
Mohammad Nasir Khan
Department of Biology, Faculty of Science, University of Tabuk, Tabuk 71491, Saudi Arabia
LiveDNA: 91.14257
Fahad Mohammed Alzuaibr
Department of Biology, Faculty of Science, University of Tabuk, Tabuk 71491, Saudi Arabia
Asian Journal of Plant Sciences
Year: 2018 | Volume: 17 | Issue: 4 | Page No.: 213-221







ABSTRACT
Background and Objectives: In recent years nano-materials have emerged as an important tools in manipulating crop performance worldwide. Also hydrogen sulfide (H2S) has gained substantial attention of plant biologists. Present study was planned to investigate the effect of nano-titanium dioxide (nTiO2) on the synthesis of H2S and their role in the tolerance of Eruca sativa plants to drought stress. Materials and Methods: Three week old plants of Eruca sativa were sprayed with 20 mg L–1 nTiO2 and 1 mM hypotaurine (HT, an H2S scavenger) then plants were subjected to drought stress by withholding water and nutrient supply for one week except for the control which received double distilled water (DDW) only. The treatments were given as: (1)DDW (Control), (2) nTiO2, (3) Drought stress (DS), (4) nTiO2+DS, (v) nTiO2+HT+DS. Plants treated with DDW only were considered as control. Results: Results showed that drought stress induced the generation of hydrogen peroxide (H2O2), thiobarbituric acid reactive substances (TBARS), electrolyte leakage (ELKG) and caused reduction in leaf relative water content (LRWC). At the same time drought-stressed plants also showed enhanced activities of antioxidant enzymes [superoxide dismutase (SOD), peroxidase (POX) and catalase (CAT)] and accumulation of osmolytes [proline (Pro), glycine betaine (GB)]. Moreover, drought-stressed plants pre-treated with nTiO2 showed further enhancement in the activities of antioxidant enzymes and accumulation of osmolytes that resulted in reduced H2O2 content, TBARS, ELKG and improved LRWC. Furthermore, nTiO2 also enhanced the synthesis of H2S and cysteine. Role of H2S in drought stress tolerance was confirmed using H2S scavenger hypotaurine (HT). Conclusion: Results showed that application of HT along with nTiO2 to drought stressed pants suppressed H2S content and plants showed weak tolerance against drought stress. Therefore, these results suggest that nTiO2-induced synthesis of H2S induces drought tolerance capacity of plants through enhancing the activities of antioxidant enzymes and accumulation of osmolytes.
PDF Abstract XML References Citation
Received: August 27, 2018;
Accepted: November 02, 2018;
Published: December 26, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Mohammad Nasir Khan and Fahad Mohammed Alzuaibr, 2018. Nano-titanium Dioxide-induced Synthesis of Hydrogen Sulfide and Cysteine Augment Drought Tolerance in Eruca sativa. Asian Journal of Plant Sciences, 17: 213-221.
DOI: 10.3923/ajps.2018.213.221
URL: https://scialert.net/abstract/?doi=ajps.2018.213.221
DOI: 10.3923/ajps.2018.213.221
URL: https://scialert.net/abstract/?doi=ajps.2018.213.221
INTRODUCTION
Drought has been considered as one of the most damaging environmental stresses that causes severe losses to crops across the globe. Loss of water is the signature effect of drought that disrupts water relations and suppresses water use efficiency of plants resulting in osmotic stress. Lower water status causes closure of stomata, turgor loss, reduced photosynthetic activity and suppressed carbon assimilation1,2, leading to poor dry matter accumulation in affected plants3,4. In addition, drought stress creates an imbalance between production and scavenging of reactive oxygen species (ROS) that causes excessive generation of ROS such as superoxide (O2∙–), hydrogen peroxide (H2O2), hydroperoxyl radical (HO2.), singlet oxygen (1O2) and hydroxyl radical (OH∙). Over accumulation of ROS creates oxidative stress that causes oxidation of membrane lipids, proteins and nucleic acids2,5. Being sessile in nature plants are always exposed to several abiotic stresses. Under such circumstances plants are provided with various types of defense mechanisms. To counter osmotic stress plants synthesize osmolytes such as proline (Pro), glycinebetaine (GB) which provide osmotic adjustment through stabilizing biomolecules and maintaining membrane stability6-9. Moreover, to invalidate oxidative stress plants possess a system of antioxidant enzymes such as superoxide dismutase (SOD), peroxidase (POX) and catalase (CAT). It is well documented that SOD dismutates O2∙– radicals to H2O2, whereas POX and CAT convert H2O2 into water and oxygen. Activation of these defense systems, in response to abiotic stresses, is carried out by a network of signaling molecules. Of these, hydrogen sulfide (H2S) has emerged as an important signaling molecule that mediates responses to biotic and abiotic stresses in plants. It has been studied that H2S protects plants against abiotic stress induced oxidative and osmotic stress through enhancing the activities of antioxidant enzymes and accumulation of osmolytes10,11. In plants H2S is synthesized by the degradation of cysteine (Cys), a sulfur containing precursor of various biomolecules12. Therefore, endogenous level of H2S depends on the availability of Cys in the cells.
