
Hamim Hamim
Department of Biology, Faculty of Mathematics and Natural Sciences, Bogor Agricultural University, Bogor, Indonesia
LiveDNA: 62.15393
Violita Violita
Department of Biology, Faculty of Mathematics and Natural Sciences, State University of Padang (UNP), Padang, Indonesia
Triadiati Triadiati
Department of Biology, Faculty of Mathematics and Natural Sciences, Bogor Agricultural University, Bogor, Indonesia
Miftahudin Miftahudin
Department of Biology, Faculty of Mathematics and Natural Sciences, Bogor Agricultural University, Bogor, Indonesia
Asian Journal of Plant Sciences
Year: 2017 | Volume: 16 | Issue: 2 | Page No.: 65-77







ABSTRACT
Background and Objective: Drought is an abiotic factor that significantly reduces agriculture production almost every year. Drought stress especially during excessive light results in enhancement of Reactive Oxygen Species (ROS) leading to an occurring oxidative stress. The increase of ROS also occurs in plants applied by herbicide. In this study, oxidative stress of three soybean (Glycine max L.) Merr) cultivars and a wild line soybean (G. tomentella ) were analyzed in response to drought and paraquat treatments. Methodology: Drought treatment was performed by withholding water for 12 days (for cultivars) and 22 days (for wild line soybean) in greenhouse experiment during flower initiation. Paraquat treatment was applied using manual sprayer at the same time of drought treatment application. Plant water status and photosynthetic rate were measured during the drought treatment and after rewatering and after paraquat application. During the treatment, malondialdehyde (MDA) and the activity of Glutathione Reductase (GR) and superoxide dismutase (SOD) enzymes were measured. Results: Drought treatment decreased plant relative water content up to 33 and 42% in sensitive and tolerant variety respectively. Transpiration and photosynthetic rate decreased almost to zero at the end of drought period, while those of control plant were 4.7 and 12.58 μmol m–2 sec–1, respectively. Malondialdehyde content and antioxidative enzymes GR and SOD increased substantially during the drought and paraquat application in all cultivated varieties as well as wild soybean. Conclusion: Drought and paraquat application induced oxidative stress in soybean cultivars as well as in wild soybean indicated by dramatic rising of ROS and the increase of malondialdehyde and antioxidative enzyme (GR and SOD) by approximately 2-3 folds but there was no clear pattern of enzyme activities between tolerant and sensitive varieties.
PDF Abstract XML References Citation
Received: November 14, 2016;
Accepted: February 15, 2017;
Published: March 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Hamim Hamim, Violita Violita, Triadiati Triadiati and Miftahudin Miftahudin, 2017. Oxidative Stress and Photosynthesis Reduction of Cultivated
(Glycine max L.) and Wild Soybean (G. tomentella L.) Exposed to
Drought and Paraquat. Asian Journal of Plant Sciences, 16: 65-77.
DOI: 10.3923/ajps.2017.65.77
URL: https://scialert.net/abstract/?doi=ajps.2017.65.77
DOI: 10.3923/ajps.2017.65.77
URL: https://scialert.net/abstract/?doi=ajps.2017.65.77
INTRODUCTION
Drought is an abiotic factor that significantly reduces agriculture production almost every year. Drought stress causes the plant photosynthesis rate reduces by decreasing stomatal conductance1-4 and may also cause the damage of the photosynthesis apparatus including photosystem I (PSI) and photosystem II (PSII) of photosynthesis5 and loss of ATP synthase6. In addition, partial stomatal closure due to drought stress in C3 plants increases photorespiration7,8 that causes the decrease of carbohydrate accumulation required for growth and seed filling.
The lower CO2 assimilation rate caused by drought, especially during excessive light exposure, may lead to over-reduction of PSII photosynthesis reaction center9. This situation may result in enhancement of Reactive Oxygen Species (ROS) such as superoxide radical ions (O2‾), hydrogen peroxides (H2O2) and oxygen singlets10-15. Reactive oxygen species are unstable substances that are harmful to the cells or tissue and a higher degree of ROS accumulation can cause cell and tissue damage16,17 leading to oxidative stress.
A growing body of evidence indicates that drought stress increases the accumulation of ROS in many species including wheat (Triticum aestivum L.)18, rice19, maize20, sunflower (Helianthus annuus L.)16, Cleome spinosa21 and some perennial plants such as Coffea canephora22 and rubber tree, Hevea brasiliensis23. In soybean (Glycine max L.), the high degree of nodule senescence in response to drought stress is also predicted due to high accumulation of ROS during drought stress24. Water stress-induced abscisic acid (ABA) accumulation triggers the increased generation of ROS, which in turn, leads to the up-regulation of the antioxidant defense system25.
