
Hamim Hamim
Department of Biology, Faculty of Mathematics and Natural Sciences, Bogor Agricultural University, Jl. Agathis, Kampus IPB Darmaga, 16680 Bogor, Indonesia
LiveDNA: 62.15393
Anisah Mutyandini
Department of Biology, Faculty of Mathematics and Natural Sciences, Bogor Agricultural University, Jl. Agathis, Kampus IPB Darmaga, 16680 Bogor, Indonesia
Yohana C. Sulistyaningsih
Department of Biology, Faculty of Mathematics and Natural Sciences, Bogor Agricultural University, Jl. Agathis, Kampus IPB Darmaga, 16680 Bogor, Indonesia
Hirmas Fuady Putra
Department of Biology, Faculty of Mathematics and Natural Sciences, Bogor Agricultural University, Jl. Agathis, Kampus IPB Darmaga, 16680 Bogor, Indonesia
Deden Saprudin
Department of Chemistry, Faculty of Mathematics and Natural Sciences, Bogor Agricultural University, Bogor, Indonesia
Luluk Setyaningsih
Faculty of Forestry, Nusa Bangsa University, Bogor, Indonesia
Asian Journal of Plant Sciences
Year: 2019 | Volume: 18 | Issue: 4 | Page No.: 164-174







ABSTRACT
Background and Objective: Plants have different response to heavy metals and some plants have ability to absorb and reduce heavy metal contaminant such as lead and mercury from the environment. The objective of this experiment was to analyze the morphological, anatomical and physiological responses of four non-edible oil producing plants to mercuric (Hg) treatment in hydroponic culture. Materials and Methods: About 3 months old of 4 species (Melia azedarach, Ricinus communis, Jatropha curcas and Reutealis trisperma) were grown in water culture using Hoagland solution treated with mercuric nitrate [Hg(NO3)2] of 0, 0.5, 1, 2 and 3 mM for 21 days. The observation was made to see morphological, anatomical and physiological responses of the plants to Hg treatments. Results: Mercury (Hg) decreased dramatically the growth rate of the plants including plant height, number of leaves and the dry weight of shoot as well as roots, even though there was variation among the species. Mercuric treatment also induced lipid peroxidation which was marked by the increase of malondialdehyde content (MDA), while it caused dramatic decrease of total chlorophyll content. The increase of Hg accumulation on plant tissue caused the alteration of anatomy structure like decreasing the thickness of upper layer of epidermis, palisade and spongy mesophyll cell, which at the end it caused the decrease of leaf thickness. Conclusion: Based on morphological, anatomical and physiological changes, R. trisperma was the most tolerant to mercuric treatments, followed by M. azedarach, while R. communis and J. curcas were categorized as the less tolerant to Hg treatments compared to the other two.
PDF Abstract XML References Citation
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Hamim Hamim, Anisah Mutyandini, Yohana C. Sulistyaningsih, Hirmas Fuady Putra, Deden Saprudin and Luluk Setyaningsih, 2019. Effect of Mercury on Growth, Anatomy and Physiology of Four Non-edible Oil-producing Species. Asian Journal of Plant Sciences, 18: 164-174.
DOI: 10.3923/ajps.2019.164.174
URL: https://scialert.net/abstract/?doi=ajps.2019.164.174
DOI: 10.3923/ajps.2019.164.174
URL: https://scialert.net/abstract/?doi=ajps.2019.164.174
INTRODUCTION
Mercury (Hg) is one of heavy metal pollutant that has received attention by global community since this substance has been extensively used in various fields especially in industrial activities which causes various negative impacts on the environment. There are many evidences that artisanal gold mining contributes to the largest mercuric contaminant to the earth, which resulted in wide range of environmental devastation and threatening human health1. Mercury is classified as a heavy metal that has the most dangerous toxic effects to the plant, animal and human2. Toxic effects of Hg can adversely affect plant morphology, physiology and biochemistry. Hg accumulation in plants causes an increase in the formation of reactive oxygen species (ROS) which can react with macromolecules such as DNA, pigments, proteins, lipids and other cellular molecules that cause a series of damage processes for plants called oxidative stress3. This heavy metal has also been reported to cause leakage of plasma membranes, changes in antioxidant enzyme activity in plants and induce the expression of genes that encode superoxide dismutase, peroxidase and catalase4. Therefore, serious efforts should be done systematically in order to avoid environmental damage due to these pollutants.
The efforts to clean heavy metals from polluted land can be carried out by specific (physical or chemical) treatments but it requires a very expensive cost. One alternative that can be used to overcome the problems due to heavy metal pollution with lower cost is using a method known as phytoremediation5, which use the plants to reduce or eliminate the metal content from the soil6. Many plant species including weeds, shrubs as well as trees have been suggested to have ability to reduce metal contaminants7-10. These plants may have particular physiological characteristics which able to avoid toxic effect of metal component inside the cells and tissues.
