
Nurul Syazwani Mohd Zahari
Faculty of Bioresources and Food Industry, Universiti Sultan Zainal Abidin, Kampus Besut, 22200 Besut, Terengganu, Malaysia
Hasan Nudin Nur Fatihah
Faculty of Bioresources and Food Industry, Universiti Sultan Zainal Abidin, Kampus Besut, 22200 Besut, Terengganu, Malaysia
LiveDNA: 60.31303
Syaidatul Najiah Zakaria
Faculty of Bioresources and Food Industry, Universiti Sultan Zainal Abidin, Kampus Besut, 22200 Besut, Terengganu, Malaysia
Nadiawati Alias
Faculty of Bioresources and Food Industry, Universiti Sultan Zainal Abidin, Kampus Besut, 22200 Besut, Terengganu, Malaysia
Nor Hasima Mahmod
Faculty of Bioresources and Food Industry, Universiti Sultan Zainal Abidin, Kampus Besut, 22200 Besut, Terengganu, Malaysia
Asian Journal of Plant Sciences
Year: 2020 | Volume: 19 | Issue: 1 | Page No.: 14-20







ABSTRACT
Background and Objective: Aquilaria sp. (family Thymelaeaceae) or commonly known as gaharu, karas or agarwood is one of the threatened crops that have been listed in the Convention on International Trade in Endangered Species (CITES). Overexploitation in search of valuable agarwood or resinous wood has directly led to scarcity of Aquilaria sp. in the wild. The aim of this study was to investigate the genetic diversity among three Aquilaria sp. (A. malaccensis Lam., A. subintegra Ding Hou and A. sinensis (Lour.) Spreng.) using inter simple sequence repeat (ISSR) markers. Materials and Methods: Five annealing temperatures varying from 47-51°C were optimized. Out of 10 ISSR primers screened, 4 primers revealed clear and repeatable bands. Results: The clearest bands were observed at 50°C, which was later used for primer screening and PCR amplifications. The 4 primers (UBC 855, UBC 845, UBC 811 and UBC 809) yielded a total of 23 bands, of which 20 were polymorphic (87%). The UBC 809 and 845 showed 100% polymorphism with a total of 6 and 4 bands, respectively. The UBC 855 showed 6 polymorphic bands (75%) and UBC 811 showed 4 polymorphic bands (80%). Construction of UPGMA dendrogram has grouped all the 15 accessions of 3 Aquilaria sp. into 2 major clusters, where all A. malaccensis accessions were grouped into cluster I and a combination of A. sinensis and A. subintegra accessions were grouped into cluster II. Conclusion: Findings of this study provide basic information for conservation and utilization of the Aquilaria genetic resources.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Nurul Syazwani Mohd Zahari, Hasan Nudin Nur Fatihah, Syaidatul Najiah Zakaria, Nadiawati Alias and Nor Hasima Mahmod, 2020. Genetic Diversity of Threatened Aquilaria sp. in Malaysia Using Inter Simple Sequence Repeat (ISSR) Markers. Asian Journal of Plant Sciences, 19: 14-20.
DOI: 10.3923/ajps.2020.14.20
URL: https://scialert.net/abstract/?doi=ajps.2020.14.20
DOI: 10.3923/ajps.2020.14.20
URL: https://scialert.net/abstract/?doi=ajps.2020.14.20
INTRODUCTION
Aquilaria sp. (family Thymelaeaceae) is a tropical forest tree that is also known as gaharu, karas or agarwood. There are 25 Aquilaria species have been found in the countries including Bangladesh, Bhutan, India, Indonesia, Iran, Myanmar, Philippines, Thailand and Malaysia1. However, only 5 species (A. beccariana Teigh, A. hirta Ridl., A. malaccensis, A. microcarpa Baill. and A. rostrata Ridl.) can be found in the Peninsular Malaysia and three are introduced species (A. crassna Pierre ex Lecomte, A. sinensis and A. subintegra)2. Aquilaria sp. is currently listed in the Convention on International Trade in Endangered Species (CITES) Appendix II and classified as ‘Vulnerable’ to ‘Critically Endangered’ in the International Union for Conservation of Nature (IUCN3) Red List.
Aquilaria sp. has become beneficial and prominent for the production of agarwood or resinous wood4. The highly prized of this agarwood lead to the overexploitation of this species which directly led to scarcity of these trees in the wild5. Malaysia and Indonesia are the primary exporters of agarwood since agarwood received high demand from the local and international markets6. In Malaysia, A. malaccensis is the primary producer of agarwood and the high grade of gaharu can reach7 up to RM 25 000/kg. In order to hinder the scarcity of this threatened species, several countries including Indonesia, India and China have initiated a plantation for Aquilaria sp. in order to preserve and conserve the species as well as to confront the demand of agarwood supply for industry8,9.