Among the nanomaterials (NMs), nano-titanium dioxide (nTiO2) has been shown to play significant role in growth and development of crop plants. In addition to their role in growth and development of plants, nTiO2 also plays vital role in the protection of plants against various abiotic stresses such as drought, salinity, cold, heat, metal and UV radiation13. It has been shown that nTiO2 counters drought stress14, mimics the activities of antioxidant enzymes and scavenges ROS13. Drought-stressed plants treated with nTiO2 exhibit improved morphological and physiological attributes14.
Although, various studies have been carried out to investigate the role of NMs in plants, meager information is available regarding the effect of nTiO2 on the level of H2S and Cys under drought stress. Therefore, considering the important role of nTiO2 and H2S in plants the objective of the present work was to investigate whether exogenous nTiO2 could affect endogenous levels of H2S and Cys and to explore their interactive role in the activation of antioxidant defense system and osmolytes accumulation in relation to the tolerance of Eruca sativa plants to drought stress. To achieve the objective a pot experiment was carried out under natural environmental conditions using hypotaurine (HT) as H2S scavenger.
MATERIALS AND METHODS
Plant materials and treatments: Seeds of arugula (Eruca sativa Mill.) were purchased from local market of Tabuk. Healthy and uniform seeds were surface sterilized with 1% sodium hypochlorite for 10 min, then vigorously rinsed with double distilled water (DDW). On March 14, 2018, surface sterilized seeds were sown in plastic pots (20 cm diameter and 20 cm height) containing soil/vermiculite (1:1) mixture. The plants were allowed to grow for 3 weeks under natural illuminated conditions with average day/night temperature 26/8±3°C. All the pots were supplied with 50 mL of Raukura’s nutrient solution15 daily. After three weeks (on April 3, 2018), foliar spray of nTiO2 at the rate of 20 mg L–1 and 1 mM hypotaurine (HT, an H2S scavenger) was given and then plants were subjected to drought stress by withholding water and nutrient supply for 1 week (from April 4 to April 10, 2018), except for the control which received DDW only. The treatments were given as: (1) Double distilled water (DDW: control), (2)nTiO2, (3)Drought stress (DS), (4)nTiO2+DS, (5)nTiO2+HT+DS. Plants treated with DDW only were considered as control. Each treatment was replicated three times and each replicate was consisted of three plants. After one week of drought when the plants were four weeks old, the effect of nTiO2 on drought stress was tested (on April 11, 2018) by measuring leaf relative water content (LRWC), electrolyte leakage (ELKG), hydrogen peroxide (H2O2) content, thiobarbituric acid reactive substances (TBARS) and proline (Pro) and glycine betaine (GB) content. Activities of antioxidant enzymes superoxide dismutase (SOD), peroxidase (POX) and catalase (CAT) were analyzed. Concentration of cysteine (Cys), hydrogen sulfide (H2S) and chlorophyll (Chl-a, Chl-b, total Chl and Chl a/b ratio) was also estimated.
Measurement of physiological and biochemical parameters: Leaf relative water content (LRWC) was measured by adopting the method of Yamasaki and Dillenburg16. Fresh weight (FW), dry weight (DW) and turgid weight (TW) of leaves was measured and LRWC was calculated using the equation below. The values for FW, TW and DW were used to calculate LRWC using the equation below:
Effect of drought stress on membrane permeability was assessed in term of percentage of electrolyte leakage (ELKG) by the method of Lutts et al.17. Hydrogen peroxide (H2O2) content was determined according to Velikova et al.18. The content of H2O2 was calculated based on a standard curve and was expressed as μmol g–1 leaf DW. Lipid peroxidation was determined by measuring the content of thiobarbituric acid reactive substances (TBARS) as described by Cakmak and Horst19. Content of TBARS was expressed as nmol g–1DW. Proline (Pro) and glycine betaine (GB) content was determined according to the method of Bates et al.20 and Grieve and Grattan21, respectively.
Assay of antioxidant enzymes: A crude enzyme extract was prepared prior to determination of antioxidant enzyme activities. Fresh leaves were homogenized with three volumes (w/v) of an ice-cold extraction buffer (50 mM Tris-HCl, pH 7.8, 1 mM EDTA, 1 mM MgCl2 and 1.5% (w/w) polyvinylpyrrolidone). The homogenate was centrifuged at 15,000 g for 20 min at 4°C. The supernatant was used as the crude extract for the assay of activities of superoxide dismutase (SOD), peroxidase (POX) and catalase (CAT).