Plants have various defense mechanisms including detoxifying heavy metals. Heavy metal detoxification in higher plants involves the process of metal curing in the cytosol by high affinity ligands, including amino acids and organic acids as well as two classes of polypeptides namely phytochelatins (PCs) and metallothioneins (MTs) at the intra and intercellular levels11. Non-enzymatic synthesized compounds such as proline (Pro) are also known to be able to increase the detoxification capacity of metals from intracellular antioxidant enzymes. Other important additional components of plant defense systems are symbiotic associations with bacteria or mycorrhizal fungi. Microbial organisms including bacteria and arbuscular mycorrhiza may be able not only to improve plant growth but, in many cases also be able to effectively detoxify heavy metals, increase plant antioxidant defense activity and reduce metal absorption by host plants12-14. Nonetheless, the ability of plants themselves internally to adapt to metal toxicity is necessary and become an important marker for tolerant plants before the plants are associated with certain microbes. Therefore, the selection of plant species adaptive to heavy metals is one of important factors that determine the success of phytoremediation program.
There are many specific criteria of the plant that can be used for phytoremediation program, including rapid growth, higher biomass, heavy and dept root growth, tolerant to heavy metals and able to accumulate large amounts of metal and tolerant to drought, soil pH and salinity15. In addition, utilization of plants which are able to produce non-edible oil in phytoremediation program would have double benefits because the plants have ability to reduce heavy metal while produce biodiesel feedstock16. Some species such as melia (Melia azedarach L.), castor bean (Ricinus communis L.), jatropha (Jatropha curcas) and wild-candlenut (Reutealis trisperma) are a group of non-edible oil producing plants that able to produce biodiesel stock and have been proved to be able to grow in marginal lands including in post mining areas17. Therefore, these species may be potential to be utilized as phytoremediator plants and necessary for further investigation.
This study aimed to observe the morphological, anatomical and physiological characteristics of four non-edible oil-producing plant species in response to mercuric toxicity applied using hydroponic system.
MATERIALS AND METHODS
Time and location: The experiment was carried out from April, 2018 until August, 2018, in the Glass House and the Laboratory of Physiology and Molecular Biology, Department of Biology, Faculty of Mathematics and Natural Sciences, Bogor Agricultural University (IPB University), Bogor, Indonesia.
Experimental design and Hg treatment: The experiment was carried out using completely randomized design with 2 factors. The first factor was 4 plant species namely jatropha (Jatropha curcas L.), castor bean (Ricinus communis L.), melia plant (Melia azedarach L.) and wild candle nut (Reutealis trisperma [Blanco]). The second factor was Hoagland-based water culture contained 5 different concentrations of mercuric nitrate [Hg(NO3)2] which were 0 (control), 0.5, 1, 2 and 3 mM. Three-month-old plants were transferred into water culture tank with a capacity of 8 L for 3 weeks to support plant establishment before mercuric treatments. The plants were grown in the container (20×25×20 cm3) with Hoagland solution based on the composition of Epstein and Bloom18. In each container, stereo-foam was mounted to support plant standing and equipped with aerator for air supply. Four plants were installed in each container using sponge support. After 3 weeks establishment, the water culture media were treated with mercuric nitrate [Hg(NO3)2] with various concentrations as mentioned above. All the plants were treated with different mercuric concentration for 3 weeks to observe different responses among the species. The addition of water was carried out to the container every two days until it reached its initial volume to avoid concentrated effect due to transpiration.
Morphological parameters measurement: Plant growth was observed by measuring plant height, leaf number and dry weight of shoot and roots. Plant height was measured from the base of the stem to the growing point of the plant. Shoot and roots dry weight was measured after the samples were dried using oven at 80°C for 3 days. Morphological changes were observed during the experiment between initial and the last measurement of the leaves and the plant. Shoot and root dry weights were measured at the last period of the treatments.
Analysis of leaf anatomy: Observation of leaf anatomy was carried out by creating transverse sections using a razor blade (free hand section method). Anatomic parameters observed were including upper epidermal thickness, palisade tissue, spongy tissue, lower epidermis and leaf thickness. For this experiment, only control plants (without Hg treatment) and the plants treated with 3 mM of Hg that were observed to analyze their response. Each leaf incision was observed from 5 different fields of view. Observations were made using Olympus CX-21 microscope with 40×10 magnifications. Photo preparation was taken using Optilab Viewer® v.2.1. The thickness measurements of each anatomic parameter observed were carried out with ImageJ software (NIH, USA).
Lipid peroxidation (malondialdehyde) analysis: Quantitative analysis of lipid peroxidation was performed by measuring the levels of malondialdehyde (MDA), the end result of lipid peroxidation, by following the method of Hodges et al.19. A half gram of leaves was crushed with the addition of 5 mL of trichloroacetic acid (TCA) 5%. The solution obtained was centrifuged at 2500 rpm, 20 min, 4°C. A total of 2 mL of homogenic centrifuge solution was added to a 3 mL of 0.5% thiobarbituric acid (TBA) in 5% of TCA, incubated in an 80°C water bath for 30 min, then soaked in water until cool. The solution was centrifuged back at 2500 rpm, 20 min, 4°C. The supernatant obtained was measured using spectrophotometer at wavelengths of 450, 532 and 600 nm with a blank solution of 5 mL of TCA 5% and 3 mL of 0.5% TBA in 5% of TCA. MDA levels of leave and roots/g fresh weight (FW) were determined by spectrophotometer at a wavelength of 450, 532, 600 nm and calculated using the formula:
[MDA] (μmol g–1 FW) = 6.45×(D532-D600)-0.56×D450
Chlorophyll content analysis: Total chlorophyll analysis of leaves was carried out to see the senescence level of plant leaves. Photosynthetic pigment analysis was carried out based on Quinet et al.20. Fresh leaves were cut into small pieces and weighed as much as 0.1 g. The leaves were then crushed using mortar with sufficient addition of acetone 80% to reach 10 mL in a test tube. The absorbance was measured in a spectrophotometer (Gysys 20 Thermo Spectronic) at wavelengths (λ) of 646 and 663 nm. The content of leaf chlorophyll was calculated by Lichtenthaler21 equation for 80% acetone:
Total chlorophyll (μg g–1 FW) = 7.15A663+18.71A646
Statistical analysis: The data were analyzed using two-ways ANOVA at 95% confidence level using SPSS version 16.0 software. Further testing between treatments and their interactions was carried out using Duncan's Multiple Range Test (DMRT) at α = 0.05. The average data of anatomical, morphological and physiological characters were then used22 for principle component analysis (PCA) MetaboAnalyst 4.0. Data standardization was carried out through Microsoft Excel 2016 program before the data was used in the analysis of PCA23.