According to Hamrick and Godt10, a careful assay of genetic diversity is important to formulate effective conservation strategies for the threatened and endangered species. For Aquilaria sp., the main problem is that the identification of Aquilaria seedling either in the wild or nursery is quite challenging as the morphological features among the species in the genus are almost similar and could not be clearly distinguished among the species7. Hence, selection based on genetic information using molecular markers is extremely important, as it is more dependable and persistent11. The employment of PCR-based approaches for identification of plant genetic resource has been immensely used because of their simplicity and requirement for small quantity of DNA sample12. Previous studies reported that several PCR-based approaches have been piloted for identification of Aquilaria sp. such as those using microsatellite markers13,14, random amplified Polymorphic DNA (RAPD)15 and plastid trnL-trnF intergenic spacer sequences16. However, these studies had very limited coverage of existing Aquilaria populations and the information generated was not sufficient to practically inform any management plans7. Therefore, it is needed to understand the genetic variation within and between species as it can give information and can determine its adaptability for a long-term survival17. In this study, a molecular technique of inter simple sequence repeat (ISSR) has been performed to evaluate the genetic diversity of 3 Aquilaria sp. namely A. malaccensis, A. sinensis and A. subintegra.
MATERIALS AND METHODS
Study area: This study was conducted for 10 months (September, 2018-June, 2019) in the Laboratory of Molecular Biology, Faculty of Bioresources and Food Industry, Universiti Sultan Zainal Abidin, Kampus Besut, Terengganu, Malaysia.
Plant materials and DNA extraction: A total of 15 accessions from three different Aquilaria species namely A. malaccensis (labelled as M), A. subintegra (labelled as S) and A. sinensis (labelled as SI) were used. Seedlings of A. malaccensis were collected from Forest Research Institute Malaysia (FRIM) Forest Reserve, Merchang, Terengganu; A. subintegra were collected from Alor Gajah, Melaka and A. sinensis were collected from Seri Kembangan, Selangor. All the seedlings were subsequently planted at Ladang Pasir Akar germplasm, Jertih, Terengganu. The genomic DNA was extracted from fresh leaves of each accession using DNeasy Plant Mini Kit (Qiagen, Germany) following the manufacturer’s protocol.
ISSR-PCR conditions and gel electrophoresis: The optimization of ISSR-PCR was performed varying in five different annealing temperatures (Ta) ranged from 47-51°C. Ten ISSR primers (Table 1, Eurofins, Genomics) were screened. The primers generating good amplification patterns were further used for PCR amplification of all the plant samples.
| Table 1: List of ISSR primers used in this study | ||
| ISSR primers | Sequence 5'-3' | Melting temperature (°C) |
| UBC 855 | (AC) 8YT | 53.1 |
| UBC 845 | (CT) 8RG | 48.1 |
| UBC 808 | (AG) 9G | 51.2 |
| UBC 811 | (GA) 9A | 49.0 |
| UBC 815 | (CA) 8T | 50.1 |
| UBC 833 | (AT) 8YG | 28.1 |
| UBC 849 | (GT) 8YA | 51.4 |
| UBC 836 | (AG) 8YG | 50.5 |
| UBC 840 | (GA) 8YT | 47.4 |
| UBC 809 | (GA) 9T | 48.8 |
The PCR amplifications were performed using New England Biolab’s (NEB) kit in a final reaction volume 25 μL containing 2.5 μL of 10X buffer, 0.5 μL of 10 mM dNTPs, 1.0 μL of 10 μM each primer, 2.0 μL of approximately 30 ng mL–1 DNA, 0.1 μL of Taq DNA polymerase and 18.9 μL of sterile distilled water. The PCR mix without DNA template was used as a negative control.
The PCR amplification was performed in a Thermal Cycler (Applied Biosystems™) with the following profile: An initial denaturation at 94°C for 5 min followed by 45 cycles of 94°C for 30 sec, 47-51°C for 45 sec, 72°C for 2 min and with a final extension of 72°C for 5 min. The amplified DNA fragments were separated using electrophoresis on 1% (w/v) horizontal agarose gel in 1 X TBE buffer. The electrophoresis was conducted for 1 h at a constant voltage of 90 V and the gel was photographed under an UV transilluminator. The 1 kb DNA ladder (Promega) was included in the gel as a size reference. The ISSR-PCR amplifications were repeated at least twice to ensure the band profiles were stable and reproducible.