Activities of superoxide dismutase (SOD; EC 1.15.1.1), peroxidase (POX; EC 1.11.1.7) and catalase (CAT; EC 1.11.1.6) were determined by the method of Beauchamp and Fridovich22, Upadhyaya et al.23 and Cakmak and Marschner24, respectively.
Measurement of hydrogen sulfide (H2S) and cysteine (Cys) content: The method of Nashef et al.25 was adopted to estimate the concentration of H2S. Concentration of H2S was expressed in n mol g–1 DW. Cysteine content was determined according to Gaitonde26 as described by Riemenschneider et al.27 with slight modifications. The amount of Cys was calculated using pure Cys as standard and the result was expressed as nmol g–1 DW.
Estimation of chlorophyll (Chl) content: Chlorophyll (Chl) content was estimated using the method of Lichtenthaler and Buschmann28. The optical density of the pigment solution was recorded at 662 and 645 nm to determine Chl a and Chl b, respectively using a spectrophotometer.
Statistical analysis: Analysis of variance (ANOVA) was performed to evaluate the significance of the treatment means. The data were expressed as the Mean±standard error and the data were analyzed statistically using SPSS ver. 17 statistical software (SPSS Inc., Chicago, IL, USA). Treatment means were statistically compared by Duncan's Multiple Range Test (DMRT) at p<0.05% level. Each treatment was replicated three times and each replicate was consisted of three plants.
RESULTS
Effect of nTiO2 and drought stress on LRWC and membrane permeability: Hydration level of plants was assessed in term of LRWC. Perusal of the data showed that under drought stress plants exhibited 56.7% LRWC which was lower as compared with the control. However, drought-stressed plants pre-treated with nTiO2 showed 78.5% LRWC which was significantly higher than drought-stressed plants (Fig. 1a).
Effect of drought and nTiO2 on membrane permeability was tested by measuring ELKG. Drought stress caused a significant increase in ELKG compared to the control (Fig. 1a). However, pre-treatment with nTiO2 alleviated the effect of drought and caused reduction in the level of ELKG as compared with the drought-suffered plants (Fig. 1a).
Effect of nTiO2 and drought stress on H2O2 content and lipid peroxidation: The results exhibited that exposure of plants to drought caused a significant increase in H2O2 content than the control. However, exposure of nTiO2-treated plants to drought (nTiO2+DS) showed a considerable reduction in H2O2 content than drought-stressed plants not treated with nTiO2 (Fig. 1b).
The results showed that drought stress instigated peroxidation of membrane lipids as reflected by increased level of TBARS (Fig. 1c). Value of TBARS in drought-stressed plants was about two-folds higher than the control plants. Nevertheless, drought-stressed plants pre-treated with nTiO2 (nTiO2+DS) exhibited a decrease in the synthesis of TBARS than the stressed plants (DS) (Fig. 1c).
Effect of nTiO2 and drought stress on Pro and GB content: It was evident from Fig. 1d and e that drought stress enhanced the accumulation of Pro and GB content. Moreover, drought-stressed plants supplemented with nTiO2 (nTiO2+DS) showed a further increase in Pro and GB content as compared with the drought-suffered plants (Fig. 1d and e).
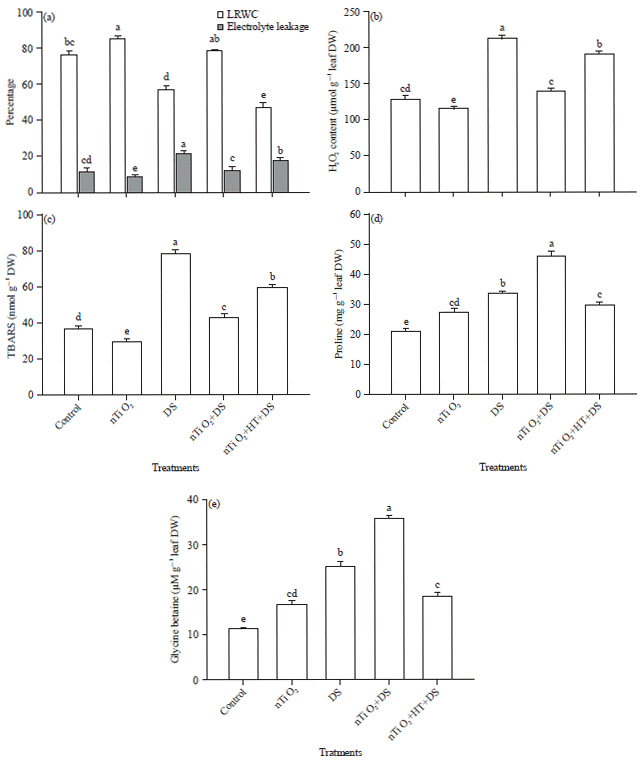 | |
| Fig. 1(a-e): | Effect of nano-TiO2 on (a) LRWC and electrolyte leakage, (b) H2O2 content, (c) TBARS, (d) Pro content and (e) GB content of Eruca sativa under drought stress. Average of three determinations is presented with bars indicating S.E. |
| Bars followed by the same letter do not differ statistically at p<0.05 (Duncan Multiple Range Test). DDW: Control, nTiO2: 20 mg L–1 nano-TiO2, DS: Drought stress, HT, an H2S scavenger: 1 mM hypotaurine | |
Effect of nTiO2 and drought stress on the activities of antioxidant enzymes: Analysis of the data showed that plants under drought stress exhibited higher activities of antioxidant enzymes (SOD, POX and CAT) than the control plants (Fig. 2a, b). In addition, application of 20 mg L–1 of nTiO2 further enhanced the activities of these antioxidant enzymes than drought-stressed plants grown without nTiO2 (DS) (Fig. 2a, b).