RESULTS AND DISCUSSION
RESULTS
Increase of plant height: The treatment with mercury caused dramatic inhibition to growth of all the species. Based on the increase of plant height, among all the species, J. curcas had the highest growth rate followed by R. communis, while M. azedarach had the lowest.
| Table 1: | Increase of plant height (%) of 4 species in response to different concentration of Hg for 21 days |
 | |
Values followed by similar alphabets are not significantly different based on DMRT test at 5% of α | |
| Table 2: | Increase of leaves number of 4 species in response to Hg exposure for 21 days |
 | |
Values followed by similar alphabets are not significantly different based on DMRT test at 5% of α | |
The measurement of plant height showed that the increase of plant height was almost stagnant during 21 days of the Hg exposure even at the lowest concentration of Hg (0.5 mM), except for R. communis and J. curcas that still grew (Table 1). These 2 species categorized as shrubs which under control treatment (without Hg) had higher growth rate than the other two which were tree species (M. azedarach and R. trisperma). Mercuric treatments from 1 mM of Hg caused all the species exactly ceased and did not grow any more (Table 1). The concentration of 0.5 mM of Hg treatment was critical for the species observed in this experiment since only half of the species was still able to grow.
Increase of leaves number: The Hg exposure reduced leaves development especially for J. curcas, R. communis and M. azedarach which was associated with Hg concentration (Table 2), while in R. trisperma the reduction of leaves number was only relatively small and not significantly different (p>0.05). The Table 2 also showed that the dramatic reduction of leaves number was happened even started at the concentration of Hg 0.5 mM, although until 3 mM of Hg, the development of leaves in R. trisperma and J. curcas were still happen.
Plant biomass: The negative effect of mercury was detected significantly (p<0.05) by the decrease of plant biomass due to mercuric treatments for 21 days. Hg treatment caused dramatic decrease in roots dry weight of all the species except R. trisperma which did not undergo reduction in roots dry weight until the plant was treated with 3 mM of Hg (Fig. 1). Similar pattern was provided by shoot dry weight which also decreased dramatically in response to Hg treatments, even though M. azedarach and R. trisperma underwent less reduction (Fig. 2).
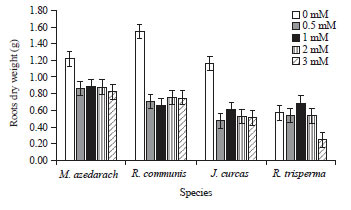 | |
| Fig. 1: | Roots dry weight of 4 species treated with different Hg concentration of 0, 0.5, 1, 2 and 3 mM for 21 days |
Mean+SE of DMRT test at α of 5%, n = 3 | |
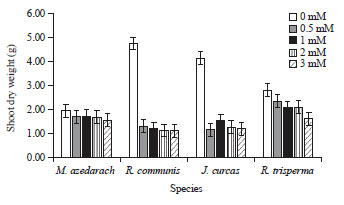 | |
| Fig. 2: | Shoot dry weight of 4 species treated with different Hg concentration of 0, 0.5, 1, 2 and 3 mM for 21 days |
Mean+SE of DMRT test at α of 5%, n = 3 | |
R. communis and J. curcas were the most affected by Hg treatment with the reduction of roots as well as shoot dry weight more than 50% compared to the control without Hg treatment (Fig. 1 and 2).
There was different response of roots and shoot of M. azedarach which indicated that its roots dry weight was more affected by Hg exposure than the shoot.
| Table 3: | Leaves anatomical structure of 4 species in response to Hg treatments at 0 and 3 mM |
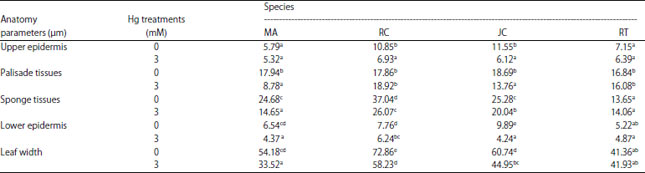 | |
Values followed by similar alphabets are not significantly different based on DMRT test at 5% of α, MA: M. azedarach, RC: R. communis, JC: J. curcas, RT: R. trisperma | |
On the other hand, the other 3 species showed that shoot was more susceptible to Hg exposure than roots (Fig. 1 and 2). Among all Hg concentration that were applied, the effect in biomass reduction was almost similar starting from 0.5 mM until 3 mM, except in R trisperma which showed different effect and for this species the dramatic biomass reduction was happened when Hg was applied at 3 mM (Fig. 1 and 2).