Data analysis: Only clear and distinct ISSR bands were scored as presence (1) or absence (0). A binary ISSR data matrix was prepared for each species and used in the subsequent analysis.
Basic parameters such as the total number of bands, number of polymorphic bands and the percentage of polymorphic bands were manually calculated according to the equation18:
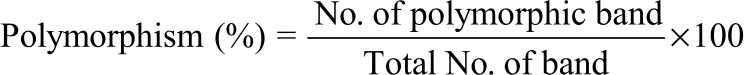 |
A dendrogram was constructed by unweighted pair group method using arithmetic averages (UPGMA)19 and the genetic distances were calculated based on Nei20 unbiased genetic distance. The analysis was performed by PAUP* software (version 4.0)21.
RESULTS AND DISCUSSION
Annealing temperature and primer screening: In order to obtain an efficient identification, conservation and utilization of the threatened species of Aquilaria, extensive research on PCR-based markers is needed. Among all dominant markers, ISSR have its advantages with higher polymorphism of bands, lower cost and have been widely used for the genetic diversity studies of various crop plants22-24. The adjustment of annealing temperature is known to have a great impact on the richness and legibility of DNA banding profile.
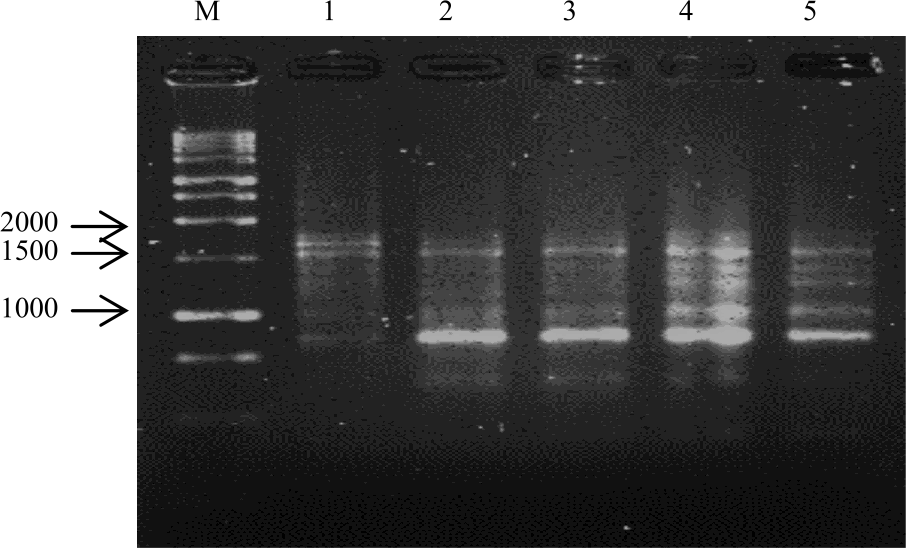 |
| Fig. 1: | Effects of annealing temperature on ISSR-PCR amplification (UBC 808 using sample A. subintegra) Lane M: 1 kb marker, Lane 1: 47°C, Lane 2: 48°C, Lane 3: 49°C, Lane 4: 50°C, Lane 5: 51°C |
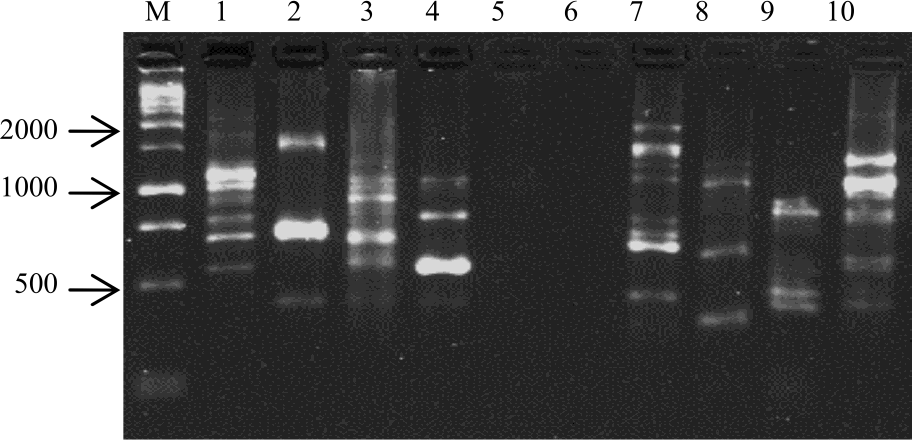 |
| Fig. 2: | ISSR-PCR band profiles generated by ten primers using sample A. sinensis Lane M: 1 kb marker, Lane 1: UBC 855, Lane 2: UBC 845, Lane 3: UBC 808, Lane 4: UBC 811, Lane 5: UBC 815, Lane 6: UBC 833, Lane 7: UBC 849, Lane 8: UBC 836, Lane 9: UBC 840, Lane 10: UBC 809 |
The higher temperature is favoured for large fragments amplification whilst the lower temperature is favoured for short fragments amplification25. According to Bornet and Branchard12, annealing temperature (Ta) is primer specific and always superior to melting temperature (Tm) because of the need of high stringency to facilitate annealing of primers. They reported that Ta values of 45-50°C were optimal for ISSR amplification with different primers. In the present study, the optimization of Ta varying from 47-51°C was achieved by performing a gradient PCR. Using the primer UBC 808 and DNA sample of A. subintegra, the clearest and intensive bands were observed at an optimal annealing temperature of 50°C (Fig. 1), which was later used for primer screening. Out of 10 primers screened, 4 primers (UBC 855, UBC 845, UBC 811 and UBC 809) produced the clearest, intensive and reproducible bands, whilst the primers UBC 815 and UBC 833 gave no amplified bands on the tested sample of A. sinensis (Fig. 2).