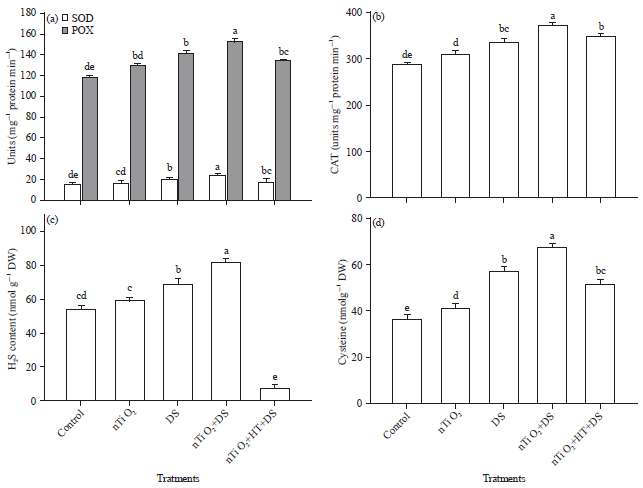 | |
| Fig. 2(a-d): | Effect of nano-TiO2 on (a) SOD and POX activities, (b) CAT activity, (c) H2S content and (d) Cysteine content of Eruca sativa under drought stress. Average of three determinations is presented with bars indicating S.E |
| Bars followed by the same letter do not differ statistically at p<0.05 (Duncan Multiple Range Test). DDW: Control, nTiO2: 20 mg L–1 nano-TiO2, DS: Drought stress, HT, an H2S scavenger: 1 mM hypotaurine | |
| Table 1: | Effect of nano-TiO2 on chlorophyll content of Eruca sativa under drought stress |
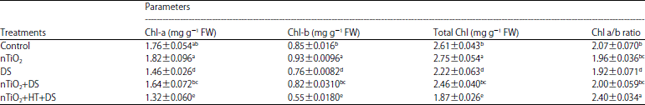 | |
Average of three determinations is presented with ±indicating standard error. Values with the same letters with in a column do not differ statistically at p<0.05 (Duncan Multiple Range Test). DDW: Control, nTiO2: 20 mg L–1 nano-TiO2, DS: Drought stress, HT, an H2S scavenger: 1 mM hypotaurine
Effect of nTiO2 and drought stress on H2S and Cys content: Plants exposed to drought for one week synthesized more H2S and Cys than the control plants (Fig. 2c, d). Moreover, drought-stressed plants treated with nTiO2 showed further enhancement in H2S and Cys levels than drought stressed plants (Fig. 2c, d).
Effect of nTiO2 and drought stress on Chl content: Results showed that exposure of plants to drought stress caused a significant reduction in Chl-a, Chl-b and total Chl content and Chl a/b ratio than the control (Table 1). However, drought-suffered plants pre-treated with nTiO2 (nTiO2+DS) countered detrimental effects of drought and showed an increase in Chl-a, Chl-b and total Chl content and Chl a/b ratio than drought-stressed plants that did not receive nTiO2 (Table 1).