Leaf anatomy analysis: Mercury treatment also affected leaf anatomy of 4 species even though there was variation among the species. The results showed that Hg treatment caused significant changes in leaves thickness due to reduction of some tissues width except in R. trisperma (Table 3). The highest decrease in leaf thickness was found in M. azedarach (38%), followed by J. curcas (26%) and R. communis (17 %). R. trisperma did not undergo any changes in leaf width but even experienced slightly thickening on spongy tissues which caused slight increase of leaves width (Table 3). This showed that R. trisperma was the least affected by Hg treatments based on the changes in leaf anatomical structure compared to three other plants.
Lipid peroxidation analysis: Malondialdehyde is a product of lipid peroxidation process that occurs when the plants experience stress. Increased concentration of Hg in the media caused an increase in leaf MDA concentration dramatically in all 4 plant species. The MDA content in M. azedarach and R. communis increased dramatically in response to mercuric treatment, while the lowest increased of MDA was found in R. trisperma (Fig. 3). For the control plants, R. communis had the lowest MDA content than the other three, therefore at the maximum Hg treatment (3 mM of Hg), MDA level in this species increased more than 8 times than that of control, while in M. azedarach, the increase of MDA level due to maximum Hg treatment was only approximately 3.5 times than control level (Fig. 3).
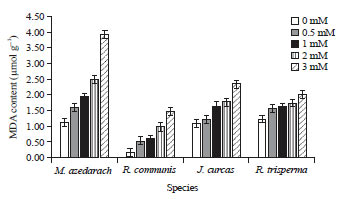 | |
| Fig. 3: | Leaves MDA content of 4 plant species subjected to Hg treatment at the concentration of 0, 0.5, 1, 2 and 3 mM Mean+SE of DMRT test at α of 5%, n = 3 |
The MDA level in J. curcas and R. trisperma was only increase 2.2 and 1.6 times higher respectively than control plants.
Chlorophyll content analysis: Mercuric treatments dramatically influenced chlorophyll content of all the species (Fig. 4). At the control plants chlorophyll content of R. trisperma was the highest, followed by M. azedarach, while it was quite low in R. communis and J. curcas (less than half compared to that in R. trisperma). The increase of Hg concentration caused a significant decrease in the total chlorophyll content in the leaves of all the species. R. trisperma and M. azedarach underwent the highest chlorophyll content reduction due to Hg treatments, while J. curcas was the lowest (Fig. 4). Under the highest Hg treatment (3 mM of Hg), M. azedarach plants had the highest total chlorophyll content followed by R. trisperma, while R. communis and J. curcas had the lowest, even though not significantly different from that of R. trisperma (Fig. 4). The total chlorophyll content of the leaves at 3 mM treatment decreased up to around 45.26-72.69% compared to that of control plants with the smallest decrease found in J. curcas and the biggest decrease in R. trisperma plants (Fig. 4).
There was a very strong negative relationship between MDA and chlorophyll contents in response to mercuric exposure, with the value of coefficient correlation more than 80% (Fig. 5). Among the 4 species, R. trisperma had the steepest curve while the other three had only gentle curve. The lowest correlation between MDA and chlorophyll content showed by R. communis where R2 value was only 0.83, while the other species had R2 value more than 0.95 (Fig. 5).
Principle component analysis of all parameters: The principle component analysis (PCA) was used to distinguish the tolerance level of the 4 species (M. azedarach, R. communis, J. curcas and R. trisperma) to mercuric treatment, even though from the observation, all the 4 species were relatively tolerant to the mercury. The parameters that were analyzed consisted of increased leaves number (ILN), leaf width (LW), shoot dry weight (SW), root dry weight (RW) and total leaf chlorophyll content (CHL). PCA analysis was carried out to discriminate cumulative responses of individual species toward Hg treatment using two-dimensional score plot and biplot.
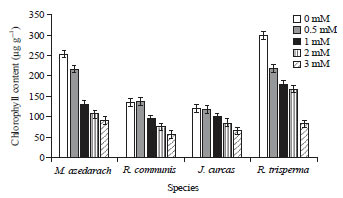 | |
| Fig. 4: | Leaf total chlorophyll content of 4 species treated with Hg at the concentration of 0, 0.5, 1, 2 and 3 mM for 21 days |
Mean+SE of DMRT test at α of 5%, n = 3 | |
From the analysis it was found that the 4 species were divided into 2 groups based on their response toward Hg treatment (Fig. 6). The R. trisperma and M. azedarach was categorized as more tolerant to Hg, while R. communis and J. curcas were categorized as less tolerant to mercuric treatments (Fig. 6).