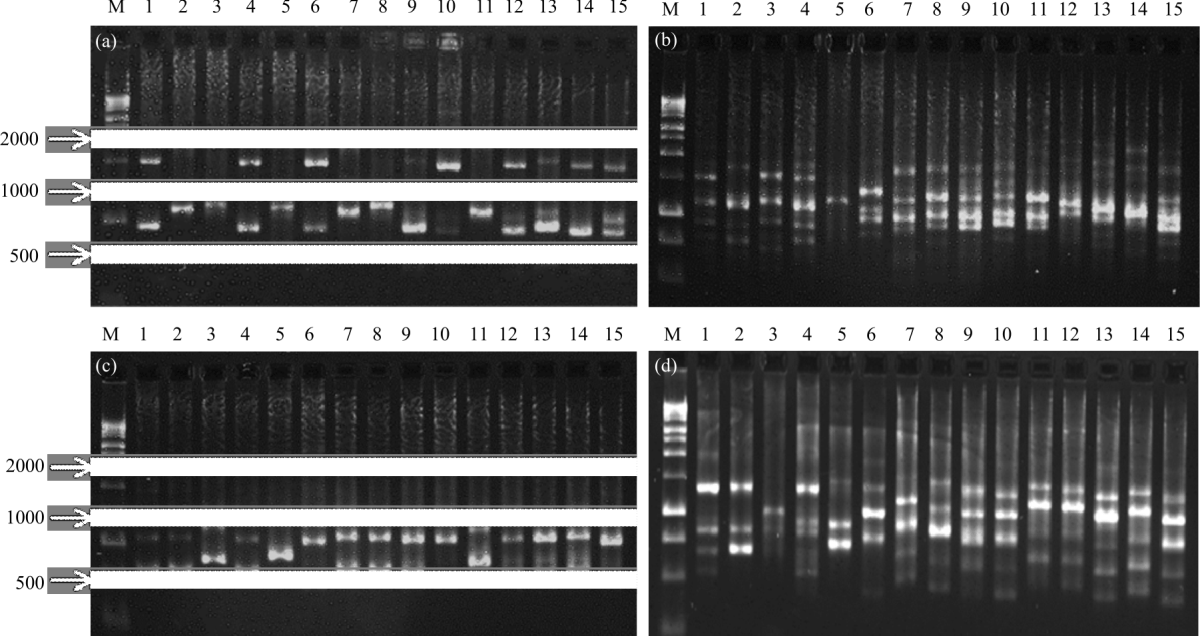 |
| Fig. 3(a-d): | PCR amplifications of 4 selected primers, (a) UBC 855, (b) UBC 845, (c) UBC 811 and (d) UBC 809 Lane M: 1 kb marker, Lane 1: A. malaccensis accession M1, Lane 2: A. malaccensis accession M3, Lane 3: A. malaccensis accession M5, Lane 4: A. malaccensis accession M7, Lane 5: A. malaccensis accession M8, Lane 6: A. subintegra accession S2, Lane 7: A. subintegra accession S4, Lane 8: A. subintegra accession S5, Lane 9: A. subintegra accession S7, Lane 10: A. subintegra accession S9, lane 11: A. sinensis accession SI 1, Lane 12: A. sinensis accession SI 3, Lane 13: A. sinensis accession SI 5, Lane 14: A. sinensis accession SI 6, Lane 15: A. sinensis accession SI 8 |
| Table 2: Percentage of polymorphic band obtained from selected primers | ||||
| Primers | No. of polymorphic band | No. of monomorphic band | Total band produced | Polymorphism (%) |
| UBC 855 | 6 | 2 | 8 | 75 |
| UBC 845 | 4 | 0 | 4 | 100 |
| UBC 811 | 4 | 1 | 5 | 80 |
| UBC 809 | 6 | 0 | 6 | 100 |
ISSR polymorphism and genetic diversity: The 4 selected primers (UBC 855, UBC 845, UBC 811 and UBC 809) were subsequently used for genetic diversity analysis of three Aquilaria sp. A total of 23 bands were amplified, of which 20 bands were polymorphic (87%) and the remaining bands were monomorphic (13%). The percentage of polymorphism for each primer varied from 75-100% (Table 2). The ISSR-PCR amplification by UBC 855 resulted in 8 polymorphic bands ranging from 625-2000 bp, UBC 845 produced 4 bands ranging from 750-1750 bp, UBC 809 produced 6 bands ranging from 500-1500 bp, whilst amplification by UBC 811 resulted in 5 bands ranging from 500-2000 bp (Fig. 3). The primer UBC 855 produced the highest number of bands but the lowest percentage of polymorphism (75%). On the other hand, both primers UBC 845 and 809 showed 100% polymorphism and similar result is also reported by Banu et al.26. Variable banding patterns were observed in three Aquilaria sp., however, no species-specific band was produced by any primer at any band range.
