
Brexidis Mandila
School of Natural Resource Management, University of Eldoret, P.O. Box 1125-30100, Eldoret, Kenya
LiveDNA: 254.33730
Kenneth Odhiambo
School of Natural Resource Management, University of Eldoret, P.O. Box 1125-30100, Eldoret, Kenya
Alice Muchugi
World Agroforestry (ICRAF), P.O. Box 30677-00100, Nairobi, Kenya
Daniel Nyamai
School of Agriculture, Natural Resources and Environmental Studies, Rongo University, P.O. Box 103-40404, Rongo, Kenya
Agnes Gachuiri
World Agroforestry (ICRAF), P.O. Box 30677-00100, Nairobi, Kenya
Asian Journal of Plant Sciences
Year: 2021 | Volume: 20 | Issue: 2 | Page No.: 183-195







ABSTRACT
Background and Objective: Cultivating Calotropis procera for fiber supply to the textile industry can improve the livelihoods of communities in arid and semi-arid regions. This study determined edaphic and climatic factors affecting phenological traits of C. procera in the semi-arid regions of Kenya. Materials and Methods: Repeated measure research design was used with multistage sampling technique to monitor activity indices, number of flowers and fruits and phenophase intensities. Climatic and edaphic factors of study sites were also monitored. Data was analyzed using linear, Poisson log linear regression based on Generalized Estimation Equation (GEE) and Mixed Analysis of Variance (ANOVA). Results: High Soil Organic Carbon (OC) content (3%) and exchangeable Na (112.5 ppm) at (0-20) cm soil depth were recorded in Tharaka. High mean monthly rainfall (160.37 mm) was recorded in Makueni. Flowering activity indices in (June-August, 2018) were 64.97% and 69.6% in Tharaka and Makueni, respectively . Available P, average monthly rainfall and temperature had significant association with flowering and fruiting activity indices (p<0.05). The mean number of flowers and fruits per stem were significantly associated with soil available P, exchangeable Na and OC content (p<0.05). Though edaphic factors were not significantly associated with phenophase intensities of C. procera, average monthly rainfall and temperature were positively and negatively associated with phenophase intensities, respectively . Conclusion: Available P, exchangeable Na, available K and OC content noticeably affect phenological traits of naturally growing C. procera. Rains and temperatures are critical climatic factors affecting phenological traits of C. procera.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Brexidis Mandila, Kenneth Odhiambo, Alice Muchugi, Daniel Nyamai and Agnes Gachuiri, 2021. Edaphic and Climatic Factors Affecting Phenology of Naturally Growing Calotropis procera in Semi-arid Regions of Kenya. Asian Journal of Plant Sciences, 20: 183-195.
DOI: 10.3923/ajps.2021.183.195
URL: https://scialert.net/abstract/?doi=ajps.2021.183.195
DOI: 10.3923/ajps.2021.183.195
URL: https://scialert.net/abstract/?doi=ajps.2021.183.195
INTRODUCTION
Climate change is leading to acute food shortage, inadequate livestock forage and decreased income in arid and semi-arid lands (ASALs)1. Therefore, there is need to develop environment friendly and conservation conscious techniques to increase communities’ resilience to climate change. Domesticating multipurpose trees and shrubs can significantly bring new opportunities for livelihood improvements2. This is because domestication enhances provision of products and services from trees to increase productivity, combat malnutrition and adapt to anthropogenic climate change3.
Calotropis procera is among shrub species that can be domesticated in semi-arid regions. The shrub is ever green with deep and solid tap root, it is drought and salt tolerant, can grow in ecosystems with less than 1000 mm annual precipitation and temperature range of 20-30°C4,5. The species can be used for medicinal and fodder purposes, while its genes can be used in genetic modification to enhance cotton fiber strengths6. However, the shrub has been reported to be having undesirable characteristics such as invasiveness in some parts of the world like Australia7,8.
Different kind of research has established that its seeds and fruits can produce quality calotrope fiber that can be used in the textile industry. Compared to silk and cotton fiber, calotrope fiber has good stable lengths, fiber strengths, fiber uniformity ratio, fiber fairness and moisture absorption characteristics9-11. Therefore, under proper management, C. procera can be ecologically, economically, culturally and socially important to ASAL communities. However, the phenological behavior of C. procera under different climatic and edaphic conditions has received limited research attention with most studies having been conducted in greenhouses12,13. Lack of adequate information regarding this species makes it difficult to conclusively predict how climate change and changes in soil condition as a result of erosion and salination will influence the phenology of the species when domesticated14. Therefore, this study determined edaphic and climatic factors affecting the phenological traits of naturally growing C. procera in the semi-arid regions of Kenya.
MATERIALS AND METHODS
Research site: The study was carried out in the semi-arid regions of Tharaka and Makueni in the Eastern part of Kenya from June, 2018 to April, 2020. Tharaka lies between latitudes 00° 07' and 00° 26' S and longitudes 37° 19' and 37° 46' E, while Makueni lies between latitude 1° 35' and 3° 00' S and Longitudes 37° 10' and 38° 30' E. The two regions lie in the agro-climatic and eco-climatic zone V, which is characterized by low and unreliable rainfall, dispersed population, marginal agricultural lands and infertile soils15. The study was specifically conducted in lowland areas (altitude less than 600 m above sea level) that receive unreliable and poorly distributed rains of less than 500 mm per year and higher temperatures of up-to 40°C at certain periods16.
Research design: The study used a repeated measure research design by taking multiple measurements of the dependent variable on the same object over a period of time17. This was appropriate because the purpose of the study was to evaluate phenological plasticity of C. procera over a period of time under different climatic seasons and edaphic conditions.
Sampling procedures and sample sizes: Purposeful sampling technique was used in selecting research blocks (farms) with naturally growing C. procera and whose owners voluntarily allowed research to be conducted. In Makueni two blocks (Kyumani and Kyanguli) were selected while in Tharaka three blocks (Kathwana, Kilimangare and Kajiampau) were chosen. In each block, permanent main plots measuring (20×20 m) were marked using blue painted pipes. Each main plot was sub-divided into 15 permanent sub-plots measuring (5×5 m) that were demarcated using red painted pipes. Systematic random sampling technique was used in selecting sub-plots to be included in the study, where every third sub-plot was selected. The total number of sub-plots selected per plot was determined as explained by Dell et al.18, Eq. 1:
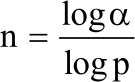 | (1) |
Where:
| N | = | Sample size (number of subplots) |
| α | = | Permitted error at 95% confidence level = 0.05 |
| p | = | Proportion of sub-plots estimated as having a particular characteristics, in this case C. procera |
Since it was not known, it was estimated at 50% (0.5) as recommended by Dell et al.18:
Therefore, number of sub-plots per plot was:
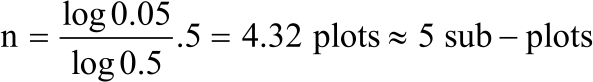 |
All C. procera stems in sub-plots were numbered and included in the sample.
In each sub-plot, one pit was randomly dug to collect subsoil at 0-20 cm and deep soil at 20-40 cm. Soil samples at 0-20 cm and 20-40 cm from all sub-plots in a plot were mixed to form subsoil and deep soil composites, respectively . From each composite in the respective depth, one sample was picked and put into 2000 g well labeled bags for laboratory analysis.
Data collection
Phenology of naturally growing C. procera: Naturally growing C. procera stems with and without flowers and fruits were identified and counted. Activity index was calculated by dividing the number of stems with flowers or fruits by the total number of stems in a sub-plot. The number of flowers and fruits (green or ripe) per stem were counted and recorded. On every stem the total number of branches, number of branches with flowers and fruits were counted in the selected sub-plots. This was used to calculate the Phenophase Intensity (Pi) levels as indicated in Eq. 2 and 313:
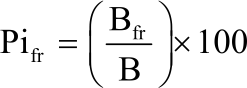 | (2) |
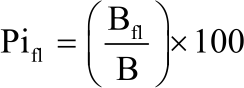 | (3) |
Where:
| Pifr and Pifl | = | Phenophase intensity levels for fruits and flowers, respectively |
| Bfr and Bfl | = | Branches with fruits and flowers, respectively |
| B | = | With total number of branches on an individual stem |
Soil properties: Soil samples were taken to Kenya Forest Research Institute (KEFRI) laboratory for analysis. Sample preparation and analysis of soil pH using pH meter, Electric Conductivity (EC) using conductivity meter, Organic Carbon (OC) using Walkley Black method, Phosphorus (P) using UV-Spectrophotometer and Magnesium (Mg), Nitrogen (N), Sodium (Na), Calcium (Ca) and Potassium (K) based on Atomic Absorption Spectrophotometer (AAS) were conducted according to Udelhoven et al.19.
Climatic factors: The geographical coordinates of the study area were used in obtaining rainfall and temperature data from National Aeronautics and Space Administration website.