DISCUSSION
Loss of water in the plants is hallmark of drought stress which was assessed in terms of LRWC. Perusal of the data showed that under drought stress plants exhibited lower LRWC (Fig. 1a). It is well established that drought stress adversely affects water relations that lead to reduction in leaf water potential, turgor loss and stomatal closure. All these together reduce water uptake capacity of plants29,30 as witnessed by lower LRWC. Maintenance of optimum water status of plants is highly desirable for normal functioning of cellular system under abiotic stresses. To cope with deprived water status, plants accumulate osmolytes such as Pro and GB that maintain normal hydration level of plants31,32. Drought stress enhanced the accumulation of Pro and GB content (Fig. 1d, e). But in spite of increase in the level of these osmolytes, a decrease in LRWC was noticed under drought stress. It indicates that increased concentration of Pro and GB content was not sufficient to counter drought stress-induced decrease in LRWC. However, drought-stressed plants treated with nTiO2 showed a further increase in Pro and GB concentration that possibly increased osmotic pressure resulting in the enhancement of water uptake capacity of treated plants as shown by improved LRWC (Fig. 1a). Furthermore, it was cleared from the results that nTiO2 accelerated the synthesis of H2S which had been shown to induce the activity of Pro-synthesizing enzymeΔ1-pyrroline-5-carboxylate synthetase and reduces the activity of Pro degrading enzyme, Pro-dehydrogenase33 that resulted in enhanced accumulation of Pro. H2S also enhances the activity of betaine aldehyde dehydrogenase, a key enzyme in the biosynthesis of betaine which induces GB synthesis that stabilizes biological membranes and protects the plants against adverse effects of abiotic stress34,35. Involvement of H2S in osmotic adjustment of stressed plants was further confirmed when H2S scavenger HT was applied that decreased Pro and GB content to the level recorded from drought-stressed plants.
Onset of drought stress induces generation of ROS such as H2O2 that creates oxidative stress36,37. To cope with oxidative stress, plants possess a system of antioxidant enzymes which continuously scavenge ROS and maintain the normal level of ROS. However, under suppressed activities of antioxidant enzymes the rate of ROS production exceeds the rate of ROS scavenging which results in over production of ROS. Excessive accumulation of ROS causes peroxidation of membrane lipids and leakage of electrolytes38,39 (Fig. 1a, c). The results showed that plants under drought stress enhanced the activities of antioxidant enzymes (SOD, POX and CAT) but a parallel increase in H2O2 content was also noticed. It shows that increase in plants’ antioxidant defense system was not efficient to counter oxidative stress. However, drought-stressed plants pre-treated with nTiO2 showed a further increase in the activities of antioxidant enzymes to a level which was effective in scavenging ROS as witnessed by decreased levels of H2O2 content coupled with reduced ELKG and TBARS (Fig. 1a-c). It confirms that nTiO2 can regulate the activities of antioxidant enzymes to the level required to counter ROS and can also modulate ROS dependent signaling pathways40 leading to significant enhancement in plant growth41,42. These results also corroborated the findings of Khan43. Moreover, nTiO2 enhanced the synthesis of H2S which has been shown to induce antioxidant defense system of plants10,44. On the contrary, application of H2S scavenger HT suppressed the activities of antioxidant enzymes and an increase in the generation of H2O2 content was noticed which galvanized the leakage of electrolytes and levels of TBARS (Fig. 1a-c). It validates the role of H2S against oxidative stress.
Exposure of plants to drought caused a significant increase in H2S content (Fig. 2c). It has been already observed that drought up-regulates the expression levels of H2S-synthesizing genes that induce production of H2S and tolerance to drought stress45. Although, enhanced concentration of H2S improved the activities of antioxidant enzymes and Pro and GB content but at the same time an increase in H2O2 content, ELKG and TBARS was also noticed with an antiparallel decrease in LRWC and Chl content. It shows that enhanced level of H2S was not sufficient to provide complete protection against drought-induced impairments. However, application of nTIO2 further enhanced the accumulation of H2S to a level required to boost the activities of antioxidant enzymes and accumulation of Pro and GB that resulted in reduced H2O2 content, ELKG and TBARS and increased LRWC. In order to maintain uninterrupted synthesis of H2S under stress conditions, there should be continuous supply of Cys. Application of nTIO2 not only increased the synthesis of H2S but also of Cys and thus Cys pool was maintained that assisted the plants to synthesize more H2S for proper functioning of cellular system under stressful conditions. These results were in agreement with the findings of Khan et al.10 who observed that osmotic stress enhanced the activities of H2S and Cys-synthesizing enzymes that contributed to enhanced levels of H2S and Cys, respectively. Based on these observations, it can be speculated that nTiO2 might had enhanced H2S and Cys synthesis by accelerating the activities of H2S and Cys-synthesizing enzymes.