DISCUSSION
Toxic effects of mercury compounds have been reported in several plants, including Triticum aestivum24, Phaseolus vulgaris L.25, Tomatoes26 and several other plants. According to Ortega-Villasante et al.27 Hg at high concentrations is very toxic to cells thus inducing damage to cells and causing physiological changes. Hg accumulation can also inhibit plant growth, which consequently decreases plant productivity. In this study, the growth of the plants indicated by the value of plant height, leaf number, roots dry weight and shoot dry weight in all 4 species (M. azedarach, R. communis, J. curcas and R. trisperma) decreased dramatically due to mercuric treatments. The dramatic decrease was started even at 0.5 mM of Hg, suggesting that 0.5 mM was the important threshold for mercuric toxicity for the plants.
Plant height, number of leaves, shoot and roots dry weight are most common indicators of plant growth to see the plant's response to the environment. The decrease of all those parameters was occurred due to Hg toxicity to plant tissues. The higher Hg concentration exposed to the plants, the lower plant growth was observed (Table and Fig. 1 and 2). The results also showed that there was a dramatic decrease in the growth rate of vegetative organs including the stem and leaves due to Hg treatments which caused a decrease in shoot and root dry weight. The largest decrease in canopy dry weight was found in the R. communis plant, while the smallest decrease was in R. trisperma plants (Fig. 1 and 2).
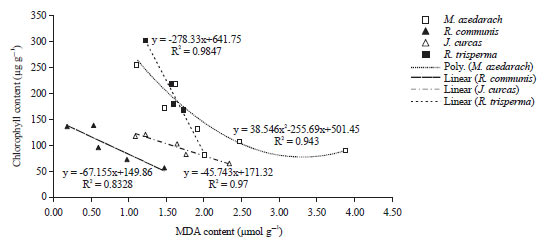 | |
| Fig. 5: | Regressions and coefficient correlations of chlorophyll and MDA contents 4 species in response to mercuric treatment of 0, 0.5, 1, 2 and 3 mM during 21 days exposure |
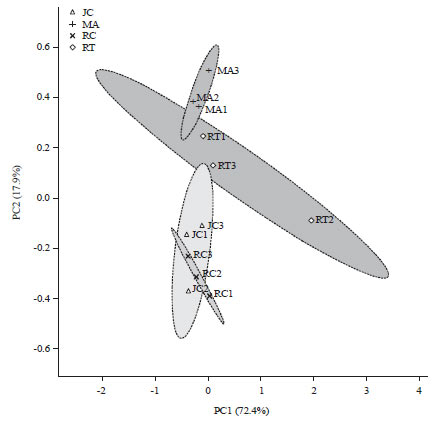 | |
| Fig. 6: | 2-dimensional score plot graph from PCA analysis of 4 species based on anatomical, morphological and physiological characteristics of the plants in response to mercuric treatments |
MA: Melia azedarach, JC: Jatropha curcas, RC: Ricinus communis, RT: Reutealis trisperma | |
This was happened because heavy metals can cause the inhibition of cell division and elongation, water and nutrient absorption and enzymatic activity so that the growth rate was inhibited28. Heavy metals have also been observed to cause inhibition of roots and shoot which reduced crop production29-31. Specifically, Patra and Sharma24 observed that Hg accumulation inhibited root and shoot growth, decreased the ratio of shoots to roots and dry weight and dissolved protein content in the canopy of Triticum aestivum. Suszcynsky and Shann32 also showed that at a concentration of 1.0 μg mL–1, Hg caused inhibition of tobacco roots due to mitotic inhibition, decreased synthesis of cell wall components and photosynthetic activity.
Hg stress also has a significant effect on the anatomical structure of the plant leaves. The results showed a decrease in leaf thickness, upper epidermis, palisade tissues, spongy tissues and lower epidermis in response to the Hg treatments (Table 3). The anatomical structure of R. trisperma plant did not experience much changes compared to the other 3 plants (M. azedarach, R. communis and J. curcas), which significantly reduced (p<0.05) due to Hg treatment. This was probably because R. trisperma did not accumulate more Hg metal in the leaves tissues or the plant had mechanism to resist Hg toxicity. Anatomical alteration is one important mechanism of the plant in response to abiotic stress such as drought33 and heavy metal stress34. The previous report noted that R. trisperma was categorized as resistant plant to metal Pb subjected by gold mine tailing35 and the opportunity to have such resistant also to Hg was much possible. The highest decrease in leaf thickness was found in M. azedarach plants due to Hg treatment compared to the other 3 plants. The previous experiment also reported that heavy metals caused a decrease in the size of epidermal cells which caused a decrease in the epidermis thickness and the thickness of the leaves as a whole in Picris divaricata34. This result also showed that the persistence of leaf anatomical parameters in R. trisperma suggested to its resistance to Hg toxicity up to 3 mM of mercuric nitrate.
Mercuric treatments also influenced a dramatic change in physiological parameters including lipid peroxidation and chlorophyll content in the leaves. Analysis of malondialdehyde (MDA) content showed that mercuric treatments caused significant increase (p<0.05) in MDA content of all the species (Fig. 3), while at the same time the treatments caused dramatic reduction in chlorophyll content (Fig. 4). Several studies have shown that oxidative stress is the main destructive factor in plants due to environmental stress, including heavy metals36,37.