Data analysis: Mixed ANOVA was used to determine statistically significant differences in the mean flowering and fruiting activity indices and phenophase intensities within research time points. Relationships between phenological traits with edaphic and climatic factors were established using linear and Poisson regression based on GEE. Analysis was conducted up-to a level that all remaining variables were significantly associated with phenological traits. Therefore, variables indicating insignificant association were removed from the model list-wise for the next analysis level.
RESULTS
Edaphic and climatic factors in Tharaka and Makueni semi-arid regions
Edaphic factors in Tharaka and Makueni semi-arid regions: Soil OC content and exchangeable Na at 0-20 cm soil horizon were 3.0% and 112.5 ppm in Tharaka and 3.08% and 75 ppm in Makueni, respectively , compared to 2.92% and 85 ppm in Tharaka and 2.63% and 74 ppm in Makueni, respectively at (20-40) cm soil depth (Table 1).
Climatic factors in Tharaka and Makueni semi-arid regions: The mean monthly rainfall of 143.83 mm and 160.37 mm were experienced in the period of (October, 2019 to February, 2020) in the semi-arid regions of Tharaka and Makueni, respectively (Fig. 1). Monthly average relative humidity of 60.42% and 61.52% and wind speed of 3.6 m/s and 3.07 m/s were experienced in (April-September, 2019) in Tharaka and Makueni, respectively (Fig. 2).
Factors Affecting Flowering and Fruiting Activity Indices of C. procera
Flowering and fruiting activity indices of C. Procera : Flowering activity indices of naturally growing C. procera decreased from 75.87% in Tharaka and 64.97% in Makueni in (June-August, 2018) to 48.05% in Tharaka and 50.48% in Makueni in (September-November, 2019) (Fig. 3). Similarly, fruiting activity indices decreased from 83.06% in Tharaka and 69.6% in Makueni to 42.71% and 43.64% over the same research time point, respectively (Fig. 3). Mixed ANOVA showed that mean flowering and fruiting activity indices varied significantly within research time points with (F(3,267) = 27.211, p< 0.001, ηp2 = 0.234) and (F(3,267) = 15.692, p< 0.001, ηp2 = 0.150), respectively .
Edaphic and climatic factors affecting flowering and fruiting activity indices: An increase in soil OC content, exchangeable Ca, exchangeable Na, soil EC, total N, exchangeable K and exchangeable Mg at 0-20 sm and 20-40 cm depth were neither increasing nor decreasing flowering and fruiting activity indices of C. procera significantly (p>0.05).
| Table 1: Edaphic conditions in the semi-arid regions of Tharaka and Makueni | ||||||||||||
| Tharaka | Makueni | |||||||||||
| Soil property | Soil depth (cm) | (June-August) 2018 | (March-May) 2019 | (November-September) 2019 | (February-April) 2020 | Mean | (June-August) 2018 | (March-May) 2019 | (November-September) 2019 | (February-April) 2020 | Mean | |
| Soil pH | (0-20) | 7.2 | 7.3 | 7.2 | 7.3 | 7.3 | 6.7 | 6.8 | 6.8 | 6.8 | 6.8 | |
(20-40) | 7.2 | 7.3 | 7.2 | 7.4 | 7.3 | 6.6 | 6.9 | 6.7 | 6.9 | 6.8 | ||
| Soil EC (mS cm–1) | (0-20) | 0.15 | 0.11 | 0.11 | 0.12 | 0.12 | 0.09 | 0.08 | 0.09 | 0.09 | 0.09 | |
(20-40) | 0.15 | 0.14 | 0.13 | 0.15 | 0.14 | 0.11 | 0.11 | 0.11 | 0.12 | 0.11 | ||
| N content (%) | <(0-20) | 0.14 | 0.13 | 0.15 | 0.16 | 0.15 | 0.23 | 0.26 | 0.23 | 0.21 | 0.23 | |
(20-40) | 0.17 | 0.18 | 0.17 | 0.20 | 0.18 | 0.24 | 0.28 | 0.25 | 0.24 | 0.25 | ||
| OC (%) | (0-20) | 2.75 | 3.01 | 3.24 | 2.98 | 3.00 | 3.29 | 3.25 | 3.35 | 2.38 | 3.08 | |
