low, 3 capsule/axil, moderate resistant against Fusarium oxysporum | ||
| P3 (Mutant 48) | Mutant line | Cairo Univ.* | Branching, 3 capsules/axil, high susceptible against Fusarium oxysporum | ||
| P4 (Giza 32) | Local variety | Ministry of Agric. and Land Reclamation, Egypt | Heavy seed weight, medium branching, one capsule/axil, long capsule, late maturity, moderate resistant against Fusarium oxysporum | ||
| P5 (NM59) | Exotic line | India through IAEA** | Stiff stem, late maturity, one capsule/axil, resistant against Fusarium oxysporum | ||
| P6 (Babil) | Exotic variety | Iraq through IAEA** | Low branching, 3 capsules/axil, semi-shattering capsules, resistant against Fusarium oxysporum | ||
| C | Local variety | Ministry of Agric. and Land Reclamation, Egypt | Heavy seed weight, medium branching, three capsule/axil, long capsule, susceptible against Fusarium oxysporum | ||
| Advanced breeding materials resulted from the breeding program conducted at Agronomy Department, Faculty of Agriculture, Cairo University. **International Atomic Energy Agency. Lines C1.5 and C1.6 resulted from a hybrid P1* P2, line C3.8 resulted from a hybrid P1*P4 and line C6.5 resulted from a hybrid P2*P3 | |||||
The percentage of seed infection (PI), relative occurrence (%) (RO) of fungal genera and species on seeds were calculated as follows:
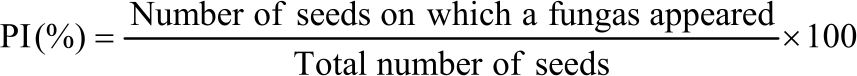 |
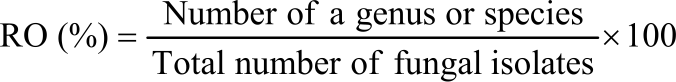 |
The total fungal counts and the frequency occurrence of different fungi associated seeds were determined. All fungal isolates of none and sterilized seeds were identified to the generic or species level according to Samson et al.26. Seeds were allowed to germinate for 14 days.
Extraction and purification of sesame oil: The 100 g of dried seeds (from lines and C) were pulverized in a heavy-duty grinder to obtain crowed powder and extracted with 95% ethanol (300 mL) using the Soxhlet apparatus. The ethanolic extract was purified by suspending 2 g of activated charcoal, just boiled and filtered and the solvent was removed by evaporation under reduced pressure and temperature. A small portion (1 g) of the extract subjected to GC/MS analysis.
Phytochemical analysis of sesame oil: The GC-MS analysis of the essential oil samples was carried out using gas chromatography-mass spectrometry instrument stands at National Research Centre with the following specifications, instrument: a TRACE GC Ultra Gas Chromatographs (THERMO Scientific Corp., USA), coupled with a THERMO mass spectrometer detector (ISQ Single Quadruple Mass Spectrometer). The GC-MS system was equipped with a TG-WAX MS column (30 m×0.25 mm i.d., 0.25 μm film thicknesses). The analysis was carried out using helium as carrier gas at a flow rate of 1.0 mL min–1 and a split ratio of 1:10 using the following temperature program: 40°C for 1 min; rising at 4.0°C min–1 to 160°C and held for 6 min; rising at 6°C min–1 to 210°C and held for 1 min. The injector and detector were held at 210°C. Diluted samples (1:10 hexane, v/v) of 0.2 μL of the mixtures always injected. Mass spectra were obtained by electron ionization (EI) at 70 eV, using a spectral range of m/z 40-450. Components preliminary identified by comparison of their mass spectra with those of a computer library or with authentic compounds and confirmed by comparison of their retention indices, either with those of authentic compounds or with data published in the literature. The retention indices calculated for all volatile constituents using a homologous series of alkanes27.
Molecular analysis
Genomic DNA extraction: Genomic DNA was extracted from genotypes using the DNeasy Plant Mini Kit (Qiagen) according to the manufacturer’s instructions. The DNA isolated checked on a 1% agarose gel.
R-ISSR analysis: Ten R-ISSR combinations (RAPD and ISSR) used for the molecular analysis and were synthesized by Sigma Aldrich, India (Table 2).
PCR reactions were performed according to Williams et al.28 in DNA thermal cycler. The reaction mixture (25 μL) contained a 2.5 μL of 10×PCR buffer, 0.5 μL of dNTPs (2.5 mM), a 2.5 μL of MgCl2 (2.5 mM), 2.0 μL of primer (RAPD+ISSR), 1 U of Taq DNA polymerase enzyme (Fermentasi) and 2 μL of DNA template. The PCR reactions were performed for R-ISSR under the following conditions: Initial denaturation at 94°C for 4 min; 35 cycles of 1 min denaturation at 94°C; 1min annealing at 40°C and extension primer at 72°C for 2 min and 10 min at 72°C for the final product extension.
| Table 2: List of R-ISSR combinations, sequences and annealing temperature | |||||
| R-ISSR | Primer name | Sequence | Annealing temperature (°C) | ||
| 1 | OP-A10+UBC-811 | GTGATCGCAG+(GA)8C | 40 | ||
| 2 | OP-A10+UBC-825 | GTGATCGCAG+(AC)8T | 40 | ||
| 3 | OP-A10+UBC-855 | GTGATCGCAG+(AC)8YT | 40 | ||
| 4 | OP-A10+UBC-864 | GTGATCGCAG+(ATG)6 | 40 | ||
| 5 | OP-A10+UBC-868 | GTGATCGCAG+(GAA)6 | 40 | ||
| 6 | OP-B05+UBC-811 | TGCGCCCTTC+(GA)8C | 40 | ||
| 7 | OP-B05+UBC -825 | TGCGCCCTTC+(AC)8T | 40 | ||
| 8 | OP-B05+UBC-855 | TGCGCCCTTC+(AC)8YT | 40 | ||
| 9 | OP-B05+UBC-864 | TGCGCCCTTC+(ATG)6 | 40 | ||
| 10 | OP-B05+UBC-868 | TGCGCCCTTC+(GAA)6 | 40 | ||
| ISSR: Inter simple sequence repeat polymorphism | |||||
The amplification product was separated by electrophoresis on agarose (1.5%) in 1×TBE buffer run for one hour at 100 V and stained with ethidium bromide. The DNA bands in the gel were observed under UV transilluminator filter. The bands were photographed using a digital camera. Solis BioDyne 100 bp DNA Ladder (100-3000 bp) was used as a size marker.
Data analysis: Amplified fragments were considered as a binary character for the present (scored 1) and absent (Scored 0). Similarities were estimated by Jaccard's coefficient. Cluster analysis was carried out with NTSYS-pc software, UPGMA algorithm29.
RESULTS
Variance and mean performance: The analysis of variance manifested a statistically significant (p<0.05) among the genotypes for seed yield ha–1. In this study, C and line C1.5 had the highest seed yield ha–1 compared to other lines in F8 and F9 generations.