Table 3 showed the genetic distances based on the similarity coefficients of Nei20 between all the Aquilaria accessions. The UPGMA dendrogram showed 2 major clusters, I and II (Fig. 4). Cluster I consisted of all A. malaccensis accessions. Cluster II consisted of combination of A. subintegra and A. sinensis accessions. The major cluster I formed 2 sub clusters, IA and IB. The sub cluster IA was then divided into 2 sub clusters, IA1 and IA2. Two accessions of A. malaccensis (M1 and M7) were clustered in IA1 while another two accessions of A. malaccensis (M3 and M5) were clustered in IA2. The sub cluster IB consisted of one accession of A. malaccensis (M8). Major cluster II composed of two sub clusters, IIA and IIB. The sub cluster IIA was further divided into 2 sub clusters, IIA1 and IIA2. Three A. subintegra accessions (S2, S9 and S7) were clustered in the sub cluster IIA1 while two A. subintegra accessions (S4 and S5) clustered in the sub cluster IIA2. The sub cluster IIB consisted of all A. sinensis. An accession of A. sinensis (SI 1) was clustered in sub cluster IIB1 followed by the four accessions of A. sinensis (SI 3, SI 6, SI 8 and SI 5) which were clustered in IIB2.
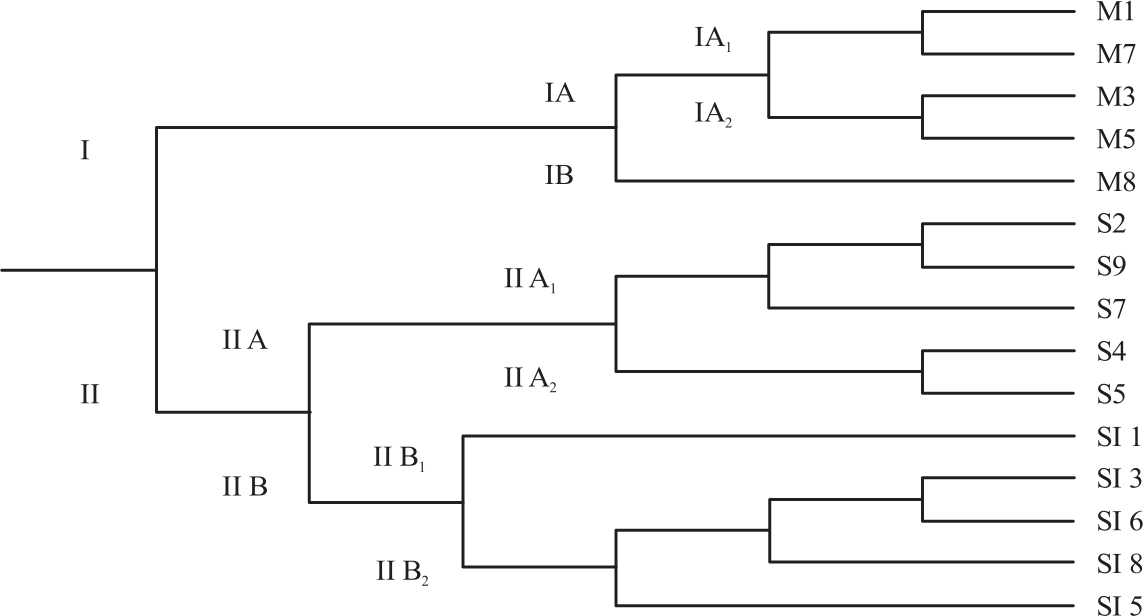 |
| Fig. 4: | Dendrogram of 15 accessions of A. malaccensis (labelled as M), A. subintegra (labelled as S) and A. sinensis (labelled as SI) based on ISSR data |
| Table 3: Genetic distance matrix of three Aquilaria sp. based on 23 bands generated by 4 primers used, based on Nei20 coefficients | |||||||||||||||
| Strain | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 |
| 1 | - | ||||||||||||||
| 2 | 0.2174 | - | |||||||||||||
| 3 | 0.3913 | 0.1739 | - | ||||||||||||
| 4 | 0.0000 | 0.2174 | 0.3913 | - | |||||||||||
| 5 | 0.4783 | 0.3478 | 0.3478 | 0.4783 | - | ||||||||||
| 6 | 0.3478 | 0.5652 | 0.5652 | 0.3478 | 0.5652 | - | |||||||||
| 7 | 0.4783 | 0.3478 | 0.4348 | 0.4783 | 0.5217 | 0.2173 | - | ||||||||
| 8 | 0.4348 | 0.3044 | 0.3913 | 0.4348 | 0.4783 | 0.2609 | 0.0434 | - | |||||||
| 9 | 0.2609 | 0.4783 | 0.5652 | 0.2609 | 0.5652 | 0.0867 | 0.2174 | 0.1734 | - | ||||||
| 10 | 0.3478 | 0.5652 | 0.5652 | 0.3478 | 0.5652 | 0.0000 | 0.2174 | 0.2609 | 0.0870 | - | |||||