(20-40) | 2.83 | 2.91 | 3.12 | 2.80 | 2.92 | 3.37 | 2.29 | 2.42 | 2.43 | 2.63 | ||
| P (ppm) | (0-20) | 4.53 | 4.79 | 4.90 | 4.90 | 4.78 | 10.58 | 10.50 | 10.71 | 10.77 | 10.64 | |
(20-40) | 4.66 | 4.68 | 5.01 | 5.02 | 4.84 | 10.75 | 10.58 | 10.84 | 10.87 | 10.76 | ||
| K (ppm) | (0-20) | 103.56 | 104.26 | 122.24 | 128.73 | 118.18 | 225.36 | 212.04 | 204.16 | 204.19 | 211.44 | |
(20-40) | 143.08 | 150.86 | 134.87 | 161.12 | 147.48 | 231.74 | 225.47 | 227.01 | 228.58 | 228.20 | ||
| M (ppm) | (0-20) | 79.59 | 81.06 | 76.35 | 74.76 | 77.76 | 105.22 | 109.17 | 94.67 | 105.39 | 103.61 | |
(20-40) | 93.12 | 89.41 | 81.88 | 87.06 | 87.87 | 115.06 | 114.72 | 105.5 | 116.72 | 113.00 | ||
| Ca (ppm) | (0-20) | 1014.00 | 1084.00 | 1042.00 | 1018.00 | 1040.00 | 1333.00 | 1443.00 | 1220.00 | 1369.00 | 1341.00 | |
(20-40) | 1198.00 | 1178.00 | 1040.00 | 1102.00 | 1130.00 | 1535.00 | 1502.00 | 1329.00 | 1527.00 | 1473.00 | ||
| Na (ppm) | (0-20) | 116.00 | 114.00 | 108.00 | 112.00 | 112.5 | 77.00 | 75.00 | 76.00 | 77.00 | 75.00 | |
(20-40) | 88.00 | 87.00 | 86.00 | 85 | 85.00 | 70.00 | 72.00 | 69.00 | 85.00 | 74.00 | ||
| Table 2: Soil edaphic factors affecting flowering activity index of C.procera | |||||||||||
| 95% wald confidence interval | Hypothesis test | 95% wald confidence interval for Exp(B) | |||||||||
| Parameters | B | Lower | Upper | Wald chi-square | df | p-vale | Exp(B) | Lower | Upper | ||
| (Intercept) | 54.130±3.0732 | 48.107 | 60.154 | 310.247 | 1 | <0.001 | 3.224 | 7.8 | 13.31 | ||
| P at (20-40) cm | 0.006±0.0022 | 0.001 | 0.010 | 6.887 | 1 | 0.009 | 1.006 | 1.001 | 1.010 | ||
| Table 3: Climatic factors affecting flowering and fruiting activity indices of C. procera | |||||||||||
| 95% wald confidence interval | Hypothesis test | 95% wald confidence interval for Exp(B) | |||||||||
| Parameters | B | Lower | Upper | Wald chi-square | df | p-vale | Exp(B) | Lower | Upper | ||
| Climatic factors affecting flowering activity index | |||||||||||
| Intercept | 427.69±1.943 | 18.690 | 26.700 | 12.301 | 1 | <0.001 | 5.56 | 2.852 | 3.502 | ||
| Mean monthly rainfall | 0.15±0.095 | 0.342 | 0.434 | 2.576 | 1 | 0.001 | 1.143 | 1.001 | 1.035 | ||
| Mean monthly temperature | 11.70±4.219 | -19.977 | -3.438 | 7.700 | 1 | 0.006 | 0.958 | 0.708 | 0.920 | ||
| Mean monthly wind speed | 21.94±5.070 | -31.886 | -12.011 | 18.740 | 1 | <0.001 | 0.979 | 0.420 | 0.755 | ||
| Climatic factors affecting fruiting activity index | |||||||||||
| Intercept | 89.56±1.616 | 15.121 | 24.007 | 10.607 | 1 | 0.001 | 1.530 | 2.334 | 3.007 | ||
| Mean monthly rainfall | 0.15±0.095 | 0.431 | 0.343 | 4.674 | 1 | 0.022 | 1.144 | 1.001 | 1.031 | ||
| Mean monthly temperature | 11.24±4.148 | -19.376 | -3.114 | 7.347 | 1 | 0.007 | 0.915 | 0.847 | 1.000 | ||
| Mean monthly wind speed | 16.11±5.867 | -27.611 | -4.610 | 7.538 | 1 | 0.006 | 0.948 | 0.821 | 0.999 | ||
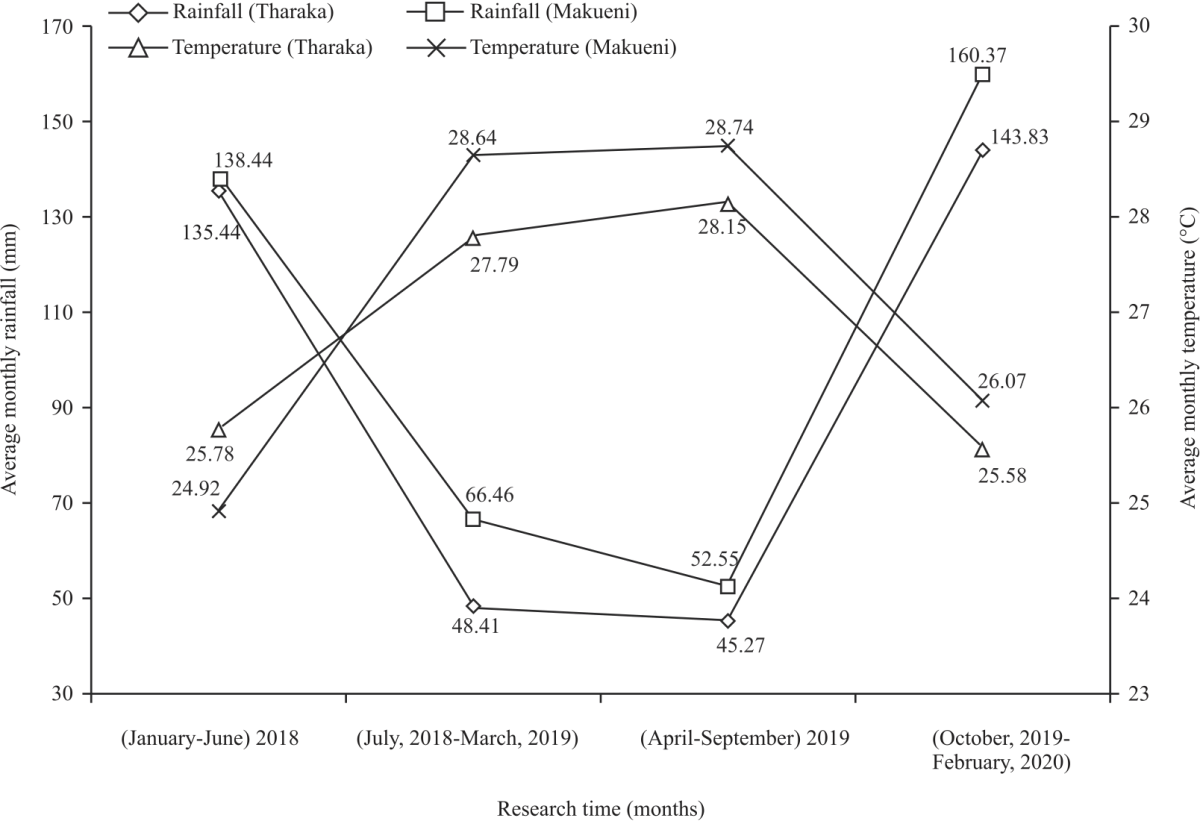 |
| Fig. 1: | Average monthly rainfall and temperature in Tharaka and Makueni |
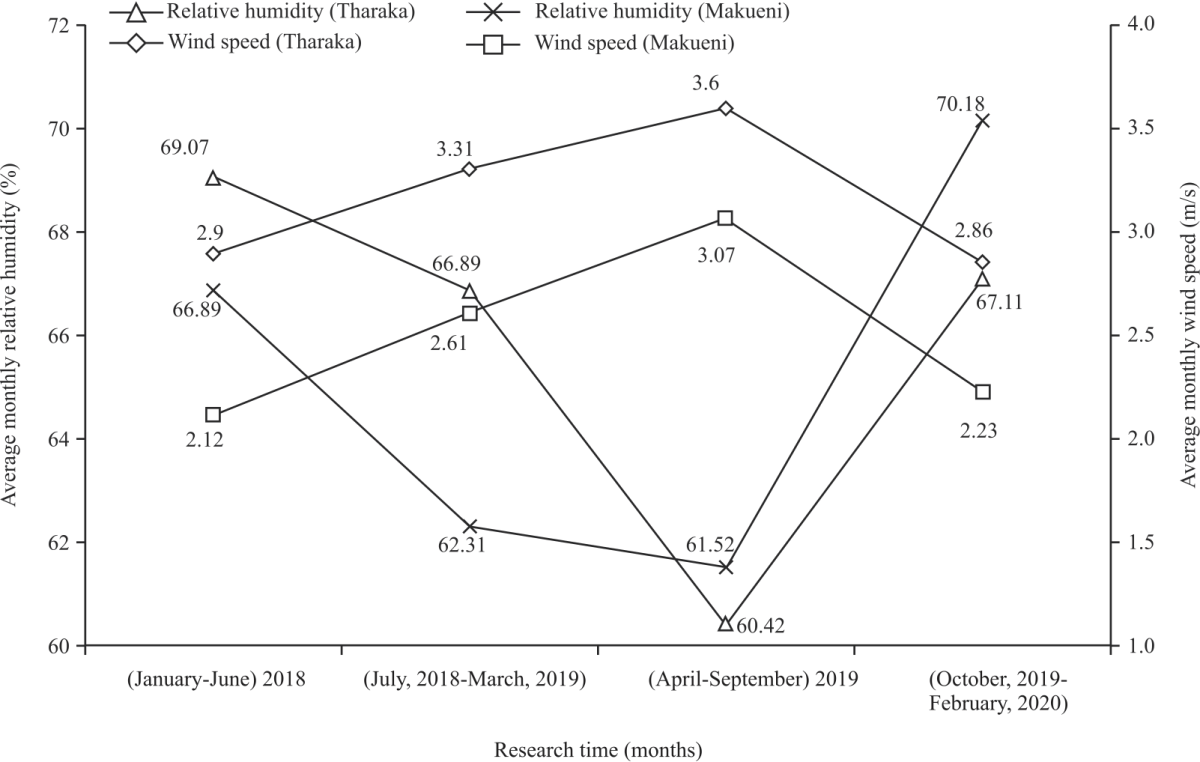 |
| Fig. 2: | Monthly relative humidity and wind speed in Tharaka and Makueni |
However, a unit increase in soil available P increased C. procera’s flowering activity index by 1.006 times in Tharaka and Makueni (Table 2). On the other hand, a unit increase in average monthly rainfall and temperature increased C. procera’s flowering and fruiting activity indices by 1.143 and 1.144 times, respectively (Table 3).