The increase of ROS also occurs in plants applied by herbicide such as in pea (Pisum sativum)26, grapevine27, winter wheat, rye and maize28, tobacco29 and crickweed (Malachium aquaticum)30 indicated by the increase of malondialdehyde or other radical compounds such as superoxide28 anion O2∙‾. This phenomenon is quite stimulating to observe further the magnitude and pattern of ROS accumulation induced by water stress as compared to that produced by herbicide treatment.
It is still unclear whether the accumulation of ROS and the elimination of this substance by antioxidative enzymes activities are different in tolerant and sensitive plants. Iturbe-Ormaetxe et al.31 have concluded that tolerance to water deficit in terms of oxidative damage largely depends on the cultivar, however, little data exists indicating the differences in antioxidative enzyme activities of tolerant and sensitive plants. Some experiments in soybean have shown that the accumulation of enzymes such as superoxide dismutase (SOD) and ascorbate peroxidase (APX) were lower in drought stressed nodules of mycorrhizal plants than in non-mycorrhizal plants, whereas Glutathione Reductase (GR) activity was higher in nodules from mycorrhizal plants than in non-mycorrhizal plants24.
In this study the analysis of photosynthetic rate, ROS accumulation and antoxidative enzyme activities of tolerant and sensitive soybean varieties (G. max L.) and wild soybean (G. tomentella L.) was carried out in response to drought stress and paraquat, a herbicide that is able to induce oxidative stress31 by accumulation of high ROS in the plants.
MATERIALS AND METHODS
The experiment was carried out in a greenhouse at Department of Biology, Faculty of Mathematics and Natural Sciences, Bogor Agricultural University, Bogor, Indonesia. The plants used in this experiment were drought-tolerant, moderately tolerant and sensitive soybean varieties (Glycine max L.) and a wild soybean (Glycine tomentella L.). A drought-tolerant variety (Tidar), a moderately tolerant variety (Burangrang) and a drought-sensitive variety (Panderman) were provided by the Indonesian Beans and Tubers Research Bureau (BALITKABI), Malang, East Java, Indonesia, while a drought-tolerant wild soybean was provided by the Life Science and Biotechnology Research Center, Bogor Agricultural University, Bogor, Indonesia.
Plant growing and treatment experiment: The plants were grown in 8 kg capacity polybags containing a mixture of soil and compost 2:1 (v/v). The plants were fertilized with nitrogen, phosphorus and potassium fertilizer (15:15:15) with the dosage of 2 g per polybag. Four seeds were sown in each polybag and then 1 week old seedlings were thinned and selected to become 2 seedlings per polybag. The experiment carried out using a completely randomized design with two factors. The first factor was plant varieties and species comprised of drought-tolerant [(1) Tidar], moderately tolerant [(2) Burangrang] and drought-sensitive varieties [(3) Panderman] and a drought-tolerant wild soybean [(4) Glycine tomentella L.]. The second factor was environmental stress including (1) Drought stress, (2) Paraquat application and (3) Normal watering (of control plants). The stress treatment (drought and paraquat application) was applied to 25 days old plants, when the plants started to flower. Drought stress was administered by withholding water for 12 days until the plants were heavily wilted. After the drought stress period, the plants were rewatered to recovery (to the condition of the control plants). Paraquat application was performed by spraying paraquat on the leaves in the morning between 07:00-08:00 am with a dosage of 90 g ha–1 of active compound. Based on a previous experiment, this dosage did not kill the plants but it reduced by 35% the biomass dry weight after 2 weeks of treatment.
Parameters measurement and photosynthetic analysis: The parameters measured in this experiment were media water content, relative water content of plants, transpiration and photosynthetic rate, antioxidant enzymes activity and lipid peroxidation activity. Gas exchange analysis was carried out to analyze the transpiration and photosynthetic rate of the youngest fully expanded leaves using photosynthetic leaf chamber analyzer type of LCA-4 with the light intensity of approximately 950 μmol m–2 sec–1. The photosynthetic rate was measured at 0, 4, 8, 10 and 12 days after drought stress treatment for cultivated soybean and at 0. 4, 10, 18 and 22 days after drought stress treatment for wild soybean. The measurement was again carried out 2 days after rewatering to analyze photosynthetic rate of recovered plants. For the plants treated with paraquat herbicide, the photosynthetic rate measurement was carried out before paraquat application, 4 h after application and 1, 3 and 5 days after application.