Drought stress also caused a significant reduction in Chl content (Table 1). As mentioned earlier that drought stress induced synthesis of ROS (H2O2 content). Excessive accumulation of ROS causes lipid peroxidation, leakage of electrolytes and photo-oxidative damage to chlorophyll46,47, instability of protein complexes and increase in the activity of Chl-degrading enzyme chlorophyllase48 leading to destruction of Chl. It is noteworthy here that drought stress caused higher decrease in Chl-a than Chl-b which indicates that Chl-a was more sensitive to drought. However, drought-stressed plants pre-treated with nTiO2 showed higher concentration of Chl. Khan et al.43 also observed that nTiO2 induces Chl content under salt stress. Moreover, nTiO2 also improved the synthesis of H2S which causes decline in H2O2 content, ELKG and TBARS and increase in LRWC through enhancing the activities of antioxidant enzymes and accumulation of Pro and GB. All these together might have contributed to the alleviation of photo-oxidative damage and reduction in the activity of chlorophyllase that resulted in improved Chl concentration. These results are supported by the findings of Zhang et al.49 and Wei et al.50 who observed that H2S plays an active role in suppressing Chl degradation.
CONCLUSION
The results showed that, nTiO2 induced the synthesis of H2S and Cys in drought stressed plants. Improved level of H2S and Cys together with nTiO2, alleviated drought stress by inducing the activities of antioxidant enzymes viz. SOD, POX and CAT and accumulation of Pro and GB content. Activated antioxidant enzymes significantly countered oxidative stress by suppressing the generation of H2O2 that resulted in the reduction of lipid peroxidation and electrolyte leakage. Similarly, enhanced synthesis of Pro and GB maintained normal osmotic pressure that facilitated drought-stressed plants to uptake more water that was reflected in the form of increased LRWC and Chl content.
SIGNIFICANCE STATEMENT
This study was carried out to explore the interactive role of nTiO2, H2S and Cys in the protection of Eruca sativa plants against drought stress. The results showed that application of nTiO2 to drought stressed plants enhanced the synthesis of H2S and Cys. Improved level of H2S gave protection to the plants against detrimental effects of drought stress through enhancing the activities of antioxidant enzymes and accumulation of osmolytes. Role of H2S in the tolerance of plants to drought stress was confirmed by the application of H2S scavenger HT.
ACKNOWLEDGMENT
This study was supported by the Deanship of Scientific Research (DSR), University of Tabuk, Saudi Arabia (Project No. 0095-1438-S).
REFERENCES
- Chaves, M.M., J.S. Pereira, J. Maroco, M.L. Rodrigues and C.P.P. Ricardo et al., 2002. How plants cope with water stress in the field? Photosynthesis and growth. Ann. Bot., 89: 907-916.
CrossRefDirect Link - Signorelli, S., F.J. Corpas, O. Borsani, J.B. Barroso and J. Monza, 2013. Water stress induces a differential and spatially distributed nitro-oxidative stress response in roots and leaves of Lotus japonicas. Plant Sci., 201: 137-146.
CrossRefDirect Link - Li, Q., X. Zhang, Q. Lv, D. Zhu and T. Qiu et al., 2017. Physcomitrella patens dehydrins (PpDHNA and PpDHNC) confer salinity and drought tolerance to transgenic Arabidopsis plants. Front. Plant Sci., Vol. 8.
CrossRefDirect Link - Mathobo, R., D. Marais and J.M. Steyn, 2017. The effect of drought stress on yield, leaf gaseous exchange and chlorophyll fluorescence of dry beans (Phaseolus vulgaris L.). Agric. Water Manage., 180: 118-125.
CrossRefDirect Link - Foyer, C.H. and G. Noctor, 2000. Oxygen processing in photosynthesis: Regulation and signalling. New Phytol., 146: 359-388.
CrossRefDirect Link - Gadallah, M.A.A., 1999. Effects of proline and glycinebetaine on Vicia faba responses to salt stress. Biol. Plant., 42: 249-257.
CrossRefDirect Link - Makela, P., J. Karkkainen and S. Somersalo, 2000. Effect of glycinebetaine on chloroplast ultrastructure, chlorophyll and protein content and RuBPCO activities in tomato grown under drought or salinity. Biol. Plant., 43: 471-475.
CrossRefDirect Link - Bandurska, H., A. Gorny and M. Zielezinska, 2008. Effects of water deficit on the relative water content, proline accumulation and injury of cell membranes in leaves of old and modern cultivars of winter wheat. Zeszyty Problemowe Postepow Nauk Rolniczych, Vol. 524.
Direct Link - DaCosta, M. and B. Huang, 2006. Osmotic adjustment associated with variation in bentgrass tolerance to drought stress. J. Am. Soc. Horticult. Sci., 131: 338-344.
Direct Link - Khan, M.N., M. Mobin, K.A. Zahid and H.S. Manzer, 2017. Nitric oxide-induced synthesis of hydrogen sulfide alleviates osmotic stress in wheat seedlings through sustaining antioxidant enzymes, osmolyte accumulation and cysteine homeostasis. Nitric Oxide, 68: 91-102.