Diseases studies
Prevalence of seed-borne fungi: Data presented in Table 4 showed that all Surfaces Sterilized (SS) and non (NS) of genotypes were contaminated with fungi.
The percentage of fungal infection was higher in sterilized seeds of C (68.13%), C6.3 (64.29%) and C1.5 (52.32%) and low for C1.6 (25.0%) while a line C3.8 was zero. On the other hand, the percentage of fungal infection of the non sterilized seeds was higher in C6.3 (85.71%), C1.5 (78.75%) and C (78.75%) and low in C3.8 (96.43%). Concerning the germination test, data in Table 4 showed that there was a difference in seed germination of genotypes, seed germination ranged from 35.79-100% in sterilized seeds and between 14.29-75.00% in non sterilized seeds. Of the seed genotypes that germinated in high rates occurred in sterilized seeds of C3.8 recorded 100% and C recorded 81.8%. And the same trend occurred in non-sterilized seeds of C3.8 recorded 75.0% and C recorded 64.38%.
Total count and frequency occurrence of fungi: It is clear from Table 5 that the number of fungi as cfu per 100 seeds on the PDA medium was ranged from 28.6 for line C3.8 to 471.4 for C. Seeds of C3.8 showed low fungal densities compared to other lines and C. A total of 16 fungal species were isolated from seeds of genotypes. The common genera were Aspergillus (five species), Fusarium (three species) and Penicillium (two spp.). Other genera were represented by a single species. Aspergillus flavus, A. niger, Fusarium oxysporum and Rhizopus nigricans showed the highest incidence on the sesame seeds with a mean frequency occurrence of 10.6, 18.1, 12.3 and 15.3%, respectively. Among the five species of Aspergillus reported, A. niger and A. flavus showed the highest incidence. Three Fusarium species were identified, among this F. oxysporum was found on seeds of C with high frequency reaching 18.2% while seeds of C3.8 were free of this fungus and M. phaseolina was not detected on seeds of C3.8. Accordingly, it could be considered that seeds of C3.8 were the most resistant ones for charcoal rot and wilt diseases caused by M. phaseolina and F. oxysporum. Data also showed that fungi were much more abundant in seeds of C since the seeds contained a large number of fungi reaching 15 species in comparison with seeds of line C3.8 (only two species).
Identification and quantification of the constituent of C3.8 seeds: On the basis of chemical test, crude ethanolic extract of C3.8 seeds contains numerous polar and nonpolar bioactive compounds including steroids, flavonoids, terpenoids and different types of ester compounds. The chromatogram obtained by ethanolic extract of sesame seeds is shown in Table 6. The main constituent found in C3.8 seeds were heptenoic acid, α-naphthoflavone (14.9%), hexadecanoic acid C, C1.5, C3.8, C6.3 and C1.6: Genotypes, X: lines mean, SS: Surface sterilized, Non: Non-surface sterilized-tests were rune in triplicates.
| Table 3: Mean performance and variance of genotypes for seed yield ha–1 in F9 and F10 generations | |||||
| Line | F9 | F10 | X | ||
| C | 846.4 | 856.0 | 851.2 | ||
| C1.5 | 820.3 | 864.2 | 842.3 | ||
| C3.8 | 652.3 | 715.0 | 683.3 | ||
| C6.3 | 529.6 | 537.2 | 533.4 | ||
| C1.6 | 776.2 | 779.3 | 777.8 | ||
| p<0.05 | 0.002** | 0.007** | - | ||
| LSD0.05% | 97.0 | 75.3 | - | ||
| CV (%) | 12.2 | 10.0 | - | ||
| F9 and F10: Filial generations, C, C1.5, C3.8, C6.3 and C1.6: Genotypes, X: Lines mean X: overall of lines, **p<0.05%, LSD: Least significant difference, CV (%): Coefficient of variation | |||||
| Table 4: Percentages of fungal infection and germination of genotypes | |||||
| Incidence (%) | Germination (%) | ||||
| Line | SS | NON | SS | NON | |
| Seed (NS) | |||||
| C | 68.13B | 78.75B | 81.88BC | 64.38AB | |
| C1.5 | 52.32B | 78.75B | 71.43CD | 60.64B | |
| C3.8 | 0.00D | 96.43A | 100.00A | 75.00AB | |
| C6.3 | 64.29B | 85.71AB | 35.79E | 14.29E | |
| C1.6 | 25.00C | 50.00C | 37.50E | 20.84D | |
| x̄ | 44.25 | 63.84 | 67.72 | 50.77 | |
| C, C1.5, C3.8, C6.3 and C1.6: Genotypes, X: lines mean, SS: Surface sterilized, Non: Non-surface sterilized-tests were rune in triplicates. Counts represent the number of fungi/100 seeds incubated at 27±2° C. Each figure represents average of three replicates, incubated at 27±2�C for 9 days (solid) PDA medium. The same letter: No differences | |||||
Counts represent the number of fungi/100 seeds incubated at 27±2°C. Each figure represents average of three replicates, incubated at 27±2°C for 9 days (solid) PDA medium. The same letter: No differences (6.9%), phenyl ester (6.77%) and pinolenic acid (4.9%). This is in addition to the presence of some compounds that have antifungal activity such as methyl ester, naphthalene acetic acid, cyanuric acid, hydroxyisocaproic acid, methylcoumarin and xanthine.
Identification and quantification of the constituent of C seeds: The 32 major bioactive chemical compounds identified from crude ethanolic extract of C seeds were Cis-13-Eicosenoic acid, n-Hexadecanoic acid, Dehydrocholic acid, Erucic acid, Indole-3-pyruvic acid, Octadecanoic acid, Oleic acid, butyl ester, Cis-Vaccenic acid, Pentadecanoic acid and Oleic acid which presented by 60.6, 6.19, 4.58, 2.2, 1.79, 1.71, 1.43, 1.31, 1.18 and 1.12%, respectively (Table 7). In addition to the presence of some compounds less than 1% identified also from C seeds such as Linoleic acid ethyl ester, 15-Tetracosenoic acid, methyl ester, Nonanoic acid 5-methyl-ethyl ester, Linoleic acid ethyl ester, E)-13-Docosenoic acid, Oxiraneoctanoic acid, 3-octyl-,cis, Cic-11-Eicosenoic acid, Heneicosanoic acid, methyl ester, Quinine,3,7,8,2-tetramethoxyflavone, 5,6,7,3,4-Pentamethoxyflavone, Palmitoleic acid, Oleanolic acid, Gardenin, Isomyristic acid, Β-Sitosterol, Gitoxigenin, Stigmasterol, Acitretin and 3-(3,4-Dimethoxyphenyl)-4methylcoumarin. Differences were observed in the chemical composition of the C3.8 seeds and the C type.