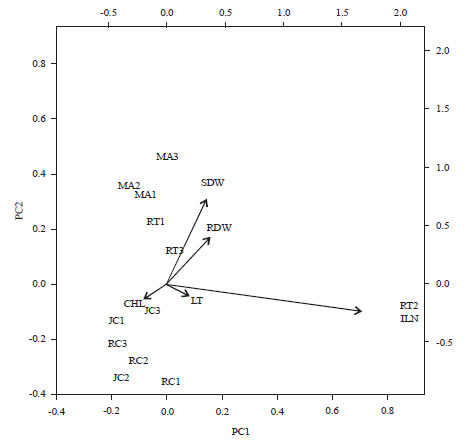 | |
| Fig. 7: | Biplot graph from PCA Analysis of 4 species based on anatomical, morphological and physiological characteristics of the plants in response to mercuric treatments |
MA: Melia azedarach, JC: Jatropha curcas, RC: Ricinus communis, RT: Reutealis trisperma, SDW: Shoot dry weight, RDW: Root dry weight, ILN: Increased leaf number, LT: Leaf thickness, CHL: Chlorophyll content | |
Oxidative stress is normally indicated by lipid peroxidation reactions which are initiated by hydroxyl radicals and lead to the formation of highly reactive peroxyl radicals and can react with other fatty acids38. The chain reaction can be terminated after the combination of 2 radical compounds into non-radical compounds, namely malondialdehyde (MDA) as the end product of lipid peroxidation38,39. MDA content is an important index to evaluate the level of plant damage after stress treatment, which is the main cytotoxic product of lipid peroxidation and an indicator of free radical production40. The higher MDA value indicated that the plants underwent oxidative stress and produced higher reactive oxygen species (ROS) which caused oxidative damage to lipids, proteins, chlorophyll pigments and nucleic acids36,41.
The results of the study showed that the Hg treatment in high concentrations decreased significantly (p<0.05) the total chlorophyll content of the four plant species (Fig. 4). The correlation date between MDA and chlorophyll content in response to Hg treatments also an important indicative that oxidative stress due to Hg toxicity directly caused photosynthetic pigment damage (Fig. 5). This result is also consistent with the experiment carried out by Solymosi et al.42 who reported that Hg stress induced the inhibition of protochlorophyllide photoreduction on wheat leaves, so that the total chlorophyll value of the leaves decreased with increasing Hg concentration. This decrease occurs because heavy metals can inhibit chlorophyll biosynthesis through the inhibition of 2 highly sensitive enzymes, namely α-aminolaevulinic acid (ALA) dehydratase and protochlorophyllide reductase enzymes. Both of these enzymes play a pivotal role in the early and late stages of chlorophyll biosynthesis43. Mercury was also reported to cause magnesium ion replacement in photosynthetic pigments, causing a decrease in photosynthesis rates44.
Among the four non-edible oil producing plants, there was variation response based on morphological, anatomical and physiological characteristics. The study using the principle component analysis (PCA) showed that two of the three principle components had produced Eigen value >1 and had contribution to 90.2% of data variations (Fig. 6). The principle component 1 (PC1) was able to explain 72.4% and the main component 2 (KU2) was able to explain 17.9% of the data variations (Fig. 6). Based on PCA analysis it showed that the R. trisperma and M. azedarach were more tolerant to Hg treatments among the 4 species and was able to survive for 21 days in the treatment (Fig. 6 and 7). This result is also in agreement with the previous result which proved that the R. trisperma was able to grow and survive in gold mine liquid wastes16. R. trisperma plants has also been reported to be able to grow well in the media provided by 100% gold mine tailings treated during 90 days of treatment35.
CONCLUSION
Mercuric treatment caused dramatic inhibition of the growth of four non-edible oil producing plants starting from 0.5 mM of Hg. Hg treatment also caused significant decrease in almost all the leaf tissues width which resulted in a decrease of leaf thickness in M. azedarach, R. communis and J. curcas but not in R. trisperma. The treatments also caused a dramatic increase in leaf MDA content of all species, while at the same time they decreased total chlorophyll content. Principle component analysis categorized the four plant species in response to Hg treatments into two groups, i.e., R. trisperma and M. azedarach in one group which categorized as more tolerant to Hg, while R. communis and J. curcas were in another group.
SIGNIFICANCE STATEMENT
Reutealis trisperma, a non-edible oil producing species, is a good candidate for phytoremediation agent of mercuric contaminated land since this species had ability to grow under higher concentration of mercuric treatment. Utilization of non-edible oil-producing species in phytoremediation program is an important strategy to reduce soil contaminants from gold mining areas. In addition, the plants that able to produce non-edible oil may have second benefit for community around the mining area who involve in phytoremediation program.
ACKNOWLEDGMENTS
This research was funded by The Directorate General of Higher Education, Ministry of Research, Technology and Higher Education Republic of Indonesia through Program of Penelitian Dasar Unggulan Perguruan Tinggi (PDUPT) at IPB University, with the Grant number of 129/SP2H/PTNBH/ DRPM/2018. We also thank to PT Aneka Tambang UPBE Pongkor, Bogor, Indonesia which allow us to use gold mine waste materials for this experiment.
REFERENCES
- Esdaile, L.J. and J.M. Chalker, 2018. The mercury problem in artisanal and small-scale gold mining. Chem.: Eur. J., 24: 6905-6916.
CrossRefDirect Link - Zahir, F., S.J. Rizwi, S.K. Haq and R.H. Khan, 2005. Low dose mercury toxicity and human health. Environ. Toxicol. Pharmacol., 20: 351-360.