CrossRefDirect Link - Li, Z.G. and L.P. Zhu, 2015. Hydrogen sulfide donor sodium hydrosulfide-induced accumulation of betaine is involved in the acquisition of heat tolerance in maize seedlings. Brazil. J. Bot., 38: 31-38.
CrossRefDirect Link - Alvarez, C., M.A. Bermudez, L.C. Romero, C. Gotor and I. Garcia, 2012. Cysteine homeostasis plays an essential role in plant immunity. New Phytol., 193: 165-177.
CrossRefDirect Link - Khan, M.N., M. Mobin, K.A. Zahid, K.A. ALMutairi and H.S. Zahid, 2017. Role of nanomaterials in plants under challenging environments. Plant Physiol. Biochem., 110: 194-209.
CrossRefDirect Link - Aghdam, M.T.B., H. Mohammadi and M. Ghorbanpour, 2016. Effects of nanoparticulate anatase titanium dioxide on physiological and biochemical performance of Linum usitatissimum (Linaceae) under well-watered and drought stress conditions. Brazil. J. Bot., 39: 139-146.
CrossRefDirect Link - Smith, G.S., C.M. Johnston and I.S. Cornforth, 1983. Comparison of nutrient solutions for growth of plants in sand culture. New Phytol., 94: 537-548.
CrossRefDirect Link - Yamasaki, S. and L.R. Dillenburg, 1999. Measurements of leaf relative water content in Araucaria angustifolia. Revista Brasileira de Fisiologia Vegetal, 11: 69-75.
Direct Link - Lutts, S., J.M. Kinet and J. Bouharmont, 1995. Changes in plant response to NaCl during development of rice (Oryza sativa L.) varieties differing in salinity resistance. J. Exp. Bot., 46: 1843-1852.
Direct Link - Velikova, V., I. Yordanov and A. Edreva, 2000. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci., 151: 59-66.
CrossRefDirect Link - Cakmak, I. and W.J. Horst, 1991. Effect of aluminium on lipid peroxidation, superoxide dismutase, catalase and peroxidase activities in root tips of soybean (Glycine max). Physiol. Plant., 83: 463-468.
CrossRefDirect Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Grieve, C.M. and S.R. Grattan, 1983. Rapid assay for determination of water soluble quaternary ammonium compounds. Plant Soil, 70: 303-307.
CrossRefDirect Link - Beauchamp, C. and I. Fridovich, 1971. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem., 44: 276-287.
CrossRefPubMedDirect Link - Upadhyaya, A., D. Sankhla, T.D. Davies, N. Sankhla and B.N. Smith, 1985. Effect of paclobutrazol on the activities of some enzymes of activated oxygen metabolism and lipid peroxidation in senescing soybean leaves. J. Plant Physiol., 121: 453-461.
CrossRefDirect Link - Nashef, A.S., D.T. Osuga and R.E. Feeney, 1977. Determination of hydrogen sulfide with 5, 5'-dithiobis-(2-nitrobenzoic acid), N-ethylmaleimide and parachloromercuribenzoate. Anal. Biochem., 79: 394-405.
CrossRefDirect Link - Riemenschneider, A., K. Riedel, R. Hoefgen, J. Papenbrock and H. Hesse, 2005. Impact of reduced O-acetylserine (thiol) lyase isoform contents on potato plant metabolism. Plant Physiol., 137: 892-900.
CrossRefDirect Link - Lichtenthaler, H.K. and C. Buschmann, 2001. Chlorophylls and Carotenoids: Measurement and Characterization by UV-VIS Spectroscopy. In: Current Protocols in Food Analytical Chemistry, Wrolstad, R.E., T.E. Acree, H. An, E.A. Decker and M.H. Penner et al. (Eds.)., John Wiley and Sons, New York, USA, pp: F4.3.1-F4.3.8.
Direct Link - Jaleel, C.A., P. Manivannan, A. Wahid, M. Farooq, H.J. Al-Juburi, R. Somasundaram and R. Panneerselvam, 2009. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol., 11: 100-105.
Direct Link - Haider, M.S., M.M. Kurjogi, M. Khalil-ur-Rehman, T. Pervez and J. Songtao et al., 2018. Drought stress revealed physiological, biochemical and gene-expressional variations in 'Yoshihime' peach (Prunus persica L) cultivar. J. Plant Int., 13: 83-90.
CrossRefDirect Link - Siddiqui, M.H., M.N. Khan, F. Mohammed and M.M.A. Khan, 2008. Role of nitrogen and gibberellin (GA3) in the regulation of enzyme activities and in osmoprotectant accumulation in Brassica juncea L. under salt stress. J. Agron. Crop Sci., 194: 214-224.