The GC-MS chromatogram analysis of the ethanolic extract of line C3.8 seeds (Fig. 1) showed three peaks which indicating the presence of three phytochemical constituents. On comparison of the mass spectra of the constituents with the their retention indices, either with those of authentic compounds or with data published in the literature the three phytocompounds were characterized and identified as shown in Table 6.
On the other hand, GC-MS chromatogram analysis of the ethanolic extract of C seeds (Fig. 2) showed one peak which indicating the presence of one major constituent (Cis-13-Eicosenoic acid) presenting 60.6% (Table 7). This may explain the why line C3.8 high resistance to the wilt disease caused by the fungus Fusarium oxysporum.
Molecular analysis: The R-ISSR (RAPD+ISSR) technique was used to detect markers linked to Fusarium wilt resistance. The ten combinations between RAPD (OP-A10 and OP-B05 primers) and ISSR (five primers) revealed a polymorphism, which six only combinations developed molecular markers linked to Fusarium wilt resistance as shown in Fig. 3 (from a to j) and summarized in Table 8.
These results demonstrated that the combination of OP-A10 primer with ISSR primers as shown in Fig. 3(a, b, e, f and i) for the five tested primers gave a total of 44 bands were produced of which 18 (40.9%) bands were polymorphic. The number of bands varied from 11 for the OP-A10+UBC-855 primer to 7 for the OP-A10+UBC-825 primer. The OP-A10 primer produced smeared profiles when combined with different ISSR primers comparing to OP-B05 primer. This combination gave polymorphism percentages ranged from 27.2% (for the OP-A10+UBC-855) to 57.1% (for the OP-A10+UBC-825) with an average of 42.4%. As for OP-B05 primer with ISSR primers shown in Fig. 3(c, d, g, h, j), which gave a total of 44 bands were produced of which 23(52.3%) bands were polymorphic. The number of bands varied from 11 for the OP-B05+UBC-864 primer to 8 for the OP-B05+UBC-811, OP-B05+UBC-825 and OP-B05+UBC-855 primers. The bands were clear and distinct.
This combination gave highly informative polymorphism percentages ranged from 12.5% (for the OP-B05+UBC-825) to 72.7% (for the OP-B05+UBC-864) with an average of 50.36%. It is interesting to note that the highest number of markers was amplified by the combination of OP-B05 primer with ISSR primers (eleven markers).
| Table 5: Frequency occurrence percentage of fungi and total fungal count (CFU/100 seeds) isolated from genotypes | ||||||||||||
| Genotypes | ||||||||||||
| Fungal genera and species | C | C1.5 | C3.8 | C6.3 | C1.6 | X | ||||||
| Alternaria alternata | 3.0 | - | - | 8.3 | - | 2.3 | ||||||
| Aspergillus flavus | 15.2 | 26.7 | - | 4.2 | 6.7 | 10.6 | ||||||
| Aspergillus niger | 18.2 | 53.3 | - | 12.5 | 6.7 | 18.1 | ||||||
| Aspergillus ochraceus | 3.0 | - | - | - | 6.7 | 1.9 | ||||||
| Aspergillus terreus | 3.0 | - | - | - | - | 0.6 | ||||||
| Aspergillus versicolor | 3.0 | - | - | 4.2 | 6.7 | 2.8 | ||||||
| Chatomium globosum | 3.0 | - | - | - | - | 0.6 | ||||||
| Fusarium culmorum | 3.0 | - | - | - | 6.7 | 1.9 | ||||||
| Fusarium oxysporum | 18.2 | 13.3 | - | 16.7 | 13.3 | 12.3 | ||||||
| Fusarium solani | 2.0 | - | - | 4.2 | 6.7 | 2.6 | ||||||
| Macrophomina phaseolina | 3.0 | 6.7 | - | 8.3 | 6.7 | 4.9 | ||||||
| Mucor spp. | 6.0 | - | 16.7 | 13.3 | 7.2 | |||||||
| Penicillium chrysogenum | 3.0 | - | - | 13.3 | 3.3 | |||||||
| Penicillium spp. | 9.2 | - | 8.2 | 6.7 | 4.8 | |||||||
| Rhizopus nigricans | 3.0 | 50.0 | 16.7 | 6.7 | 15.3 | |||||||
| Trichoderma harzianum | - | 50.0 | - | - | 10.0 | |||||||
| Total (CFU/100 seeds) | 471.4 | 214.3 | 28.6 | 342.9 | 214.3 | 1528.9 | ||||||
| Total No. of species | 15 | 4 | 2 | 10 | 12 | |||||||
| C, C1.5, C3.8, C6.3 and C1.6: Genotypes, X: lines mean | ||||||||||||
| Table 6: Compounds present in the ethanol fraction of line C3.8 seeds using GC-MS analysis | |||||||||||
| No. | RT | AS | Compound name | ||||||||
| 1 | 3.14 | 4.9 | Pinolenic acid | ||||||||
| 2 | 12.5 | 6.7 | Phosphor amidic acid,(1-methylethyl)-,ethyl 3-methyl1-4-(methyl sulfinyl) phenyl ester | ||||||||
| 3 | 12.7 | 14.9 | 4-Methoxy-α-naphthoflavone | ||||||||
| 4 | 13.0 | 1.1 | 5,8,11-Eicosatriynoic acid, methyl ester | ||||||||
| 5 | 13.3 | 0.88 | 1-Naphthalene acetic acid | ||||||||
| 6 | 13.8 | 6.99 | N-Hexadecanoic acid | ||||||||
| 7 | 14.4 | 1.5 | 7,10-Octadecadienoic acid, methyl ester | ||||||||
| 8 | 14.4 | 0.52 | Cis-Vaccenic acid | ||||||||
| 9 | 14.6 | 50.9 | 2-Heptenoic acid, pentyl ester | ||||||||
| 10 | 16.2 | 1.6 | 2-Heptenoic acid, heptyl ester | ||||||||
| 11 | 16.7 | 0.98 | Octanoin acid,6- (4 carboxy-3-methylbutoxy)-7-hydroxy-,dimethyl ester | ||||||||
| 12 | 17.2 | 0.51 | Cyanuric acid | ||||||||
| 13 | 17.4 | 1.0 | Cyclohexanecarboxylic acid, heptadecyl ester | ||||||||
| 14 | 17.6 | 0.94 | Adipic acid, 2-ethylhexyl isobutyl ester | ||||||||
| 15 | 18.0 | 0.62 | Octanoic acid, 6-(4-carboxyl-3-methylbutoxy)-7-hydroxy-domethyl ester | ||||||||
| 16 | 19.1 | 0.96 | 2-hydroxyisocaproic acid, acetate | ||||||||
| 17 | 20.6 | 0.80 | Eicosanoic acid | ||||||||
| 18 | 20.9 | 0.59 | 1,2-Benzenediol, 3,5-bis(1,1dimethylethyl) | ||||||||
| 19 | 21.7 | 0.74 | Oleic acid,3-9octadecyloxy) propyl ester | ||||||||
| 20 | 21.8 | 0.89 | Glycidyl oleate | ||||||||
| 21 | 22.4 | 0.59 | 3-(3,4-Dimethoxyphenyl)-4-methylcoumarin | ||||||||
| 22 | 22.7 | 0.51 | Xanthine | ||||||||
| 23 | 23.1 | 0.53 | 2-Hydroxychalcone | ||||||||
| RT: Retention time, AS: Area sum (%) | |||||||||||
However, the less number of markers was observed in the case of OP-A10 primer combined with ISSR primers (two markers). The combination of OP-A10 primer with ISSR primers exhibited two positive markers linked to resistance with molecular sizes of 1236 bp for the OP-A10+UBC-825 and 1276 bp for the OP-A10+UBC-855 primers. On the other hand, the combination of OP-B05 primer with ISSR primers exhibited eight positive markers linked to resistance with molecular sizes of 261 bp for the OP-B05+UBC-811, 1209 bp for the OP-B05+UBC-825, 1158 bp for the OP-B05+UBC-855, 2154, 1515, 636, 438 and 350 bp for the OP-B05+UBC-864 primers. And three negative markers with molecular sizes of 520 bp for the OP-B05+UBC-811, 1293 and 331 bp for the OP-B05+UBC-855 primers. The genetic similarity coefficient varied from 0.74 between C and C3.8 also between C and C1.6, while was 0.95 between C1.6 and C6.3 (Table 9).