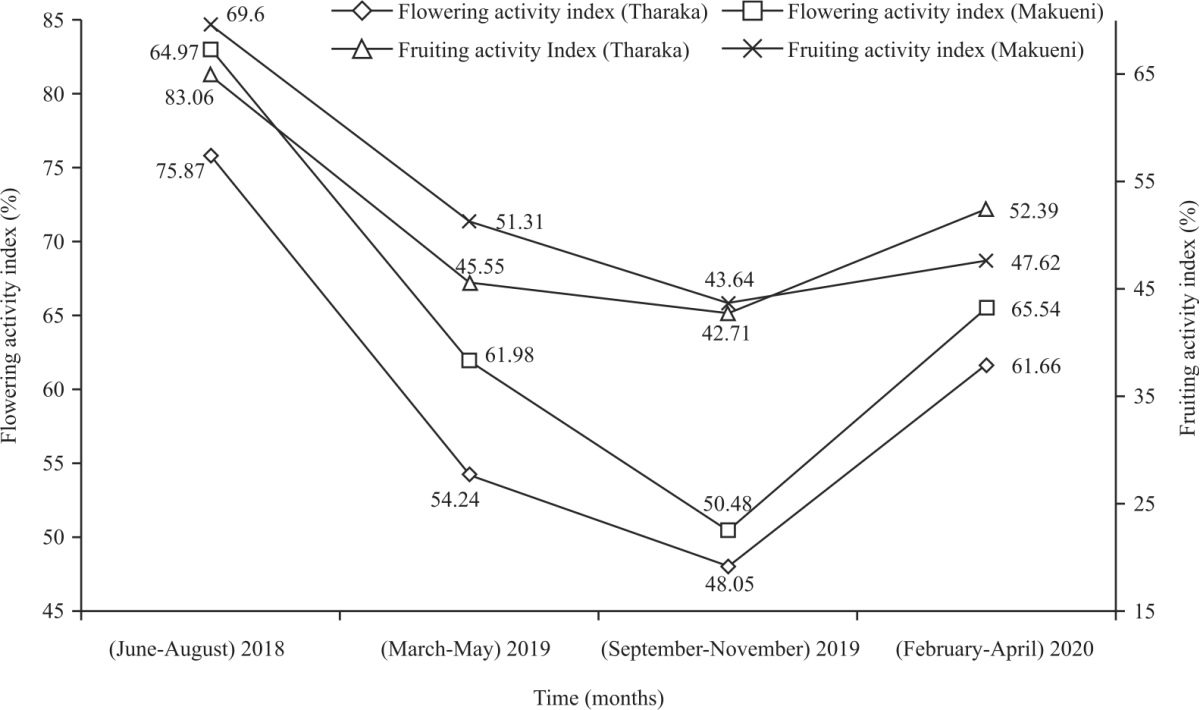 |
| Fig. 3: | Flowering and fruiting activity indices of C. procera |
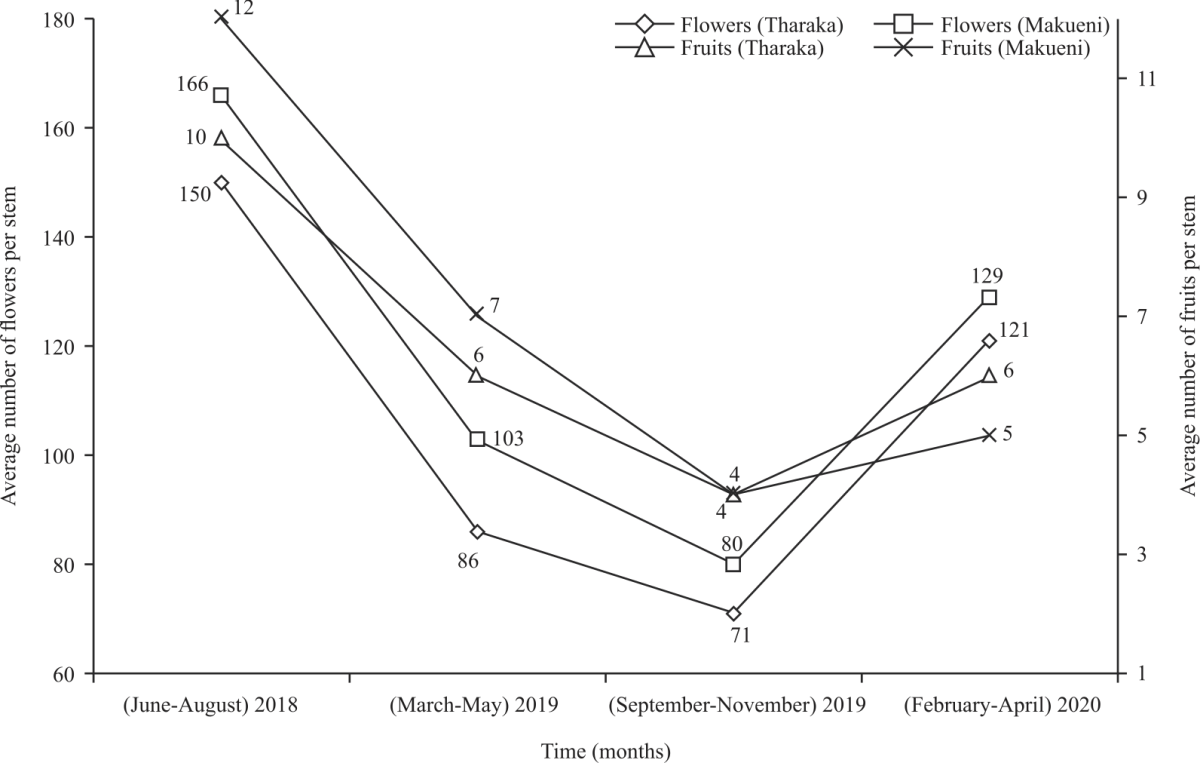 |
| Fig. 4: | Number of flowers and fruits per C. procera stem |
Factors affecting number of flowers and fruits
Number of flowers and fruits: The average number of flowers per flowering C. procera stem in Tharaka and Makueni decreased from 150 and 166 in (June-August, 2018) to 71and 80 in (September-November, 2019), respectively (Fig. 4). The highest number of fruits, 10 in Tharaka and 12 in Makueni were recorded in (June-August, 2018) (Fig. 4). Adjusted Greenhouse-Geisser, showed statistically significant variations in mean number of flowers (F(2.348, 744.261) = 185.420, p<0.001, ηp2 = 0.369) and fruits (F(2.586,778.237) = 269.464, p<0.001, ηp2 = 0.472) per flowering and fruiting C. procera stem within research time points.
Edaphic and climatic factors affecting number of flowers and fruits: It was established that a unit increase in exchangeable Na at 0-20 cm, OC content, available P, exchangeable Ca and exchangeable Na at 20-40 cm significantly increased the number of flowers by 1.002, 1.015, 1.048, 1.002 and 1.005 times, respectively (Table 4).
On the other hand, a unit increase in exchangeable Mg significantly reduced the number of flowers by 0.984 times (Table 4). On fruits, a unit increase in soil exchangeable Na at 0-20 cm, OC content, available P, exchangeable K, exchangeable Mg and exchangeable Na at 20-40 cm significantly increased the number of fruits by 1.005, 1.027, 1.049, 1.044, 1.044 and 1.009 times, respectively (Table 4).
On climatic conditions, a unit increase in monthly average rainfall and relative humidity significantly increased the number of flowers by 1.009 and 1.084 times, respectively, while a unit increase in monthly average temperature and wind speed reduced the number of flowers by 0.792 and 0.844 times, respectively (Table 5). On fruits, a unit increase in mean monthly rainfall, temperature and wind speed significantly increased the number of fruits by 1.056, 1.338 and 1.207 times, respectively (Table 5). Contrary, a unit increase in relative humidity significantly reduced the number of fruits by 0.794 times (Table 5).
Factors affecting phenophase intensity of C. procera in Tharaka and Makueni
Phenophase intensity of C. procera in Tharaka and Makueni: In (June-August, 2018), naturally growing C. procera in the semi-arid regions of Tharaka and Makueni recorded the highest flowering (77.57%) and (79.09%) phenophase intensities, respectively (Fig. 5). Mixed ANOVA showed statistically significant variations in mean flowering (F(3,936) = 67.859, p<0.001, ηp2 = 0.179) and mean fruiting (F(3,93) p<0.001, ηp2 = 0.043) phenophase intensities within research time points.
Edaphic and climatic factors affecting flowering and fruiting phenophase intensities: Parameter estimate (Table 6) indicates that a decrease of 0.999, 0.993, 0.994, 0.992, 0.997 and 0.956 times in C. procera’s flowering phenophase intensity as a result of a unit increase in soil pH, EC, OC, K, Ca and Na, respectively at 0-20 cm soil depth was not statistically significant (p>0.05). In addition, an increase of 1.002, 1.000 and 1001 times in C. procera’s flowering phenophase intensity as a result of a unit increase in soil N, P and Mg at 0-20 cm soil depth was not statistically significant (p>0.05) (Table 6). At 20-40 cm soil depth, a unit increase in soil pH, EC, N, P and Mg caused no statistically significant increase in C. procera’s flowering phenophase intensity of 1.004, 1.003, 1.005, 1.000 and 1.009 times, respectively (Table 6).