| 11 | 0.5652 | 0.4348 | 0.4348 | 0.5652 | 0.5217 | 0.4783 | 0.2609 | 0.2174 | 0.3913 | 0.4783 | - | ||||
| 12 | 0.4348 | 0.5652 | 0.5652 | 0.4348 | 0.6522 | 0.2609 | 0.3044 | 0.3478 | 0.2609 | 0.2609 | 0.2174 | - | |||
| 13 | 0.3913 | 0.5217 | 0.5217 | 0.3913 | 0.6087 | 0.3044 | 0.3478 | 0.3044 | 0.2174 | 0.3044 | 0.1739 | 0.0435 | - | ||
| 14 | 0.4348 | 0.5652 | 0.5652 | 0.4348 | 0.6522 | 0.2609 | 0.3043 | 0.3478 | 0.2609 | 0.2069 | 0.2174 | 0.0000 | 0.0435 | - | |
| 15 | 0.4348 | 0.5652 | 0.5652 | 0.4348 | 0.6522 | 0.2609 | 0.3043 | 0.3478 | 0.2609 | 0.2069 | 0.2174 | 0.0000 | 0.0435 | 0.000 | - |
| 1: A. malaccensis accession M1, 2: A. malaccensis accession M3, 3: A. malaccensis accession M5, 4: A. malaccensis accession M7, 5: A. malaccensis accession M8, 6: A. subintegra accession S2, 7: A. subintegra accession S4, 8: A. subintegra accession S5, 9: A. subintegra accession S7, 10: A. subintegra accession S9, 11: A. sinensis accession SI 1, 12: A. sinensis accession SI 3, 13: A. sinensis accession SI 5, 14: A. sinensis accession SI 6, 15: A. sinensis accession SI 8 | |||||||||||||||
Aquilaria malaccensis was genetically distant from A. subintegra and A. sinensis. This study was in agreement with Lee et al.17 where all A. malaccensis were clustered into the same cluster, in which it can strictly distinguishable from A. hirta and A. rostrata collected from the Peninsular Malaysia. The larger genetic distance observed between the A. malaccensis with A. subintegra and A. sinensis might be explained by the species geographic distributions where both A. subintegra and A. sinensis were the introduced species to Malaysia, while A. malaccensis was native to Malaysia. Geographical variability is one of the prevalent drivers that can develop high level of polymorphisms between populations, as environment differences such as the climate, moisture content, soil nutrient, forest diversity and others can be different for both East and Peninsular Malaysia forests7. Such phenomena was supported by Josiah et al.27, who stated that geographical isolation may reduce gene flow, causing pollen and seed dispersal to be limited within the population and hence, reduces the gene variability of the population.