| Table 7: Compounds present in the ethanol fraction of C seeds using GC-MS analysis | |||||
| No. | RT | AS (%) | Compound name | ||
| 1 | 13.7 | 6.19 | n-Hexadecanoic acid | ||
| 2 | 13.8 | 1.79 | Octadecanoic acid | ||
| 3 | 14.4 | 0.9 | Linoleic acid ethyl ester | ||
| 4 | 14.4 | 1.12 | Oleic acid | ||
| 5 | 14.6 | 0.65 | 15-Tetracosenoic acid, methyl. ester | ||
| 6 | 14.8 | 60.6 | Cis-13-Eicosenoic acid | ||
| 7 | 14.8 | 4.58 | Erucic acid | ||
| 8 | 14.9 | 1.31 | Cis-Vaccenic acid | ||
| 9 | 15.0 | 0.84 | Nonanoic acid, 5-methyl-ethyl.ester | ||
| 10 | 15.4 | 0.95 | 9,15-Octadecadienoic acid.methyl ester | ||
| 11 | 15.9 | 0.79 | Linoleic acid ethyl ester | ||
| 12 | 16.0 | 0.74 | E)-13-Docosenoic acid | ||
| 13 | 16.2 | 0.82 | Oxiraneoctanoic acid, 3-octyl-,cis | ||
| 14 | 16.3 | 0.87 | Cic-11-Eicosenoic acid | ||
| 15 | 16.5 | 0.78 | Heneicosanoic acid, methyl ester | ||
| 16 | 16.8 | 1.43 | Oleic acid, butyl ester | ||
| 17 | 17.0 | 0.75 | Quinine | ||
| 18 | 17.4 | 0.93 | 3,7,8,2-tetramethoxyflavone | ||
| 19 | 17.8 | 0.8 | 5,6,7,3,4-Pentamethoxyflavone | ||
| 20 | 18.0 | 0.79 | Palmitoleic acid | ||
| 21 | 18.7 | 0.67 | Oleanolic acid | ||
| 22 | 18.8 | 0.69 | Gardenin | ||
| 23 | 19.1 | 0.69 | Isomyristic acid | ||
| 24 | 19.5 | 1.71 | Dehydrocholic acid | ||
| 25 | 19.7 | 0.66 | Gitoxigenin | ||
| 26 | 19.8 | 1.06 | ∞-Santonin | ||
| 27 | 20.1 | 0.98 | ?-Sitsterol | ||
| 28 | 20.3 | 0.78 | Stigmasterol | ||
| 29 | 20.6 | 0.69 | Acitretin | ||
| 30 | 21.5 | 0.97 | 3-(3,4-Dimethoxyphenyl)-4methylcoumarin | ||
| 31 | 21.8 | 1.18 | Pentadecanoic acid, 14-bromo | ||
| 32 | 22.9 | 2.2 | Indole-3pyrovic acid | ||
| RT: Retention time, AS: Area sum (%) | |||||
| Table 8: R-ISSR primers, total number of bands, number of polymorphic bands, polymorphism percentage, molecular size, marker type and molecular weight of the product | |||||||
| Primer | Primer name | TB | PB | P (%) | MS | MT | MW (bp) |