| Table 4: Edaphic factors affecting number of flowers and fruits produced by C. procera | |||||||||||
| 95% wald confidence interval | Hypothesis test | 95% wald confidence interval for Exp(B) | |||||||||
| Parameters | B | Lower | Upper | Wald chi-square | df | p-vale | Exp(B) | Lower | Upper | ||
| Estimates of edaphic factors affecting number of flowers | |||||||||||
| Intercept | 4.194±0.077 | 4.040 | 4.345 | 96.638 | 1 | <0.001 | 2.171 | 6.817 | 7.065 | ||
| Na at (0-20) cm | 0.002±0.000 | 0.002 | 0.003 | 65.027 | 1 | <0.001 | 1.002 | 1.002 | 1.003 | ||
| OC at (20-40) cm | 0.015±0.021 | 0.200 | 0.217 | 55.145 | 1 | <0.001 | 1.015 | 1.181 | 1.270 | ||
| P at (20-40) cm | 0.047±0.009 | 0.028 | 0.065 | 23.557 | 1 | <0.001 | 1.048 | 1.028 | 1.068 | ||
| Mg at (20-40) cm | 0.016±0.003 | -0.022 | -0.010 | 27.988 | 1 | <0.001 | 0.984 | 0.979 | 0.990 | ||
| Ca at (20-40) cm | 0.002±0.000 | 0.001 | 0.002 | 51.748 | 1 | <0.001 | 1.002 | 1.001 | 1.002 | ||
| Na at (20-40) cm | 0.005±0.000 | 0.006 | 0.003 | 51.899 | 1 | <0.001 | 1.005 | 1.094 | 1.097 | ||
| Estimates of edaphic factors affecting number of fruits | |||||||||||
| Intercept | 3.384±0.2426 | 2.909 | 3.859 | 94.621 | 1 | <0.001 | 2.488 | 1.330 | 4.438 | ||
| Na at (0-20) cm | 0.005±0.000 | 0.007 | 0.004 | 33.667 | 1 | <0.001 | 1.005 | 1.007 | 1.010 | ||
| OC at (20-40) cm | 0.027±0.032 | 0.206 | 0.334 | 67.819 | 1 | <0.001 | 1.027 | 1.228 | 1.397 | ||
| P at (20-40) cm | 0.050±0.016 | 0.082 | 0.019 | 9.731 | 1 | 0.002 | 1.049 | 1.079 | 1.099 | ||
| K at (20-40) cm | 0.001±0.000 | 0.003 | 0.000 | 4.646 | 1 | 0.031 | 1.001 | 1.000 | 1.003 | ||
| Mg at (20-40) cm | 0.043±0.007 | 0.029 | 0.058 | 33.919 | 1 | <0.001 | 1.044 | 1.029 | 1.060 | ||
| Ca at (20-40) cm | 0.004±0.000 | -0.005 | -0.003 | 60.330 | 1 | <0.001 | 0.996 | 0.995 | 0.997 | ||
| Na at (20-40) cm | 0.009±0.002 | 0.005 | 0.013 | 21.674 | 1 | <0.001 | 1.009 | 1.005 | 1.013 | ||
| Table 5: Climatic factors affecting number of flowers and fruits produced by C. procera | |||||||||||
| 95% wald confidence interval | Hypothesis test | 95% wald confidenceinterval for Exp(B) | |||||||||
| Parameters | B | Lower | Upper | Wald chi-square | df | p-vale | Exp(B) | Lower | Upper | ||
| Climatic factors affecting number of flowers | |||||||||||
| Intercept | 19.514±1.553 | 16.468 | 22.559 | 57.746 | 1 | <0.001 | 2.983 | 1.419 | 2.682 | ||
| Mean monthly rainfall | 0.009±0.001 | 0.011 | 0.007 | 81.447 | 1 | <0.001 | 1.009 | 1.021 | 1.093 | ||
| Mean monthly temperature | 0.709±0.053 | -0.812 | -0.606 | 82.002 | 1 | <0.001 | 0.792 | 0.444 | 0.546 | ||
| Mean monthly wind speed | 0.813±0.056 | -0.923 | -0.702 | 107.596 | 1 | <0.001 | 0.844 | 0.397 | 0.496 | ||
| Monthly relative humidity | 0.080±0.009 | 0.063 | 0.097 | 86.797 | 1 | <0.001 | 1.084 | 1.066 | 1.102 | ||
| Climatic factors affecting number of fruits | |||||||||||
| Intercept | 5.536±2.286 | 47.050 | 56.018 | 58.143 | 1 | <0.001 | 2.148 | 4.690 | 3.664 | ||
| Mean monthly rainfall | 0.054±0.001 | 0.052 | 0.056 | 26.751 | 1 | <0.001 | 1.056 | 1.053 | 1.058 | ||
| Mean monthly temperature | 0.201±0.059 | 2.085 | 2.318 | 77.953 | 1 | <0.001 | 1.338 | 8.046 | 10.152 | ||
| Mean monthly wind speed | 0.129±0.087 | 3.158 | 3.500 | 45.911 | 1 | <0.001 | 1.207 | 23.518 | 33.115 | ||
| Monthly relative humidity | 0.231±0.010 | -0.250 | -0.211 | 53.798 | 1 | <0.001 | 0.794 | 0.779 | 0.809 | ||
| Table 6: Edaphic factors affecting flowering phenophase intensity of C. procera | |||||||||||
| 95% wald confidence interval | Hypothesis test | 95% wald confidence interval for Exp(B) | |||||||||
| Parameter | B | Lower | Upper | Wald chi-square | df | p-vale | Exp(B) | Lower | Upper | ||
| Intercept | 66.555±1.473 | 5.827 | 7.283 | 147.849 | 1 | <0.001 | 1.080 | 1.759 | 3.660 | ||
| pH at (0-20) cm | 0.472±.507 | -1.467 | 0.524 | 0.862 | 1 | 0.353 | 0.999 | 1.231 | 1.689 | ||
| EC at (0-20) cm | 3.403±2.891 | -2.790 | 1.983 | 0.118 | 1 | 0.731 | 0.993 | 0.266 | 0.873 | ||
| N at (0-20) cm | 9.697±2.614 | -3.268 | 2.662 | 2.149 | 1 | 0.143 | 1.002 | 0.038 | 0.069 | ||
| OC at (0-20) cm | 0.557±0.711 | -1.952 | 0.839 | 0.612 | 1 | 0.434 | 0.994 | 0.142 | 0.313 | ||
| P at (0-20) cm | 0.573±0.462 | -0.333 | 1.480 | 1.536 | 1 | 0.215 | 1.000 | 0.717 | 1.392 | ||
| K at (0-20) cm | 0.018±0.009 | -0.037 | 0.001 | 3.579 | 1 | 0.059 | 0.992 | 0.963 | 1.001 | ||
| Mg at (0-20) cm | 0.129±0.052 | 0.027 | 0.231 | 4.118 | 1 | 0.053 | 1.001 | 1.027 | 1.260 | ||
| Ca at (0-20) cm | 0.013±0.004 | -0.021 | -0.005 | 1.219 | 1 | 0.231 | 0.997 | 0.979 | 0.995 | ||
| Na at (0-20) cm | 0.015±0.015 | -0.045 | 0.015 | 0.966 | 1 | 0.326 | 0.995 | 0.956 | 1.015 | ||
| pH at (20-40) cm | 0.500±0.596 | -0.668 | 1.669 | 0.705 | 1 | 0.401 | 1.004 | 0.313 | 0.506 | ||
| EC at (20-40) cm | 10.877±2.311 | -1.494 | 2.248 | 2.970 | 1 | 0.085 | 1.003 | 0.225 | 0.248 | ||
| N at (20-40) cm | 11.991±1.748 | -2.298 | -2.685 | 0.377 | 1 | 0.701 | 1.005 | 0.628 | 0.680 | ||
| OC at (20-40) cm | 0.468±0.781 | -2.000 | 1.064 | 0.358 | 1 | 0.550 | 0.996 | 0.135 | 0.899 | ||
| P at (20-40) cm | 0.903±0.478 | -0.035 | 1.841 | 3.557 | 1 | 0.059 | 1.000 | 0.965 | 1.305 | ||
| K at (20-40) cm | 0.010±0.008 | -0.027 | 0.007 | 1.320 | 1 | 0.251 | 0.999 | 0.973 | 1.007 | ||
| Mg at (20-40) cm | 0.090±0.104 | -0.115 | 0.295 | 0.738 | 1 | 0.390 | 1.009 | 0.891 | 1.343 | ||