Assessment of genetic diversity and relationships within and between crop species and their wild relatives is vital not only for basic studies of evolution but also for informed utilization and protection of plant genetic resources28-31. In addition, a thorough knowledge on the genetic diversity of a crop is crucial for selection of parental material that maximizes genetic variability32. Among various PCR-based markers, ISSR markers have been recommended for assessment of genetic diversity because the ISSR markers were highly polymorphic and were successfully used to elucidate the patterns of genetic variation in plants at population, species as well as genus level17,26,33.
CONCLUSION
This study indicates that ISSR markers could provide reproducible and specific tools to study genetic diversity among three Aquilaria sp. namely A. malaccensis, A. sinensis and A. subintegra. Four ISSR makers (UBC 855, UBC 845, UBC 811 and UBC 809) are found to be suitable for polymorphism assessment in A. malaccensis, the main producer of agarwood in Malaysia. Although no species-specific band was produced, ISSR markers are successfully used to elucidate the levels and patterns of genetic variation among Aquilaria species. Such information is important to guide conservation and management efforts of this threatened species. The presence of species-specific band will be of great value for the development of species-specific marker. Further studies using more markers such as RAPD and RFLP are required.
SIGNIFICANCE STATEMENT
This study discovered the potential use of ISSR markers to investigate the genetic diversity among three threatened Aquilaria sp. namely A. malaccensis, A. sinensis and A. subintegra. The findings of this study can be beneficial for the formulation of conservation strategies and utilization of Aquilaria genetic resources. This study will help the researchers to uncover the critical areas of genetic variation among Aquilaria sp. that many researchers were not able to explore. Thus a new theory using ISSR markers describing the genetic variation and polymorphism in A. malaccensis, the main producer of agarwood in Malaysia is now available.
ACKNOWLEDGMENT
This study was funded by Special Research Grant Scheme (SRGS) from UniSZA (UniSZA/2017/SRGS/19).
REFERENCES
- Faridah-Hanum, I., M.Z. Mustapa, P. Lepun, T.I.T. Marina, M. Nazri, R. Alan and R. Mohamed, 2009. Notes on the distribution and ecology of Aquilaria Lam. (Thymelaeaceae) in Malaysia. Malaysian Forester, 72: 247-259.
Direct Link - Nor Azah, M.A., Y.S. Chang, J. Mailina, A. Abu Said, S.A.M.J.S. Husni, H. Nor Hasnida and Y.N. Yasmin, 2008. Comparison of chemical profiles of selected gaharu oils from Peninsular Malaysia. Malays. J. Anal. Sci., 12: 338-340.
Direct Link - Azhari, H., A. Mohamad and R. Othman, 2015. Molecular identification of Aquilaria spp. by using Inter-Simple Sequence Repeat (ISSR). AIP Conf. Proc., Vol. 1678, No. 1.
CrossRefDirect Link - Naef, R., 2011. The volatile and semi-volatile constituents of agarwood, the infected heartwood of Aquilaria species: A review. Flavour Fragr. J., 26: 73-87.
CrossRefDirect Link - Soehartono, T. and A.C. Newton, 2001. Reproductive ecology of Aquilaria spp. in Indonesia. For. Ecol. Manage., 152: 59-71.
CrossRefDirect Link - Mohamed, R., P.L. Jong and A.K. Kamziah, 2014. Fungal inoculation induces agarwood in young Aquilaria malaccensis trees in the nursery. J. For. Res., 25: 201-204.
Direct Link - Eurlings, M.C.M., H.H. van Beek and B. Gravendeel, 2010. Polymorphic microsatellites for forensic identification of agarwood (Aquilaria crassna). Forensic Sci. Int., 197: 30-34.
CrossRefPubMedDirect Link - Zhang, X.L., Y.Y. Liu, J.H. Wei, Y. Yang and Z. Zhang et al., 2012. Production of high-quality agarwood in Aquilaria sinensis trees via whole-tree agarwood-induction technology. Chin. Chem. Lett., 23: 727-730.
CrossRefDirect Link - Hamrick, J.L. and M.J.W. Godt, 1996. Effects of life history traits on genetic diversity in plant species. Philos. Trans. R. Soc. London B: Biol. Sci., 351: 1291-1298.
CrossRefDirect Link - Bornet, B. and M. Branchard, 2001. Nonanchored Inter Simple Sequence Repeat (ISSR) markers: Reproducible and specific tools for genome fingerprinting. Plant Mol. Biol. Rep., 19: 209-215.