CrossRefDirect Link - Khan, M.N., M.H. Siddiqui, F. Mohammad, M. Naeem and M.M.A. Khan, 2010. Calcium chloride and gibberellic acid protect linseed (Linum usitatissimum L.) from NaCL stress by inducing antioxidative defence system and osmoprotectant accumulation. Acta Physiol. Plant, 32: 121-132.
CrossRefDirect Link - Li, Z.G., X.J. Ding and P.F. Du, 2013. Hydrogen sulfide donor sodium hydrosulfide-improved heat tolerance in maize and involvement of proline. J. Plant Physiol., 170: 741-747.
CrossRefDirect Link - Khan, M.N., M.H. Siddiqui, F. Mohammad and M. Naeem, 2012. Interactive role of nitric oxide and calcium chloride in enhancing tolerance to salt stress. Nitric Oxide, 27: 210-218.
CrossRefDirect Link - Sakamoto, A. and N. Murata, 2000. Genetic engineering of glycinebetaine synthesis in plants: Current status and implications for enhancement of stress tolerance. J. Exp. Bot., 51: 81-88.
CrossRefDirect Link - Khan, A., Y. Anwar, M.M. Hasan, A. Iqbal and M. Ali et al., 2017. Attenuation of drought stress in brassica seedlings with exogenous application of Ca2+ and H2O2. Plants, Vol. 6.
CrossRef - De Carvalho, M.H.C., 2008. Drought stress and reactive oxygen species: Production, scavenging and signaling. Plant Signal Behav., 3: 156-165.
CrossRefPubMedDirect Link - Tatari, M., R.F. Ghazvini, A. Mousavi and G. Babaei, 2018. Comparison of some physiological aspects of drought stress resistance in two ground cover genus. J. Plant Nutr., 41: 1215-1226.
CrossRefDirect Link - Guo, Y.Y., H.Y. Yu, M.M. Yang, D.S. Kong and Y.J. Zhang, 2018. Effect of drought stress on lipid peroxidation, osmotic adjustment and antioxidant enzyme activity of leaves and roots of Lycium ruthenicum Murr. Seedling, Russian J. Plant Physiol., 65: 244-250.
CrossRefDirect Link - Mohammadi, R., R. Maali-Amiri and N.L. Mantri, 2014. Effect of TiO2 nanoparticles on oxidative damage and antioxidant defense systems in chickpea seedlings during cold stress. Russian J. Plant Physiol., 61: 768-775.
CrossRefDirect Link - Abdel Latef, A.A.H., A.K. Srivastava, M.S.A. El‐Sadek, M. Kordrostami and L.S.P. Tran, 2018. Titanium dioxide nanoparticles improve growth and enhance tolerance of broad bean plants under saline soil conditions. Land Degrad. Dev., 29: 1065-1073.
CrossRefDirect Link - Khan, M.N., 2016. Nano-titanium dioxide (Nano-TiO2) mitigates NaCl stress by enhancing antioxidative enzymes and accumulation of compatible solutes in tomato (Lycopersicon esculentum Mill.). J. Plant Sci., 11: 1-11.
CrossRefDirect Link - Ma, D., H. Ding, C. Wang, H. Qin and Q. Han et al., 2016. Alleviation of drought stress by hydrogen sulfide is partially related to the abscisic acid signaling pathway in wheat. Plos One, Vol. 11.
CrossRefDirect Link - Shan, C., S. Zhang and X. Ou, 2018. The roles of H2S and H2O2 in regulating AsA-GSH cycle in the leaves of wheat seedlings under drought stress. Protoplasma, 255: 1257-1262.
CrossRefDirect Link - Nield, J., K. Redding and M. Hippler, 2004. Remodeling of light-harvesting protein complexes in Chlamydomonas in response to environmental changes. Eukaryotic Cell, 3: 1370-1380.
CrossRefDirect Link - Song, Y.G., B. Liu, L.F. Wang, M.H. Li and Y. Liu, 2006. Damage to the oxygen-evolving complex by superoxide anion, hydrogen peroxide and hydroxyl radical in photoinhibition of photosystem II. Photosynth. Res., 90: 67-78.
CrossRefDirect Link - Neill, S.J., R. Desikan and J.T. Hancock, 2003. Nitric oxide signalling in plants New Phytol., 159: 11-35.
CrossRefDirect Link - Zhang, H., H. Jiao, C.X. Jiang, S.H. Wang, Z.J. Wei, J.P. Luo and R.L. Jones, 2010. Hydrogen sulfide protects soybean seedlings against drought-induced oxidative stress. Acta Physiol. Plantarum, 32: 849-857.
CrossRefDirect Link - Wei, B., W. Zhang, J. Chao, T. Zhang and T. Zhao et al., 2017. Functional analysis of the role of hydrogen sulfide in the regulation of dark-induced leaf senescence in Arabidopsis. Scient. Rep., Vol. 7.
Direct Link