| Ca at (20-40) cm | 0.009±0.008 | -0.025 | 0.008 | 1.081 | 1 | 0.298 | 0.998 | 0.975 | 1.008 | ||
| Na at (20-40) cm | 0.001±0.024 | -0.049 | 0.048 | 0.000 | 1 | 0.984 | 0.999 | 0.952 | 1.049 | ||
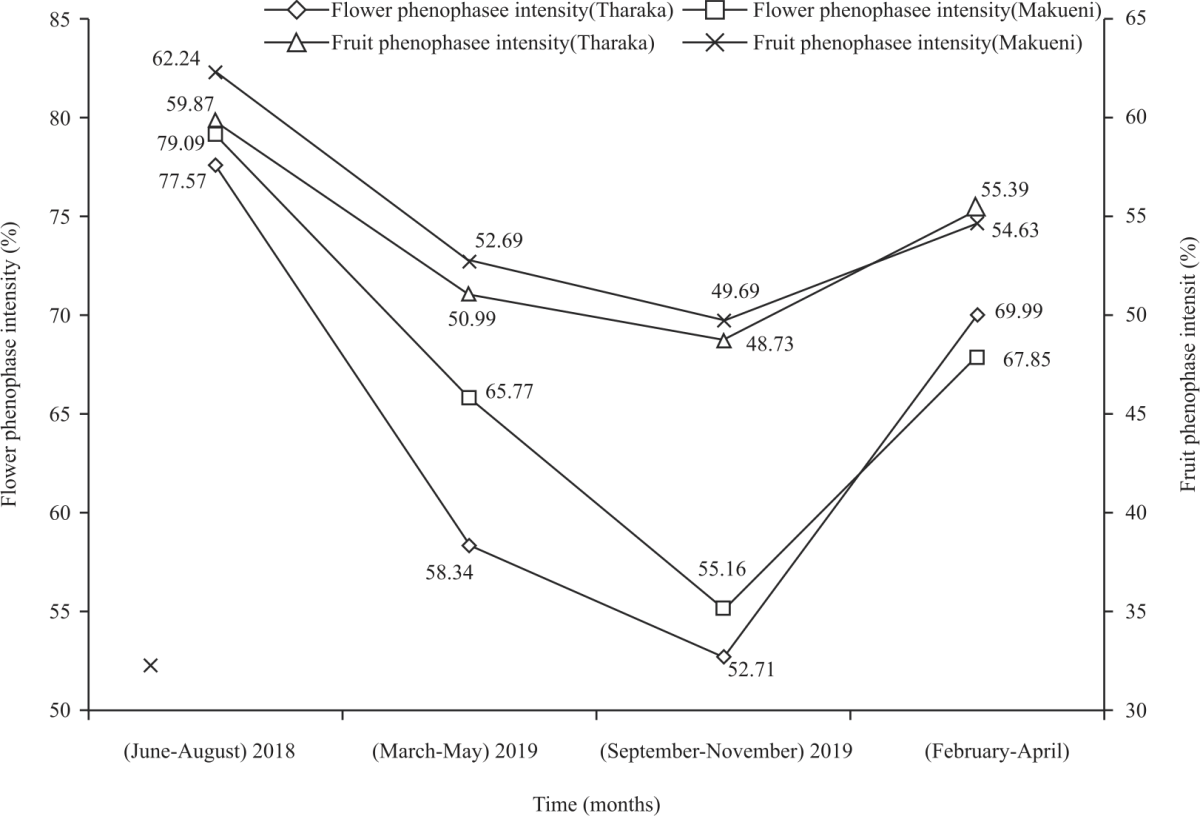 |
| Fig. 5: | Flowering and fruiting phenophase intensities of C. procera in Tharaka and Makueni |
| Table 7: Edaphic factors affecting fruiting phenophase intensity of C. procera | |||||||||||
| 95% wald confidence interval | Hypothesis test | 95% wald confidence interval for Exp(B) | |||||||||
| Parameters | B | Lower | Upper | Wald chi-square | df | p-vale | Exp(B) | Lower | Upper | ||
| Intercept | 2.869±2.428 | 3.270 | 5.469 | 44.470 | 1 | <0.001 | 1.414 | 1.399 | 2.302 | ||
| pH at (0-20) cm | 0.037±0.615 | -1.170 | 1.244 | 0.004 | 1 | 0.952 | 1.007 | 0.310 | 3.469 | ||
| EC at (0-20) cm | -4.382±3.548 | -59.096 | -1.668 | 7.887 | 1 | 0.060 | 1.005 | 2.163 | 3.889 | ||
| N at (0-20) cm | 5.660±2.581 | -9.201 | 2.520 | 0.557 | 1 | 0.455 | 1.009 | 0.000 | 0.081 | ||
| OC at (0-20) cm | -1.872±0.906 | -3.649 | -0.096 | 4.269 | 1 | 0.059 | 0.994 | 0.026 | 0.908 | ||
| P at (0-20) cm | 0.061±0.619 | -1.153 | 1.276 | 0.010 | 1 | 0.921 | 1.003 | 0.316 | 3.583 | ||
| K at (0-20) cm | 0.001±0.009 | -0.018 | 0.019 | 0.007 | 1 | 0.932 | 1.001 | 0.983 | 1.019 | ||
| Mg at (0-20) cm | 0.075±0.059 | -0.042 | 0.191 | 1.565 | 1 | 0.211 | 1.007 | 0.959 | 1.211 | ||
| Ca at (0-20) cm | 0-.002±0.004 | -0.011 | 0.007 | 0.168 | 1 | 0.682 | 0.998 | 0.989 | 1.007 | ||
| Na at (0-20) cm | 0.058±0.018 | 0.022 | 0.095 | 7.742 | 1 | 0.054 | 1.006 | 1.022 | 1.099 | ||
| pH at (20-40) cm | 0.298±0.739 | -1.151 | 1.748 | 0.163 | 1 | 0.687 | 1.018 | 0.316 | 5.744 | ||
| EC at (20-40) cm | 2.651±3.016 | -9.141 | 14.442 | 0.194 | 1 | 0.660 | 1.000 | 0.000 | 1.719 | ||
| N at (20-40) cm | 2.209±3.136 | -7.857 | 12.276 | 0.185 | 1 | 0.667 | 1.001 | 0.000 | 1.446 | ||
| OC at (20-40) cm | 5.512±0.724 | -6.931 | -4.092 | 7.921 | 1 | 0.052 | 1.004 | 0.001 | 0.017 | ||
| P at (20-40) cm | 1.696±0.420 | 0.873 | 2.519 | 6.305 | 1 | 0.059 | 1.004 | 2.394 | 12.421 | ||
| K at (20-40) cm | 0.009±0.010 | -0.012 | 0.030 | 0.761 | 1 | 0.383 | 1.009 | 0.988 | 1.031 | ||
| Mg at (20-40) cm | 0.609±0.123 | -0.852 | -0.367 | 4.303 | 1 | 0.082 | 0.994 | 0.427 | 0.693 | ||
| Ca at (20-40) cm | 0.056±0.009 | 0.037 | 0.074 | 3.763 | 1 | 0.097 | 1.007 | 1.038 | 1.077 | ||
| Na at (20-40) cm | 0.118±0.025 | -0.167 | -0.068 | 2.769 | 1 | 0.105 | 0.999 | 0.846 | 0.934 | ||
However, a unit increase in soil OC, K, Ca and Na at (20-40) cm soil depth led to a statistically no significant decrease of 0.996, 0.999, 0.998 and 0.999 times in C. procera’s flowering phenophase intensity (Table 6).
On fruiting, an increase of 1.007, 1.005, 1.009, 1.003, 1.001, 1.007and 1.006 times in C. procera’s fruiting phenophase intensities as a result of a unit increase in soil pH, EC, N, P, K, Mg and Na, respectively at 0-20 cm soil depth was not statistically significant (p>0.05) (Table 7). A unit increase in soil OC and Ca at 0-20 cm soil depth led to a statistically no significant decrease (p>0.05) in C. procera’s fruiting phenophase intensity by 0.994 and 0.998 times, respectively.
| Table 8: Climatic factors affecting phenophase intensities of C. procera | |||||||||||
| 95% wald confidence interval | Hypothesis test | 95% wald confidence interval for Exp(B) | |||||||||
| Parameters | B | Lower | Upper | Wald chi-square | df | p-vale | Exp(B) | Lower | Upper | ||
| Climatic factors affecting flowering phenophase intensity | |||||||||||
| Intercept | 15.36±2.289 | 18.643 | 19.096 | 49.888 | 1 | <0.001 | 2.017 | 1.524 | 2.668 | ||
| Mean monthly rainfall | 0.131±0.0329 | 0.067 | 0.196 | 15.930 | 1 | <0.001 | 1.014 | 1.069 | 1.216 | ||
| Mean monthly temperature (°C/month) | 3.649±0.8739 | -5.362 | -1.936 | 17.435 | 1 | <0.001 | 0.981 | 0.005 | 0.144 | ||
| Climatic factors affecting fruiting phenophase intensity | |||||||||||
| Intercept | 38.296±2.596 | 45.610 | 60.982 | 73.952 | 1 | <0.001 | 1.014 | 1.151 | 3.142 | ||
| Mean monthly rainfall (mm/month) | 0.443±0.0427 | 0.359 | 0.527 | 107.618 | 1 | <0.001 | 1.012 | 1.591 | 1.698 | ||
| Mean monthly temperature (°C/month) | 1.061±1.719 | -25.430 | -18.691 | 64.645 | 1 | <0.001 | 0.965 | 0.987 | 1.000 | ||
| Mean monthly wind speed (m/s) | 1.440±3.071 | -36.458 | -24.422 | 98.278 | 1 | <0.001 | 0.987 | 0.841 | 0.947 | ||
At 20-40 cm soil depth, soil pH, EC, N, OC, P, K and Ca led to a statistically no significant increase (p>0.05) in C. procera’s fruiting phenophase intensity by 1.018, 1.000, 1.001, 1.004, 1.004, 1.009 and 1.007 times, respectively (Table 7). On the other hand, a statistically no significant decrease (p>0.05) of 0.994 and 0.999 times in C. procera’s fruiting phenophase intensity as a result of a unit increase in soil Mg and Na, respectively at 20-40 cm soil depth (Table 7).
On the other hand, a unit increase in monthly average rainfall increased flowering and fruiting phenophase intensities by 1.014 and 1.012 times, respectively (Table 8). Contrary, a unit increase in monthly average temperature significantly reduced flowering and fruiting phenophase intensities by 0.981 and 0.965 times, respectively (Table 7). A unit increase in monthly average wind speed decreased fruiting phenophase intensity by 0.987 (Table 8).