| R-ISSR | OP-A10+UBC-811 | 9 | 4 | 44.4 | - | - | 368-1895 |
| (OP-A10+ISSRs) | OP-A10+UBC-825 | 7 | 4 | 57.1 | 1236 | P | 520-1643 |
| OP-A10+UBC-855 | 11 | 3 | 27.2 | 1276 | P | 232-1784 | |
| OP-A10+UBC-864 | 9 | 3 | 33.3 | - | - | 293-1593 | |
| OP-A10+UBC-868 | 8 | 4 | 50.0 | - | - | 371-1301 | |
| Total | 44 | 18 | 2 | ||||
| R-ISSR | OP-B05+UBC-811 | 8 | 4 | 50.0 | 520 | N | 329-1456 |
| (OP-B05+ISSRs) | 261 | P | |||||
261 | P | ||||||
| OP-B05+UBC-825 | 8 | 1 | 12.5 | 1209 | P | 330-1209 | |
| OP-B05+UBC-855 | 8 | 4 | 50.0 | 1293 | N | 331-1293 | |
1158 | P | ||||||
331 | N | ||||||
| OP-B05+UBC-864 | 11 | 8 | 72.7 | 2154 | P | 350-2154 | |
1515 | P | ||||||
636 | P | ||||||
438 | P | ||||||
350 | P | ||||||
| OP-B05+UBC-868 | 9 | 6 | 66.6 | - | - | 336-1914 | |
| Total | 44 | 23 | 11 | ||||
| ISSR: Inter simple sequence repeat (ISSR) polymorphism, TB: Total number of bands, PB: Number of polymorphic bands, P (%): Polymorphism percentage, MS: Molecular size, MT: Marker type, MW: Molecular weight of the product (bp) P: Positive marker, N: Negative marker | |||||||
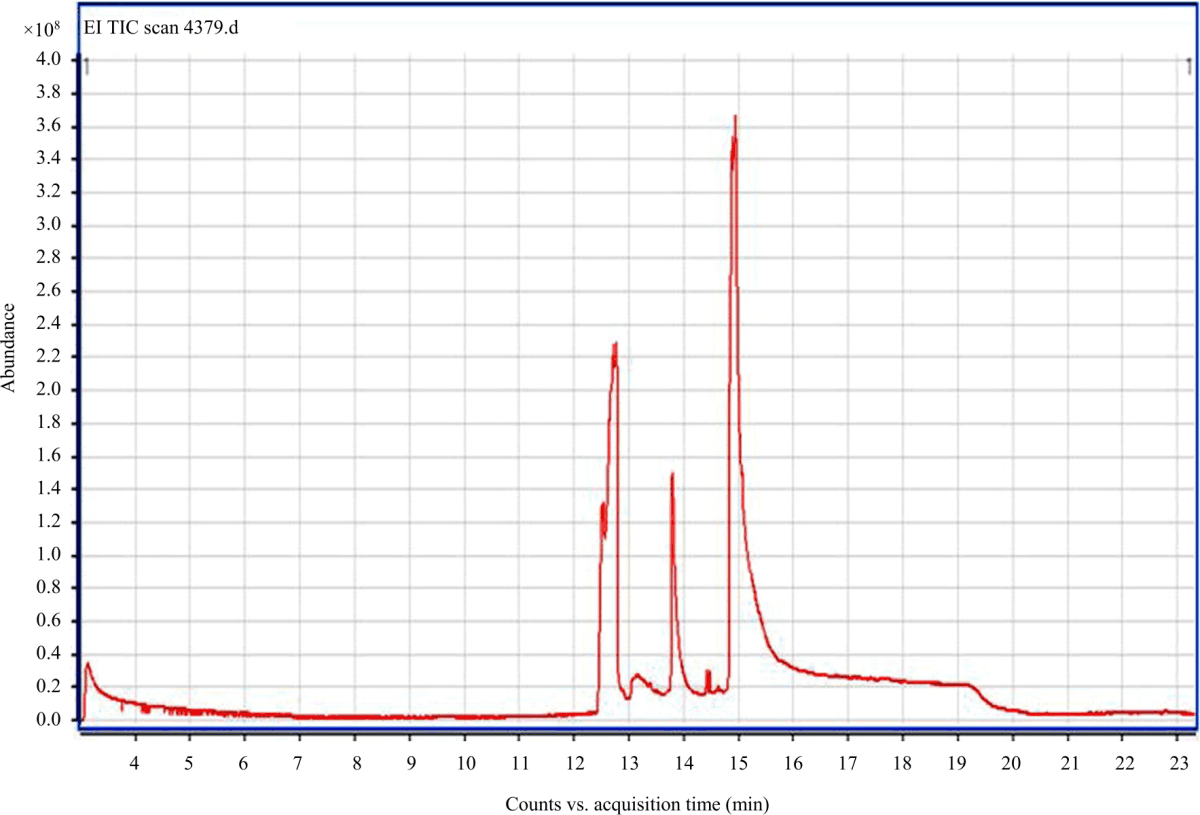 |
| Fig. 1: | GC-MS chromatogram of ethanolic extract of line C3.8 seeds |
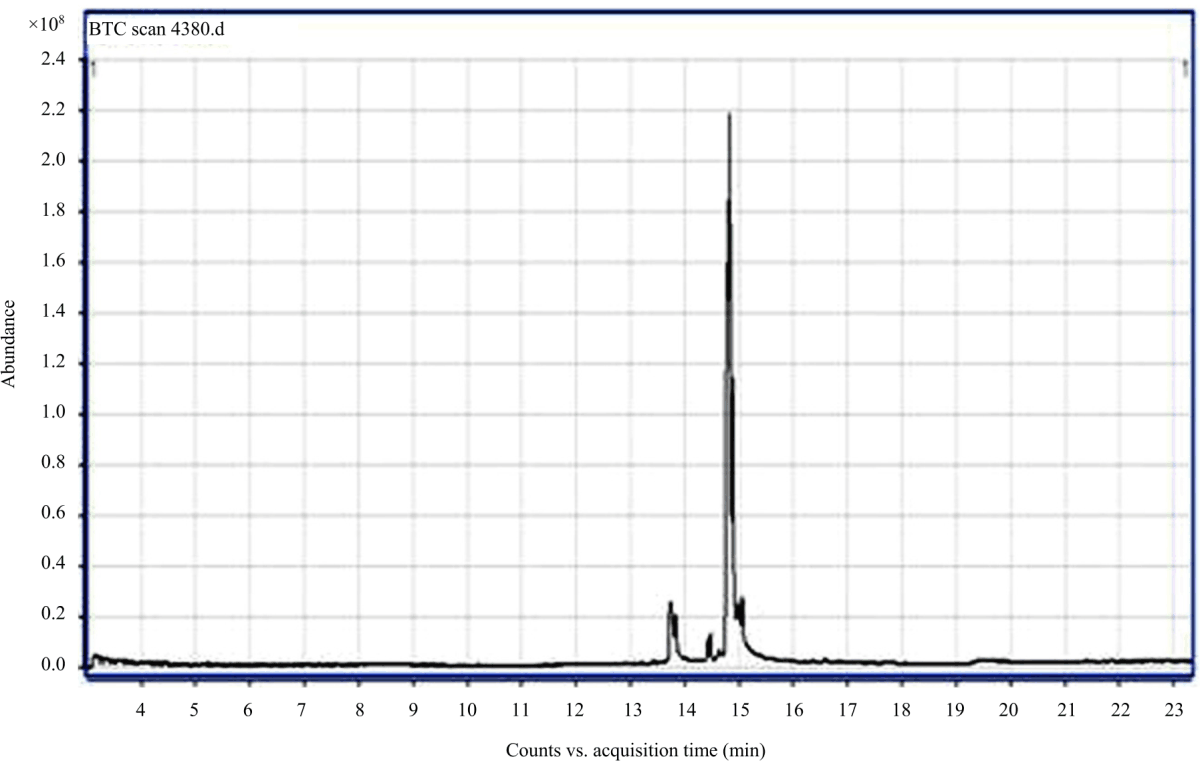 |
| Fig. 2: | GC-MS chromatogram of ethanolic extract of line C seeds |
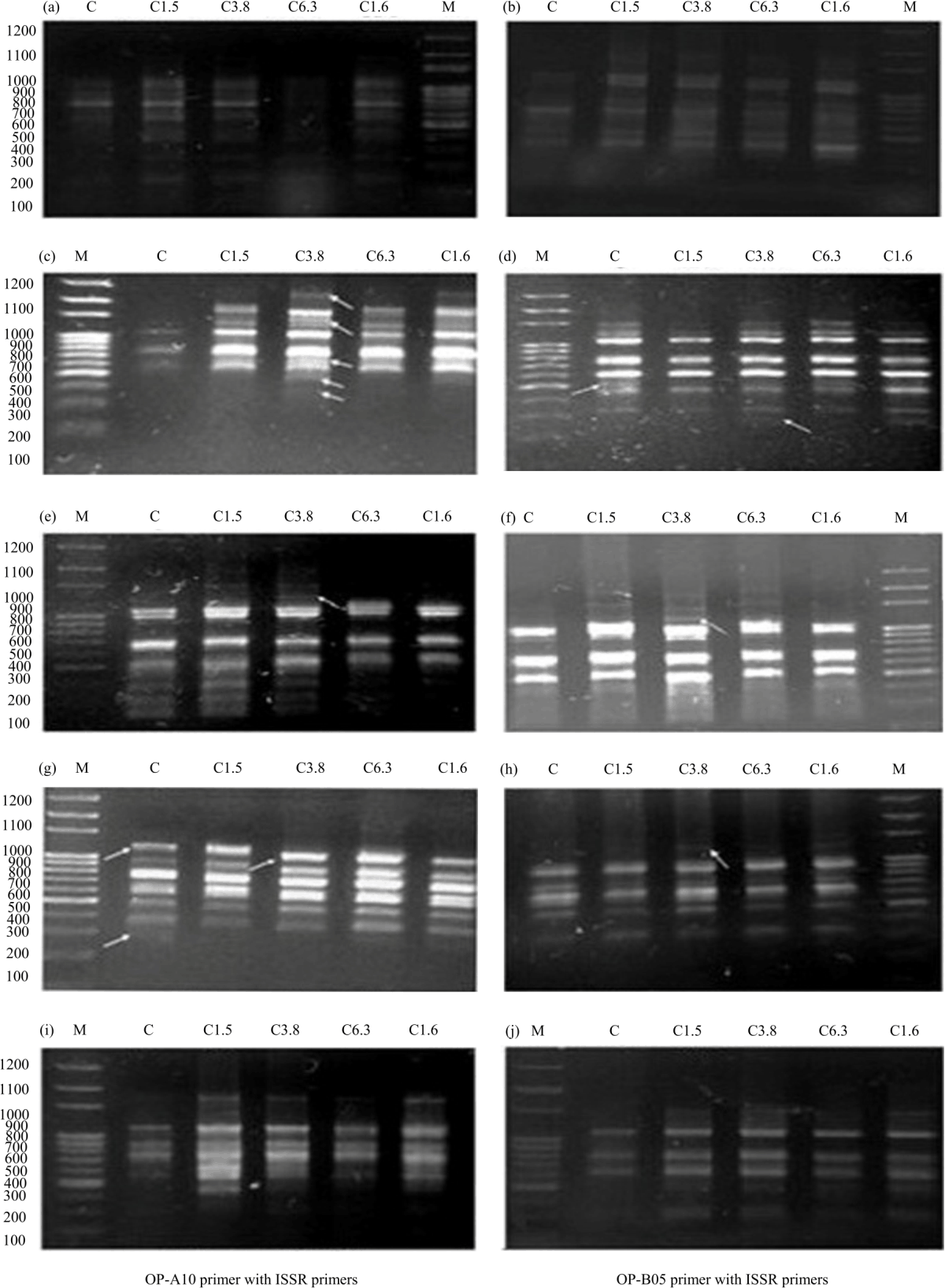 |
| Fig. 3(a-j): | Banding patterns of R-ISSR fragments of the five samples (a) A10+UBC-864, (b) A10+UBC-811, (c) B05+UBC-864, (d) B05+UBC-811, (e) A10+UBC-855, (f) A10+UBC-825, (g) B05+UBC-855, (h) B05+UBC-825, (i) A10+UBC-868 and (j) B05+UBC-868. M: Marker, C, C1.5, C3.8, C6.3 and C1.6 |
| Table 9: Genetic similarity matrix of the five samples based on R-ISSR markers | |||||
| Genotypes | C | C1.5 | C3.8 | C6.3 | C1.6 |
| C | 1 | ||||
| C1.5 | 0.79 | 1 | |||
| C3.8 | 0.74 | 0.89 | 1 | ||
| C6.3 | 0.79 | 0.90 | 0.92 | 1 | |