CrossRefDirect Link - Singh, S., P. Parihar, R. Singh, V.P. Singh and S.M. Prasad, 2016. Heavy metal tolerance in plants: Role of transcriptomics, proteomics, metabolomics and ionomics. Front. Plant Sci., Vol. 6.
CrossRefDirect Link - Zhou, Z.S., S.Q. Huang, K. Guo, S.K. Mehta, P.C. Zhang and Z.M. Yang, 2007. Metabolic adaptations to mercury-induced oxidative stress in roots of Medicago sativa L. J. Inorg. Biochem., 101: 1-9.
CrossRefDirect Link - Kumar, N., K. Bauddh, S. Kumar, N. Dwivedi, D.P. Singh and S.C. Barman, 2013. Accumulation of metals in weed species grown on the soil contaminated with industrial waste and their phytoremediation potential. Ecol. Eng., 61: 491-495.
CrossRefDirect Link - Alaribe, F.O. and P. Agamuthu, 2015. Assessment of phytoremediation potentials of Lantana camara in Pb impacted soil with organic waste additives. Ecol. Eng., 83: 513-520.
CrossRefDirect Link - Kadirvel, K.K. and M. Jegadeesan, 2014. Mercury and cadmium accumulation in selected weed plants: Implications for phytoremediation. Asian. J. Plant Sci. Res., 4: 1-4.
Direct Link - Liu, W.T., J.C. Ni and Q.X. Zhou, 2013. Uptake of heavy metals by trees: Prospects for phytoremediation. Mater. Sci. Forum, 743-744: 768-781.
CrossRefDirect Link - Marrugo-Negrete, J., S. Marrugo-Madrid, J. Pinedo-Hernandez, J. Durango-Hernandez and S. Diez, 2016. Screening of native plant species for phytoremediation potential at a Hg-contaminated mining site. Sci. Total Environ., 542: 809-816.
CrossRefDirect Link - Setyaningsih, L., A.S. Wulandari and H. Hamim, 2018. Growth of typha grass (Typha angustifolia) on gold-mine tailings with application of arbuscular mycorrhiza fungi. Biodiversitas: J. Biol. Divers., 19: 454-459.
CrossRefDirect Link - Hall, J.L., 2002. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot., 53: 1-11.
CrossRefDirect Link - Altuhaish, A., Hamim and A. Tjahjoleksono, 2014. Biofertilizer effects in combination with different drying system and storage period on growth and production of tomato plant under field conditions. Emir. J. Food Agric., 26: 716-722.
CrossRefDirect Link - Leung, H.M., Z.W. Wang, Z.H. Ye, K.L. Yung, X.L. Peng and K.C. Cheung, 2013. Interactions between arbuscular mycorrhizae and plants in phytoremediation of metal-contaminated soils: A review. Pedosphere, 23: 549-563.
CrossRefDirect Link - Ullah, A., S. Heng, M.F.H. Munis, S. Fahad and X. Yang, 2015. Phytoremediation of heavy metals assisted by plant growth promoting (PGP) bacteria: A review. Environ. Exp. Bot., 117: 28-40.
CrossRefDirect Link - Sarma, H., 2011. Metal hyperaccumulation in plants: A review focusing on phytoremediation technology. J. Environ. Sci. Technol., 4: 118-138.
CrossRefDirect Link - Hamim, M. Hilmi, D. Pranowo, D. Saprudin and L. Setyaningsih, 2017. Morpho-physiological changes of biodiesel producer plants Reutealis trisperma (Blanco) in response to gold-mining wastewater. Pak. J. Biol. Sci., 20: 423-435.
CrossRefDirect Link - Pranowo, D., M. Herma and Syafaruddin, 2015. [The multiple benefits of developing Kemiri Sunan (Reutealis trisperma (Blanco) airy shaw) in degraded land]. Perspektif, 14: 87-101, (In Indonesian).
CrossRefDirect Link - Hodges, D.M., J.M. DeLong, C.F. Forney and R.K. Prange, 1999. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta, 207: 604-611.
CrossRefDirect Link - Quinet, M., D. Vromman, A. Clippe, P. Bertin and H. Lequeux et al., 2012. Combined transcriptomic and physiological approaches reveal strong differences between short- and long-term response of rice (Oryza sativa) to iron toxicity. Plant Cell Environ., 35: 1837-1859.
CrossRefDirect Link - Lichtenthaler, H.K., 1987. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol., 148: 350-382.
CrossRefDirect Link - Xia, J. and D.S. Wishart, 2016. Using MetaboAnalyst 3.0 for comprehensive metabolomics data analysis. Curr. Protocols Bioinform., 55: 10-14.
CrossRefDirect Link - Jolliffe, I.T. and J. Cadima, 2016. Principal component analysis: A review and recent developments. Philos. Trans. R. Soc. A, Vol. 374.
CrossRefDirect Link - Patra, M. and A. Sharma, 2000. Mercury toxicity in plants. Botanical Rev., 66: 379-422.
CrossRefDirect Link - Zengin, F.K. and U.O. Munzuroglu, 2005. Effects of some heavy metals on content of chlorophyll, proline and some antioxidant chemicals in bean (Phaseolus vulgaris L.) seedlings. Acta Biol. Cracoviensia Ser. Bot., 47: 157-164.