Soils from Tharaka and Makueni were deficient in available P. This concur with Koala20 that over 65.1% of soil samples from semi-arid regions are acutely deficient in total phosphorus. This deficiency in total phosphorus is as a result of imbalance in a number of biological and biochemical processes that are significantly influenced by soil organic matter, soil texture, biotic factors and abiotic characteristics of the region21,22. The study showed that the highest and lowest average monthly rainfall recorded was 160.37 mm and 52.55 mm per month, respectively for Makueni and 143.83 mm and 45.27 mm per month for Tharaka. These concur with Camberlin et al.16 that semi-arid regions of Kenya receive low, varied and unreliable rainfall. This is not different from other semi-arid regions which experience greater inter-and intra-annual rainfall variation23. Average monthly temperature ranged from 25.78-28.15°C in Tharaka and 24.92-28.74°C in Makueni. These high temperatures may be attributed to high solar radiations, low cloud cover and their proximity to the equator24. Wind speed variations were as a result of variations in temperature, cloud cover and earth’s revolution. According to Wooten25 cloud cover affects temperature which creates pressure difference between places that eventually affects wind speed. There was noticeable relationship between edaphic factors with activity indices, number of flowers and fruits and phenophase intensities. This noticeable relationship between edaphic factors with phenological traits indicates that though C. procera can tolerate soils with low nutrient content due to its intensive root system that ensure reaching nutrients and moisture beyond 40 cm depth26, soil conditions have slight impacts on the shrub’s phenology. Adequate availability of soil N content, available P, exchangeable Ca and OC content enhances the development of plant leaves and increases plant’s tolerance to other environmental stresses27,28. Exchangeable Ca plays an important role in reducing the adverse effects of drought stress in plant crops29. A healthy plant with improved photosynthesis ensures availability of carbohydrates for plants’ flowering and fruiting30. Deficiency of soil nutrients like available P, exchangeable Ca, available K and exchangeable Mg leads to stunted growth as a result of reduced photosynthesis and lower resistance to diseases31,32. This condition leads to aborted flowers and fruits in plants by impairing female reproductive organs and reduces pollen grain formation and viability especially under high saline and drought conditions33-35. Exchangeable Mg is essential for chlorophyll a and b in light energy and synthesis of both in plants36. Soil pH and EC were not associated with phenological traits of C. procera. This was because the shrub has adaptive avoidance mechanism to salinity and pH stresses37,38. According to Fekry et al.39 high salinity inhibit growth of plants like date palm hence the need to alleviate its effects on growth and fruiting. Gulzar et al.40 recommends that a combination of nitrogen and phosphorus fertilizers can improve growth and productivity of plants that are salt stressed. The significant association between phenological traits with average monthly rainfall and temperature concur with studies like Moore and Lauenroth14 that temperature and rainfall influences phenological events especially in ASALs. This is because phenology development requires optimal temperature and adequate moisture that is influenced by rainfall41. Temperature and precipitation influences pollen and ovule viability and affects visitation by pollinators42-44. Extreme temperature and precipitation reduces photosynthetic activity of C. procera as they affect opening and closing of plant’s stomata, hence reducing availability of flowering and fruiting energy in plants. In addition, extreme environmental stresses including high temperature and low rainfall makes plants susceptible to pathogens and diseases45. However, the association was weak with low odd ratios because other factors like plant size especially in terms of crown diameter and genetic composition influences phenological traits like number of flowers and fruits46. Large crowns provide more space for flowers and fruits. In terms of genetics, though C. procera can withstand harsh climatic conditions like high temperatures and low rainfall5, the shrub experiences low fertility rates, high drop of floral buds and flower abortion after anthesis regardless of prevailing conditions47. Wind speed was slightly associated with phenology of C. procera negatively. High wind speeds causes traumatic flower and fruit fall before maturity. It also discourages flower visitation by pollinators by desiccating flower parts, making them unattractive, hence lowering fertilization rates48. However, high wind speed increases the chances of self-pollination assisted by wind48. Relative humidity affects phenology of plants indirectly by affecting pollination, photosynthesis and disease occurrence49. Low relative humidity increases transpiration, leading to water deficit for photosynthesis50. However, high relative humidity impedes dispersal of pollen grains from anthers and increase disease instances by favouring fungal growth49. Phenological traits of C. procera peaked in (June-August, 2018 and troughed in (September-November, 2019) in Tharaka and Makueni. This concur with Sobrinho13, Paradiso and Pascale51 and Moustafa and Sarah26 that C. procera show peak and low phenology traits at different times of the year depending on prevailing environmental conditions like precipitation and temperature. Semi-arid regions of Tharaka and Makueni in Kenya experience low monthly rainfalls, medium temperatures and wind speed that vary from time to time. Soils in Tharaka were deficient in available P and exchangeable K while those of Makueni were deficient in available P. Flowering activity index of C. procera requires adequate supply of soil available P while an increasing number of flowers per stem requires optimal supply of soil exchangeable Na, OC content, available P and exchangeable Mg. Optimal fruit production of C. procera fruits requires adequate supply of soil exchangeable Na, OC content, available P, available K, exchangeable Mg and exchangeable Ca. Enhancing phenological traits of C. procera requires optimal rains, temperatures and wind speed. This study discovered that phenological traits of C. procera are influenced by both edaphic and climatic factors. Soil properties such as soil exchangeable Na, OC content, available P and exchangeable Mg increased the production of flowers and fruits. Similarly, average monthly rainfall and temperature are critical factors influencing phenological traits. This information is important when introducing the plant from the wild to on farm cultivation. This study will help the researchers to uncover the critical areas in determining reproductive successes of the plant in its environment for the purpose of domestication. Thus a new theory on the success of C. procera domestication may be arrived at for a sustainable supply of fiber for the growing textile industry in Kenya. We thank all farm owners that voluntarily allowed access to their farms for research. This research was financially supported by German Academic Exchange Service (DAAD) under ICRAF- DAAD collaboration (DAAD-1157).DISCUSSION
CONCLUSION
SIGNIFICANCE STATEMENT
ACKNOWLEDGMENT
REFERENCES
- Huho, J.M. and E.M. Mugalavai, 2010. The effects of droughts on food security in Kenya. Int. J. Clim. Change: Impacts Responses, 2: 61-72.
CrossRefDirect Link - Simons, A.J. and R.R.B. Leakey, 2004. Tree domestication in tropical agroforestry. Agroforestry Syst., 61: 167-181.
CrossRef - Jamnadass, R., D.A. Ofori, I.K. Dawson, Z. Tchoundjeu, S. McMullin, P.S. Hendre and L. Graudal, 2019. Enhancing agroforestry systems through tree domestication. In: Sustainable development through trees on farms: Agroforestry in its fifth decade. Noordwijk, M.V. (Ed.)., World Agroforestry (ICRAF) Southeast Asia Regional Program, Bogor, Indonesia, ISBN: 978-602-5894-03-9 pp: 45-59.
Direct Link - Bairagi, S.M., G. Prashant and R. Gilhotra, 2018. Pharmacology of natural products: A recent approach on Calotropisgigantea and Calotropisprocera. Ars Pharm., 59: 37-44.
CrossRefDirect Link - Al-Sodany, Y.M., N.S. Al-Juaid and A.A.K. Kahil, 2016. Ecology of invasive species in Saudi Arabia, Calotropis procera (Ait) W.T. Ait.: Floristic composition and associated plant communities. Int. J. Ecotoxicol. Ecobiol., 1: 127-140.
CrossRefDirect Link - Abeysinghe, P.D., 2018. Review article on Calotropis (Wara): Is it a miracle shrub or just a plant? J. Univ. Ruhuna, 6: 55-60.
CrossRefDirect Link - Menge, E.O., M.L. Greenfield, C.A. McConchie, S.M. Bellairs and M.J. Lawes, 2017. Density-dependent reproduction and pollen limitation in an invasive milkweed, Calotropis procera (Ait.) R. Br. (Apocynaceae). Austral Ecol., 42: 61-71.
CrossRefDirect Link - Bufebo, B., T. Tessema and R. Fisshaie, 2016. Spatial distribution and abundance of invasive alien plant species in Gamo Gofa zone, Ethiopia. Int. J. Innovat. Res. Dev., 5: 23-33.
Direct Link - Bakhsh, A., E. Anayol, S.F. Özcan, T. Hussain, M. Aasim, K.M. Khawar and S. Özcan, 2015. An insight into cotton genetic engineering (Gossypium hirsutum L.): current endeavors and prospects. Acta Physiol. Plant, Vol. 37.
CrossRefDirect Link - Akhtar, S., A.A. Shahid, A.Q. Rao, K.S. Bajwa, A. Muzaffar, Ayesha Latif, Tayyab Husnain 2014. Genetic effects of Calotropisprocera CpTIP1 gene on fiber quality in cotton (Gossypiumhirsutum). Adv. life sci., 1: 223-230.
Direct Link - El-Tantawy, H., 2000. Flowering and fruiting eco-physiology of Calotropis procera (Ait.) W.T. Ait and importance of gas in Fruit dehiscence. Taeckholmia, 20: 69-80.
CrossRefDirect Link - Sobrinho, M.S., G.M. Tabatinga, I.C. Machado and A.V. Lopes, 2013. Reproductive phenological pattern of Calotropis procera (Apocynaceae), an invasive species in Brazil: Annual in native areas; continuous in invaded areas of Caatinga. Acta Bot. Bras., 27: 456-459.
CrossRefDirect Link - Moore, L.M. and W.K. Lauenroth, 2017. Differential effects of temperature and precipitation on early- vs. late-flowering species. Ecosphere, Vol. 8.