| C1.6 | 0.74 | 0.92 | 0.93 | 0.95 | 1 |
| C, C1.5, C3.8, C6.3 and C1.6: Genotypes | |||||
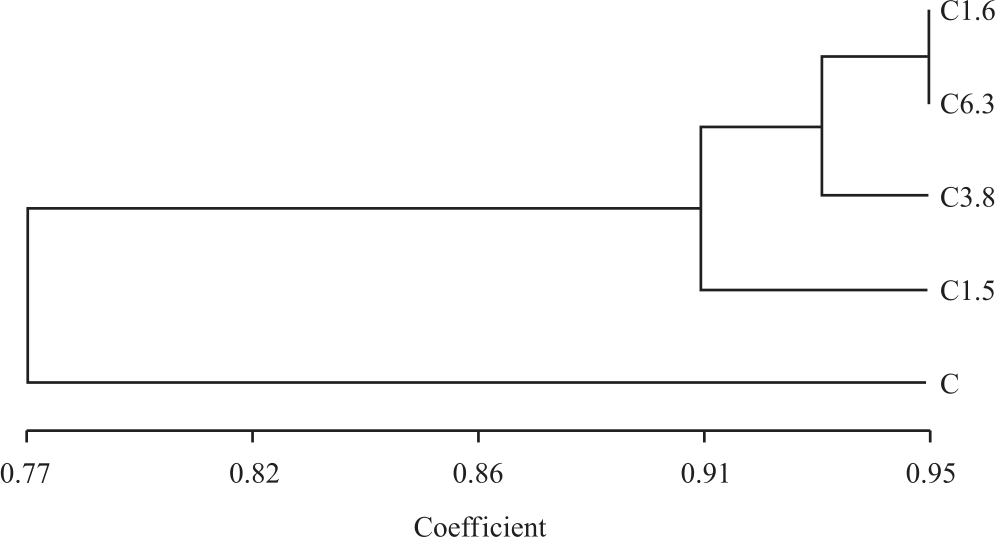 |
| Fig. 4: | UPGMA dendrograms of the five samples based on the genetic similarity matrix obtained with the Jaccard’s index for the data from R-ISSR marker |
The constructed dendrogram obtained by UPGMA analysis divided these samples into two main clusters (Fig. 4). The first one included only C. The other cluster was divided into two main sub-clusters; the first one included only C1.5, while the other one was divided into two sub-sub clusters, the first one included only C3.8, while the second one included C1.6 and C6.3. According to R-ISSR analysis, these results indicated that the most closely related samples C1.6 and C6.3, which were located in the same sub-sub cluster, while the most dissimilar samples were C, C3.8 and C1.6.
DISCUSSION
The sesame crop faces constant challenges due to fungi diseases, which limits the cultivation of sesame in Egypt. Therefore, the development of new varieties of sesame that combine high productivity with resistance to fungal diseases becomes an urgent need. The current study was based on four axes as follows; evaluation of promising lines based on the seed yield, classified of genotypes based on some fungal diseases resistance, detection of some antibiotic polyphenolic compositions in genotypes and determine molecular markers linked resistance to Fusarium wilt. According to the analysis of variance of the data, genotypes differed statistically significant (p<0.05) for seed yield ha–1 in F8 and F9 generations, indicating that there is sufficient genetic variation for phenotypic selection (Table 3). The highest average seed yield ha–1 was obtained from C followed by C1.5. Commercial variety (C) is well adapted to the growing environments also, line C1.5 may be due to its ability to genetically regulate by switching genes for expression30-32.