Direct Link - Tnah, L.H., C.T. Lee, S.L. Lee, K.K.S. Ng and C.H. Ng et al., 2012. Isolation and characterization of microsatellite markers for an important tropical tree, Aquilaria malaccensis (Thymelaeaceae). Am. J. Bot., 99: e431-e433.
CrossRefDirect Link - Zhang, Y.T., Z.F. Wang, H.L. Cao, X.Y. Li, L.F. Wu, S.B. Zhuo and X.F. Huang, 2010. Isolation and characterization of polymorphic microsatellite loci in Aquilaria sinensis (Lour.) Gilg. Conserv. Genet. Resour., 2: 5-6.
Direct Link - Lee, S.Y., J. Weber and R. Mohamed, 2011. Genetic variation and molecular authentication of selected Aquilaria species from natural populations in Malaysia using RAPD and SCAR markers. Asian J. Plant Sci., 10: 202-211.
CrossRefDirect Link - Eurlings, M.C.M. and B. Gravendeel, 2005. TrnL-trnF sequence data imply paraphyly of Aquilaria and Gyrinops (Thymelaeaceae) and provide new perspectives for agarwood identification. Plant Syst. Evol., 254: 1-12.
CrossRefDirect Link - Lee, S.Y., W.L. Ng, D.U. Lamasudin and R. Mohamed, 2018. Inter-simple sequence repeat markers reveal genetic relatedness between natural aquilaria populations in Peninsular Malaysia. Chiang Mai J. Sci., 45: 1307-1317.
Direct Link - Tidke, S. D. and P.S. Ranawade, 2017. Genetic analysis and RAPD polymorphism in wheat (Triticum aestivum L.) genotypes. Int. J. Curr. Microbiol. Applied Sci., 6: 239-246.
CrossRefDirect Link - Dje, Y., G.G. Tahi, I.A. Zoro Bi, M. Malice, J.P. Baudoin and P. Bertin, 2006. Optimization of ISSR marker for African edible-seeded Cucurbitaceae species' genetic diversity analysis. Afr. J. Biotechnol., 5: 83-87.
Direct Link - Chin, G.J.W.L., M.Z. Mohamad, S. Maili, W.T.L. Yong and K.F. Rodrigues, 2017. ISSR-PCR fingerprinting of Kappaphycus and Eucheuma (Rhodophyta, Gigartinales) seaweed varieties from Sabah, Malaysia. Trans. Sci. Technol., 4: 420-425.
Direct Link - Shaw, R.K., L.K. Acharya and A.K. Mukherjee, 2009. Assessment of genetic diversity in a highly valuable medicinal plant Catharanthus roseus using molecular markers. Crop Breed. Applied Biotechnol., 9: 52-59.
Direct Link - Moraga-Suazo, P., R. Hasbun, C. Balocchi and S. Valenzuela, 2012. Establishment and optimization of ISSR and SAMPL molecular markers as a tool for breeding programs of Pinus radiata. Bosque, 33: 93-98.
CrossRefDirect Link - Banu, S., D. Baruah, R.M. Bhagwat, P. Sarkar, A. Bhowmick and N.Y. Kadoo, 2015. Analysis of genetic variability in Aquilaria malaccensis from Bramhaputra valley, Assam, India using ISSR markers. Flora-Morphol. Distrib. Funct. Ecol. Plants, 217: 24-32.
CrossRefDirect Link - Josiah, C.C., D.O. George, O.M. Eleazar and W.F. Nyamu, 2008. Genetic diversity in Kenyan populations of Acacia senegal (L.) willd revealed by combined RAPD and ISSR markers. Afr. J. Biotechnol., 7: 2333-2340.
Direct Link - Rao, V.R. and T. Hodgkin, 2002. Genetic diversity and conservation and utilization of plant genetic resources. Plant Cell Tissue Org. Cult., 68: 1-19.
Direct Link - Govindaraj, M., M. Vetriventhan and M. Srinivasan, 2015. Importance of genetic diversity assessment in crop plants and its recent advances: An overview of its analytical perspectives. Genet. Res. Int., Vol. 2015.
CrossRefDirect Link - Brozynska, M., A. Furtado and R.J. Henry, 2016. Genomics of crop wild relatives: Expanding the gene pool for crop improvement. Plant Biotechnol. J., 14: 1070-1085.
CrossRefDirect Link - Kiani, G. and M. Siahchehreh, 2017. Diversity in squash varieties assessed by ISSR markers. Int. J. Veg. Sci., 23: 430-437.
CrossRefDirect Link - Javan, Z.S., F. Rahmani and R. Heidari, 2012. Assessment of genetic variation of genus Salvia by RAPD and ISSR markers. Aust. J. Crop Sci., 6: 1068-1073.
Direct Link