Enzymes analysis: For the analysis of antioxidant enzymes, 0.2 g of the youngest fully expanded leaf samples were ground in 4 mL solute containing 50 mM phosphate buffer (pH 7.0), 1% polyvinylpolypyrrolidone and 0.2 mM ascorbic acid, which were placed in an ice bath. The homogenate was centrifuged at 1000 rpm for 30 min at 40°C. The supernatant was used for assay of enzymes activity using method of Jiang and Huang32 with some modification. The activity of GR was determined by following the decreased of absorbance at 340 nm for 1 min due to the glutathione dependence of NADPH33. The reaction mixture contained 1 mM of ethylene diamine tetra acetic acid (EDTA), 0.5 mM of glutathione disulfide (GSSG), 0.15 mM of reduced nicotinamide adenine dinucleotide phosphate (NADPH), 100 mM of phosphate buffer (pH 7.8) and 0.5 mL of extract enzyme. Glutathione reductase activity was determined per weigh of unit protein, while the protein content was determined by Bradford method34 using bovine serum albumin as a standard. The activity of SOD enzyme was determined using method described by Beauchamp and Fridovich35 with some modification. The measurement carried out based on the inhibition of blue diformazan with the existence of riboflavin/nitro blue tetrazolium (NBT) and light. Leaf extract (30 μL) was added to the medium containing 50 mM phosphate buffer (pH 7.8), 0.1 mM of EDTA and 0.3 mM of riboflavin. After 5 min incubation at the room temperature, the extract was added by 0.03 mM NBT and placed to an approximately 20 cm under 75 W lamp for 1 min. The initial rate of the reaction was determined by the increase of absorbance for every 30 sec at 560 nm. The solution without sample was used as control. The activity of SOD was determined as U mg–1 of protein, 1 U was 50% form inhibition of blue diformazan.
Lipid peroxidation analysis: Lipid peroxidation was estimated as the content of total 2-thiobarbituric acid-(TBA) substances expressed as equivalent to malondialdehyde (MDA) production as described in Ono et al.36 with some modifications. Fresh leaves (0.2 g) were extracted in 0.5 mL of 0.1% (w/v) trichloracetic acid (TCA) at 4°C. The extract then was added to 3 mL of 1% H3PO4 and 1 mL of 0.6% TBA dissolved in 20% TCA. The solution was incubated in the oven at a temperature of 100°C for 30 min. After cooling to the room temperature, 4 mL n-butanol was added to the solution and followed by centrifugation at 4200 rpm at 28°C for 20 min. The absorbance of supernatant was then measured using a UV-Vis spectrophotometer at 532 nm and corrected for nonspecific turbidity by subtracting the absorbance at 520 nm. The concentration of MDA was calculated from its extinction coefficient (ε = 155 L mmol–1 cm–1).
Statistical data analysis: The collected data were analyzed statistically using SPSS 16 software to identify significant differences among the treatments and t-student test was applied to compare among the means.
RESULTS AND DISCUSSION
Plant water status during the drought and paraquat treatment: To analyze plant water status, Relative Water Content (RWC) of plant was measured periodically during the drought and paraquat treatments. The average of RWC of the control plants was 80.3% with small variation between 78.0-82.8% (Table 1). The drought stress treatment for 12 days significantly decreased RWC of the cultivated plant. At the last day of the drought treatment, the sensitive soybean (Panderman) had lower RWC (33%) than the tolerant (Tidar) and moderate (Burangrang) varieties (42.5 and 42.0% respectively). The most dramatic reduction of RWC occurred after 8 days of drought treatment for cultivated soybean, whereas in wild soybean G. tomentella it occurred after 12 days of drought stress (Table 1).
| Table 1: | Relative water content (%) of soybean leaves of Tidar, Burangrang, Panderman and G. tomentella in response to drought period (days) |
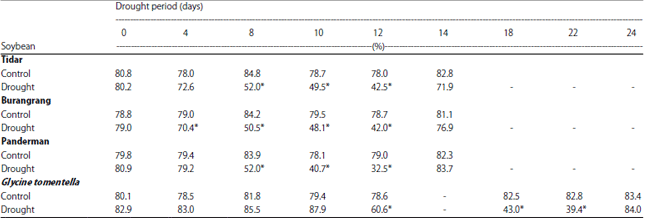 | |
| Values in the same column significantly different at 5% of t-student | |
Glycine tomentella survived 22 days of drought period with the RWC of 39.4%. Two days after rewatering the RWC of all the plants rose again to that of the control plants.