Direct Link - Cho, U.H. and J.O. Park, 1999. Changes in hydrogen peroxide content and activities of antioxidant enzymes in tomato seedlings exposed to mercury. J. Plant Biol., 42: 41-48.
CrossRefDirect Link - Ortega-Villasante, C., R. Rellan-Alvarez, F.F. Del Campo, R.O. Carpena-Ruiz and L.E. Hernandez, 2005. Cellular damage induced by cadmium and mercury in Medicago sativa. J. Exp. Bot., 56: 2239-2251.
CrossRefDirect Link - Shahid, M., S. Khalid, G. Abbas, N. Shahid and M. Nadeem et al., 2015. Heavy Metal Stress and Crop Productivity. In: Crop Production and Global Environmental Issues, Hakeem, K.R. (Ed.), Springer, Cham, Switzerland, ISBN: 978-3-319-23162-4, pp: 1-25.
CrossRefDirect Link - Peralta, J.R., J.L. Gardea-Torresdey, K.J. Tiemann, E. Gomez, S. Arteaga, E. Rascon and J.G. Parsons, 2001. Uptake and effects of five heavy metals on seed germination and plant growth in Alfalfa (Medicago sativa L.). Bull. Environ. Contam. Toxicol., 66: 727-734.
CrossRefDirect Link - Setyaningsih, L., Y. Setiadi, S.W. Budi, H. Hamim and D. Sopandie, 2018. Jabon (Anthocephalus cadamba Roxb) potency for remediating lead (Pb) toxicity under nutrient culture condition. Biotropia: Southeast Asian J. Trop. Biol., 25: 64-71.
Direct Link - Suszcynsky, E.M. and J.R. Shann, 1995. Phytotoxicity and accumulation of mercury in tobacco subjected to different exposure routes. Environ. Toxicol. Chem.: Int. J., 14: 61-67.
CrossRefDirect Link - Hamim, S. Banon and D. Dorly, 2016. Comparison of physiological and anatomical changes of C3 (Oryza sativa [L.]) and C4 (Echinochloa crusgalli [L.]) leaves in response to drought stress. IOP Conf. Ser.: Earth Environ. Sci., Vol. 31.
CrossRefDirect Link - Tang, L., R.R. Ying, D. Jiang, X.W. Zeng, J.L. Morel, Y.T. Tang and R.L. Qiu, 2013. Impaired leaf CO2 diffusion mediates Cd-induced inhibition of photosynthesis in the Zn/Cd hyperaccumulator Picris divaricata. Plant Physiol. Biochem., 73: 70-76.
CrossRefDirect Link - Hilmi, M., H. Hamim, Y.C. Sulistyaningsih and Taufikurahman, 2018. Growth, histochemical and physiological responses of non-edible oil producing plant (Reutealis trisperma) to gold mine tailings. Biodiversitas: J. Biol. Divers., 19: 1294-1302.
CrossRefDirect Link - Wu, F., G. Zhang and P. Dominy, 2003. Four barley genotypes respond differently to cadmium: Lipid peroxidation and activities of antioxidant capacity. Environ. Exp. Bot., 50: 67-78.
CrossRefDirect Link - Shanker, A.K., M. Djanaguiraman, R. Sudhagar, C.N. Chandrashekar and G. Pathmanabhan, 2004. Differential antioxidative response of ascorbate glutathione pathway enzymes and metabolites to chromium speciation stress in green gram (Vigna radiata (L.) R.Wilczek. cv CO 4) roots. Plant Sci., 166: 1035-1043.
CrossRefDirect Link - Blokhina, O., E. Virolainen and K.V. Fagerstedt, 2003. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot., 91: 179-194.
CrossRefPubMedDirect Link - Valko, M., H. Morris and M.T.D. Cronin, 2005. Metals, toxicity and oxidative stress. Curr. Med. Chem., 12: 1161-1208.
CrossRefPubMedDirect Link - Fu, J. and B. Huang, 2001. Involvement of antioxidants and lipid peroxidation in the adaptation of two cool-season grasses to localized drought stress. Environ. Exp. Bot., 45: 105-114.
CrossRefPubMedDirect Link - Hegedus, A., S. Erdei and G. Horvath, 2001. Comparative studies of H2O2 detoxifying enzymes in green and greening barley seedlings under cadmium stress. Plant Sci., 160: 1085-1093.
CrossRefDirect Link - Solymosi, K., K. Lenti, B. Mysliwa-Kurdziel, J. Fidy, K. Strzalka and B. Boddi, 2004. Hg2+ reacts with different components of the NADPH: protochlorophyllide oxidoreductase macrodomains. Plant Biol., 6: 358-368.
CrossRefDirect Link - De Fillipis, L.F., R. Hampp and H. Ziegler, 1981. The effect of sublethal concentrations of zinc, cadmium and mercury on Euglena. Adenylates and energy charge. Zeitschrift Pflanzenphysiologie, 103: 1-7.
CrossRefDirect Link - Kupper, H., F. Kupper and M. Spiller, 1998. In situ detection of heavy metal substituted chlorophylls in water plants. Photosynth. Res., 58: 123-133.
CrossRefDirect Link