The sterilized seeds of line C3.8 when infected with fungal the percentage was zero and this line germinated by 100%25. May be due to impenetrable reaction, due to the hypersensitive the reactions of line, resulting from the prevention of pathogen to reach the line (host) consequently, the rate of reproductions of it become zero. The line C3.8 (host) has resistance genes; the disease (pathogen) has genes for virulence and resistance reaction would be the result when the genes of a pathogen are not able to identify with all the resistance genes in line with proper virulence genes according to the gene-for-gene relationship33.The frequency occurrence percentage of fungi and total fungal count (CFU/100 seeds) isolated from genotypes seeds on PDA medium in Table 5 clearly indicated that line C3.8 showed low densities compared to other genotypes. May be due to the presence of certain substances in it which have antifungal activity on fungal growth, Shabana et al.6 found that line C3.8 was highly resistant against wilt disease caused by Fusarium oxysporum. In this study, GC/MS analysis used to interpret why line C3.8 showed low fungal densities compared to C. Data in Tables 6 and 7 showed differences in the chemical compositions of the C3.8 and C (Fig. 1 and 2). The line C3.8 seed has some chemical compositions that have antifungal activity but absents in C. Perhaps line C3.8 was a hypersensitive the response, which makes certain unique molecules, called elicitors, of a pathogen. The elicitors are recognized by receptor-like molecules present in the line. This recognition sets several biochemical reactions to the production phytoalexin, which are antibiotic polyphenolic compositions15. Bawazir et al.34 found that sesame oil compounds have a negative effect and antifungal activity against Aspergillus niger and Aspergillus flavus. It was clear from this study; line C3.8 has defense response genes against fungi diseases, which produced bioactive chemical compounds that conferred resistance. This study suggested that some chemical compounds found in line C3.8 can use as a basis for generate new broad-spectrum antimicrobial formulations. The results of this study demonstrate the possibility of using the R-ISSR technique could be used to select the resistant and susceptible lines to Fusarium wilt. It was shown that R-ISSR can be effectively generated more markers linked to Fusarium wilt resistance (10 positives+ 3 negative markers) compared with RAPD and ISSR when used alone [five RAPD and three ISSR markers (in press)]. This result indicated that the combination of ISSR and RAPD primers amplified regions that will fail to amplify when they are used alone. Some authors noted that when study genotypes close related, the analysis of variability could require more than one DNA-based technique35. The 10 R-ISSR combinations revealed polymorphic banding patterns amongst the five tested samples with 41 polymorphic bands out of 88 bands that covered the molecular size between 232 and 2154 bp. The average of polymorphism obtained in this study (46.38%) was high compared with previous studies in which a low level of polymorphism (33%) in sesame with ISSR markers36,37, but this average was low as compared with (70.1%) in sesame with RAPD markers. Finally, the results confirm that C was different from the four tested lines, especially; C3.8 which observed more bands didn’t appear in C. This part of the study is complementary to the previous study6. In F6 generation, they (excluding line C1.5) were evaluated under artificially infested soil with M. phaseolina and F. oxysporum as the causal pathogens of charcoal rot and wilt under greenhouse conditions and open field. The results were as follows: Lines C6.3 and C3.8 were the most resistant ones for charcoal rot disease. Lines C1.6, C3.8 and C6.3 were the most resistant lines against F. oxysporum infection. From this results, line C3.8 can be considered as novel genes in the primary gene pool to resistant to Fusarium wilt disease in sesame.
CONCLUSION
Soil borne diseases are among the most destructive elements in sesame production and resistant varieties are the appropriate solution to confront this problem. Therefore, new sesame lines and commercial variety Shandaweel were evaluated based on seed yield ha–1, tested for seed-borne fungi and examined using R-ISSR. The line C3.8 recorded the low fungal incidence and it has chemical compositions that have antifungal activity and absent in C. R-ISSR technique can be used for identification of molecular markers linked to Fusarium wilt resistance. Line C3.8 is offering aspects to form new varieties resistant to Fusarium wilt disease in sesame. And, this line has compounds (antifungal activity) can use as a basis for generate new broad-spectrum antimicrobial formulations.
SIGNIFICANCE STATEMENT
This study discovered that line C3.8 has chemical compositions that have antifungal activity that can be beneficial for generating new broad-spectrum antimicrobial formulations. R-ISSR generated more markers linked to Fusarium wilt resistance. This study will help the sesame breeder to use new valuable resource to form new varieties resistant to Fusarium wilt in sesame.
ACKNOWLEDGMENTS
Many thanks to the National Research Centre, Egypt, for funding this study. Special thanks to Prof. Adel Hoballah, Cairo University for the development of genetic materials.
REFERENCES
- Johnson, L.A., T.M. Suleiman and E.W. Lusas, 1979. Sesame protein: A review and prospectus. J. Am. Oil Chem. Soc., 56: 463-468.
CrossRefDirect Link - Warra, A.A., 2011. Sesame (Sesamum indicum L.) Seed oil methods of extraction and its prospects in cosmetic industry: A review. Bayero J. Pure Applied Sci., 4: 164-168.
Direct Link - Lee, J., Y. Lee and E. Choe, 2008. Effects of sesamol, sesamin, and sesamolin extracted from roasted sesame oil on the thermal oxidation of methyl linoleate. LWT-Food Sci. Technol., 41: 1871-1875.
CrossRefDirect Link - Islam, F., R.A. Gill, B. Ali, M.A. Farooq, L. Xu, U. Najeeb and W. Zhou, 2016. Sesame. In: Breeding Oilseed Crops for Sustainable Production: Opportunities and Constraints, Gupta, K.S. (Ed.)., Elsevier Science, Cambridge, USA, pp: 135-147.
Direct Link - Wang, L., Y. Zhang, X. Zhu, X. Zhu and D. Li et al., 2017. Development of an SSR-based genetic map in sesame and identification of quantitative trait loci associated with charcoal rot resistance. Sci. Rep., 7: 1-8.
CrossRefDirect Link - Shabana, R., A.A.A. El-Mohsen, M.M.A. Khalifa and A.A. Saber, 2014. Quantification of resistance of F6 sesame elite lines against Charcoal-rot and Fusarium wilt diseases. Adv. Agric. Biol., 1: 144-150.
CrossRefDirect Link - Blal, A., S. Kamel, H. Mahfouz and M. El-Wahed, 2012. Impact of pollination and fertilization on sesame production in the reclaimed lands, Ismailia governorate, Egypt. J. Agric. Sci., 57: 121-133.