CrossRefDirect Link - Cloudsley-Thompson, J.L., 2009. Rangelands management and ecology in east Africa. London: Hodder and Stoughton. Envir. Conserv., 5: 316-316.
CrossRefDirect Link - Camberlin, P., J. Boyard-Micheau, N. Philippon, C. Baron, C. Leclerc and C. Mwongera, 2014. Climatic gradients along the windward slopes of Mount Kenya and their implication for crop risks. Part 1: climate variability. Int. J. Climatol., 34: 2136-2152.
CrossRefDirect Link - Kraska, M., 2010. Repeated measures design. In: Encyclopedia of research design. Salkind, N.J. (Ed.)., Sage Publications Inc., Thousand Oaks, pp: 1244-1248.
CrossRefDirect Link - Dell, R.B., S. Holleran and R. Ramakrishnan, 2002. Sample size determination. ILAR J., 43: 207-213.
CrossRefDirect Link - Udelhoven, T., C. Emmerling and T. Jarmer, 2003. Quantitative analysis of soil chemical properties with diffuse reflectance spectrometry and partial least-square regression: A feasibility study. Plant Soil, 251: 319-329.
CrossRefDirect Link - Koala, S., J.R. Sims, H. El-Attar and M. El-Halfawi, 2011. Phosphorus deficiency in the semi-arid tropics and implications for grain legume production. In: World crops: Cool season food legumes. Current Plant Science and Biotechnology in Agriculture, Summerfield, R.J. (Ed.)., Springer, Dordrecht, Berlin/Heidelberg, Germany, ISBN: 978-94-009-2764-3, Pages: 1230.
CrossRefDirect Link - Suñer, L. and J. Galantini, 2015. Texture influence on soil phosphorus content and distribution in semiarid pampean grasslands. Int. J. Plant Soil Sci., 7: 109-120.
CrossRefDirect Link - Bhat, N.A., A. Riar, A. Ramesh, S. Iqbal, M.P. Sharma, S.K. Sharma and G.S. Bhullar, 2017. Soil biological activity contributing to phosphorus availability in Vertisols under long-term organic and conventional agricultural management. Front. Plant Sci. Vol. 8.
CrossRefDirect Link - Pilgrim, D.H., T.G. Chapman and D.G. Doran, 1998. Problems of rainfall-runoff modelling in arid and semiarid regions. Hydrolo. Sci. J., 33: 379-400.
CrossRefDirect Link - Scholes, R.J., 2020. The future of semi-arid regions: A weak fabric unravels. Climate, Vol. 8.
CrossRefDirect Link - Wooten, R.D., 2011. Statistical analysis of the relationship between wind speed, pressure and temperature. J. Applied Sci., 11: 2712-2722.
CrossRefDirect Link - Moustafa, A.R.A. and S.Q. Sarah, 2017. Population ecology and economic importance of Calotropis procera as an exotic medicinal plant. J. Ecol. Nat. Res., 1: 1-11.
CrossRefDirect Link - Sebetha, E. and S. Modisapudi, 2019. Effect of nitrogen fertilizer source, soil type and season on growth performance of two sorghum cultivars. Asian J. Plant Sci., 18: 175-180.
CrossRefDirect Link - Razaq, M., P. Zhang, H.L. Shen and Salahuddin, 2017. Influence of nitrogen and phosphorous on the growth and root morphology of Acer mono. PLoS ONE, Vol. 12.
CrossRefDirect Link - Sadak, M.S., M.A.M. El-Enany, B.A. Bakry, M.M.S. Abdallah and H.M.S. El-Bassiouny, 2020. Signal molecules improving growth, yield and biochemical aspects of wheat cultivars under water stress. Asian J. Plant Sci., 19: 35-53.
CrossRefDirect Link - Fischer, G., P.J. Almanza-Merchán and F. Ramírez, 2012. Source-sink relationships in fruit species: A review. Rev. Colomb. Cienc. Hortic., 6: 238-253.
CrossRefDirect Link - Guo, J., Y. Jia, H. Chen, L. Zhang and J. Yang et al., 2019. Growth, photosynthesis, and nutrient uptake in wheat are affected by differences in nitrogen levels and forms and potassium supply. Sci. Rep., Vol. 9.
CrossRefDirect Link - Cruz, F.J.R., R. de Mello Prado, G. Felisberto, Á.S. Santos and R.F. Barreto, 2017. Potassium nutrition in fruits and vegetables and food safety through hydroponic system. IntechOpen3.
CrossRefDirect Link - Hasanuzzaman, M., M.H.M. Bhuyan, K. Nahar, M.S. Hossain and J. Al Mahmud et al., 2018. Potassium: A vital regulator of plant responses and tolerance to abiotic stresses. Agronomy, Vol. 8.
CrossRefDirect Link - Erel, R., U. Yermiyahu, H. Yasuor, D.C. Chamus, A. Schwartz, A. Ben-Gal and A. Dag, 2016. Phosphorous nutritional level, carbohydrate reserves and flower quality in olives. PLoS ONE, Vol. 11.
CrossRefDirect Link - Teixeira, L.A.J., J.A. Quaggio, H. Cantarella and E.V. Mellis, 2011. Potassium fertilization for pineapple: Effects on plant growth and fruit yield. Rev. Bras. Fruticult., 33: 618-626.
CrossRefDirect Link - Hasanah, Y., L. Mawarni, H. Hanum, F.E. Sitepu and B. Ardi, 2020. Effect of kieserite on physiological characteristics of soybean varieties under dryland conditions. Asian J. Plant Sci., 19: 21-25.
CrossRefDirect Link - Ibrahim, A., 2013. Tolerance and avoidance responses to salinity and water stresses in Calotropis procera and Suaedaaegyptiaca. Turk J. Agric. Fot., 37: 352-360.
CrossRefDirect Link - Leal, L.C., M.V. Meiado and A.V. Lopes and Leal, I.R., 2013. Germination responses of the invasive Calotropis procera (Ait.) R. Br. (Apocynaceae): comparisons with seeds from two ecosystems in northeastern Brazil. An. Acad. Bras. Ciênc., 85: 1025-1034.
CrossRefDirect Link - Fekry, W.M.E., M.A. Rashad and A.H. Alalaf, 2020. Attempts to improve the growth and fruiting of barhi date-palms under salinity stress. Asian J. Plant Sci., 19: 146-151.
CrossRefDirect Link - Gulzar, N., M.Y. Mir and S. Hamid, 2019. Effect of nitrogen and phosphorus on the growth and redox homeostasis of salt-stressed mustard plants Asian J. Plant Sci., 18: 52-59.
CrossRefDirect Link - Moore, L.M., W.K. Lauenroth, D.M. Bell and D.R. Schlaepfer, 2015. Soil water and temperature explain canopy phenology and onset of spring in semi-arid steppe. Great Plains Res., 25: 121-138.
CrossRefDirect Link - Bita, C.E. and T. Gerats, 2013. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Frontiers Plant Sci., Vol. 4.
CrossRefDirect Link - Hatfield, J.L. and J.H. Prueger, 2015. Temperature extremes: Effect on plant growth and development. Weather Clim. Extremes, 10: 4-10.
CrossRefDirect Link - van der Kooi, C.J., P.G. Kevan and M.H. Koski, 2019. The thermal ecology of flowers. Ann. Bot., 124: 343-353.
CrossRefDirect Link - Velásquez, A.C., C.D.M. Castroverde and S.Y. He, 2018. Plant and pathogen warfare under changing climate conditions. Curr. Biol., 28: R619-R634.
CrossRefDirect Link - Bustamante, E. and A. Búrquez, 2008. Effects of plant size and weather on the flowering phenology of the organ pipe cactus (Stenocereus thurberi). Ann. Bot., 102: 1019-1030.
CrossRefDirect Link - Almeida, I.V., M. Monteiro, F.C. Batista, E. Ramalho and R. L. Bruno, 2019. Phenology of Calotropis procera (Ait.) W.T. Aiton accessions based on morphophysiological characteristics. Rev. Caatinga, 32: 543-551.
CrossRefDirect Link - Young, A., P. Gomez-Ruiz, J. Pena, H. Uno and R. Jaffé, 2018. Wind speed affects pollination success in blackberries. Sociobiology, 65: 225-231.
CrossRefDirect Link - Talley, S.M., P.D. Coley and T.A. Kursar, 2002. The effects of weather on fungal abundance and richness among 25 communities in the Intermountain West. BMC Ecol., Vol. 2.
CrossRefDirect Link - Chater, C.C.C., J. Oliver, S. Casson and J.E. Gray, 2014. Putting the brakes on: Abscisic acid as a central environmental regulator of stomatal development. New Phytol., 202: 376-391.
CrossRefDirect Link - Paradiso, R. and S. De Pascale, 2014. Effects of plant size, temperature, and light intensity on flowering of Phalaenopsis hybrids in mediterranean Greenhouses. Sci. World J., Vol. 2014.
CrossRefDirect Link