CrossRefDirect Link - Li, D.H., L.H. Wang, Y.X. Zhang, H.X. Lv, X.Q. Qi, W.L. Wei and X.R. Zhang, 2012. Pathogenic variation and molecular characterization of Fusarium species isolated from wilted sesame in China. Afr. J. Microbiol. Res., 6: 149-154.
Direct Link - Savary, S., A. Ficke, J.N. Aubertot and C. Hollier, 2012. Crop losses due to diseases and their implications for global food production losses and food security. Food Secur., 4: 519-537.
CrossRefDirect Link - Velásquez, A.C., C.D.M. Castroverde and S.Y. He, 2018. Plant and pathogen warfare under changing climate conditions. Curr. Biol., 28: R619-R634.
CrossRefDirect Link - Rao, N.R. and M. Vijayalakshmi, 2000. Studies on Alternaria sesami pathogenic to sesame. Micro. Res., 155: 129-131.
CrossRefDirect Link - Nayyar, B.G., S. Woodward, L.A. Mur, A. Akram, M.S. Arshad, M. Naqvi and S. Akhund, 2017. The incidence of Alternaria species associated with infected Sesamum indicum L. seeds from fields of the Punjab, Pakistan. Plant Path. J., 33: 543-553.
CrossRefDirect Link - Kato, M.J., A. Chu, L.B. Davin and N.G. Lewis, 1998. Biosynthesis of antioxidant lignans in Sesamum indicum seeds. Phytochemistry, 47: 583-591.
CrossRefDirect Link - Qadir, A., A. Ali, M. Arif, A.H. Al-Rohaimi and S.P. Singh et al., 2017. Solvent extraction and GC-MS analysis of sesame seeds for determination of bioactive antioxidant fatty acid/fatty oil components. Drug Res., 68: 344-348.
CrossRefDirect Link - Myint, D., S.A. Gilani, M. Kawase and K.N. Watanabe, 2020. Sustainable sesame (Sesamum indicum L.) production through improved technology: An overview of production, challenges, and opportunities in Myanmar. Sustainability, 12: 1-21.
CrossRefDirect Link - Wu, M.S., L.B.B. Aquino, M.Y.U. Barbaza, C.L. Hsieh, K.A. De Castro-Cruz, L.L. Yang and P.W. Tsai, 2019. Anti-inflammatory and anticancer properties of bioactive compounds from sesame (Sesamum indicum L.). Molecules, 24: 1-24.
CrossRefDirect Link - Esse, H.P., T.L. Reuber and D. Does, 2020. Genetic modification to improve disease resistance in crops. New Phytol., 225: 70-86.
CrossRefDirect Link - Panth, M., S.C. Hassler and F. Baysal-Gurel, 2020. Methods for management of soil borne diseases in crop production. Agriculture, 10: 1-21.
CrossRefDirect Link - Yumurtaci, A., 2015. Utilization of Diverse Sequencing Panels for Future Plant Breeding. In: Advances in Plant Breeding Strategies: Breeding, Biotechnology and Molecular Tools, Al-Khayri, J.M., S.M. Jain and D.V. Johnson (Eds.)., Springer International Publishing, Cham, Switzerland, pp: 539–561.
CrossRefDirect Link - Maisuria, H.J., R.M. Patel and K.P. Suthar, 2017. Validation of molecular markers linked to Fusarium wilt resistance in Chickpea genotypes. Int. J. Pure App. Biosci., 5: 254-260.
CrossRefDirect Link - Mahmoud, F.A and E.S. Abd El-Fatah, 2020. Genetic diversity studies and identification of molecular and biochemical markers associated with Fusarium wilt resistance in cultivated faba bean (Vicia faba L.). Plant Pathol. J., 36: 11-28.
CrossRefDirect Link - Saleh, B., 2011. R-ISSR marker as a useful tool for detection of new genomic loci in Arthrocnemum macrostachyum. Biol. Plant., 55: 327-330.
CrossRefDirect Link - Altaf, N., K. Shahid Akhtar, A. Mushtaq, A. Rehana, R. Ashfaq Ahmed, S. Shabnum, M. Zafar and M. Saqib, 2004. Seed borne mycoflora of sesame (Sesamum indicum L.) and their effect on germination and seedling. Pak. J. Biol. Sci., 7: 243-245.
CrossRefDirect Link - Samson, R.A., J. Houbraken, U. Thrane, J.C. Frisvad and B. Andersen, 2010. Food and Indoor Fungi. CBS-KNAW Fungal Biodiversity Centre, Utrecht, Netherlands, ISBN: 9789070351823, Pages: 390.
Direct Link - Davies, N.W., 1990. Gas chromatographic retention indices of monoterpenes and sesquiterpenes on methyl silicon and Carbowax 20M phases. J. Chromatogr. A, 503: 1-24.
CrossRefDirect Link - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link - Rohlf, F.J., 1987. NTSYS-pc: Microcomputer programs for numerical taxonomy and multivariate analysis. Am. Stat., 41: 330-330.
CrossRefDirect Link - Baraki, F., Y. Tsehaye and F. Abay, 2016. Analysis of genotype × environment interaction and seed yield stability of sesame in Northern Ethiopia. J. Plant Breed. Crop Sci., 8: 240-249.
Direct Link - Anter, A.S., 2019. Identification of drought-tolerant sesame (Sesamum indicum L.) lines and effect of drought stress on xanthium strumarium and cynodon dactylon weeds. Middle East J. Appl. Sci., 9: 1038-1046.
CrossRefDirect Link - Baraki, F. and M. Berhe, 2019. Evaluating performance of sesame (Sesamum indicum L.) genotypes in different growing seasons in Northern Ethiopia. Inter. J. Agron., 2019: 1-7.
CrossRefDirect Link - Dodds, P.N. and J.P. Rathjen, 2010. Plant immunity: Towards an integrated view of plant-pathogen interactions. Nat. Rev. Gen., 11: 539-548.
CrossRefDirect Link - Bawazir, A.M.A. and M. Shantaram, 2018. Effect of Yemeni sesame oil against some pathogenic bacteria and fungi. Inter. J. Pharma. Sci. Res., 9: 2507-2512.
Direct Link - Costa, R., G. Pereira, I. Garrido, M.M. Tavares-de-Sousa and F. Espinosa, 2016. Comparison of RAPD, ISSR and AFLP molecular markers to reveal and classify orchardgrass (Dactylis glomerata L.) germplasm variations. PLoS ONE, Vol. 11.
CrossRefDirect Link - Kumar, V. and S.N. Sharma, 2011. Comparative potential of phenotypic, ISSR and SSR markers for characterization of sesame (Sesamum indicum L.) varieties from India. J. Crop Sci. Biotechnol., 14: 163-171.
CrossRefDirect Link - Parsaeian, M., A. Mirlohi and G. Saeidi, 2011. Study of genetic variation in sesame (Sesamum indicum L.) using agro-morphological traits and ISSR markers. Russ. J. Gene., 47: 314-321.
CrossRefDirect Link