The RWC of plants treated with paraquat was also reduced significantly 1 day after treatment. The RWC started to decline 4 h after paraquat application and the maximum reduction was observed 1 day after treatment. Three days after paraquat application, the RWC rose again for tolerant, moderate and wild soybeans, while it remained low for the sensitive variety (Table 2). Even though the RWC of wild soybean G. tomentella dropped dramatically one day after paraquat application, it recovered very well 3 days after application. The RWC of the paraquat-treated sensitive variety recovered 5 days after the treatment (Table 2).
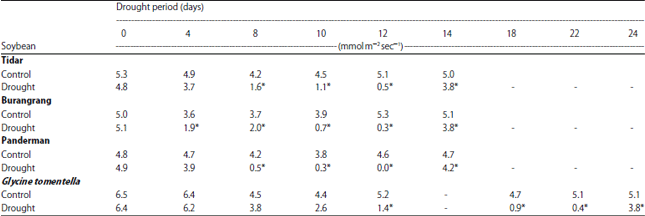
Gas exchange analysis: Transpiration rate (E) varied during the treatment with the average of 4.7 mmol m–2 sec–1 for control plants. During the drought stress period, the E decreased significantly and dropped almost to zero after 12 days in cultivated soybeans (Tidar, Burangrang and Panderman), whereas in wild soybean (G. tomentella) the maximum reduction of E occurred 22 days after the drought stress period. Three days after rewatering, the E increased again to the level of control plants (Table 3). Application of paraquat also reduced E and the reduction became significantly different from that of control plants 3 days after application.
The photosynthesis rate (Pn) measured under green house conditions with average PPFD of 950 mmol m–2 fluctuated during the day. The average Pn of control plants and both cultivated and wild soybean, measured between 08:00-10:00 am was 12.6 mmol m–2 sec–1. As the drought stress period increased, the Pn reduced gradually with the maximum reduction on 12 and 22 days after drought stress treatment for cultivated and wild soybeans, respectively (Fig. 1).
| Table 2: | Relative water content (%) of soybean leaves of Tidar, Burangrang, Panderman and G. tomentella after Paraquat application and control plants |
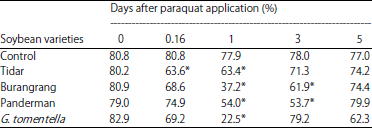 | |
| Values in the same column significantly different at 5% of t-student | |
The Pn of the drought-sensitive soybean (Panderman) decreased more than that of the tolerant (Tidar) and moderate (Burangrang) varieties after 8 and 10 days drought stress treatment (Fig. 1). At the period of maximum drought stress, the Pn of all plants dropped to near zero. Meanwhile, rewatering increased the Pn back to the normal (control) condition 2 days after rewatering (Fig. 1).
Paraquat treatment also caused significant reduction of the Pn of all treated plants. Even though Pn reduction was detected 4 h after paraquat application, significant reduction of Pn occurred 1 day after application (Fig. 2). The reduction of Pn continued until it reached the minimum level (approximately 0.14 mmol m–2 sec–1) on the 3rd day after application. The Pn increased again 5 days after the plants recovered from stress. In contrast to the Pn recovery after drought stress, the recovery of Pn 5 days after paraquat application did not return back to the level of the control plants (Fig. 2).
Lipid peroxidation: Lipid peroxidation was analyzed by the measurement of MDA accumulation in the leaf tissues as a result of membrane lipid degradation.
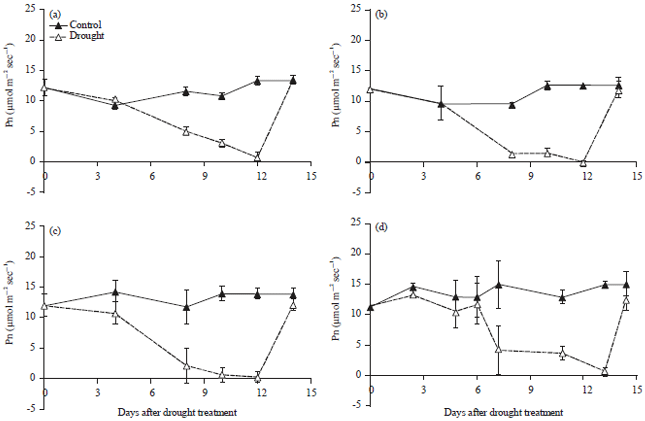 | |
| Fig. 1(a-d): | Photosynthetic rate (Pn) of cultivated soybean varieties (a) Tidar, (b) Burangrang, (c) Panderman and (d) Wild soybean Glycine tomentella during drought period (Mean+SE of t-student test at α of 5%, n = 3) |
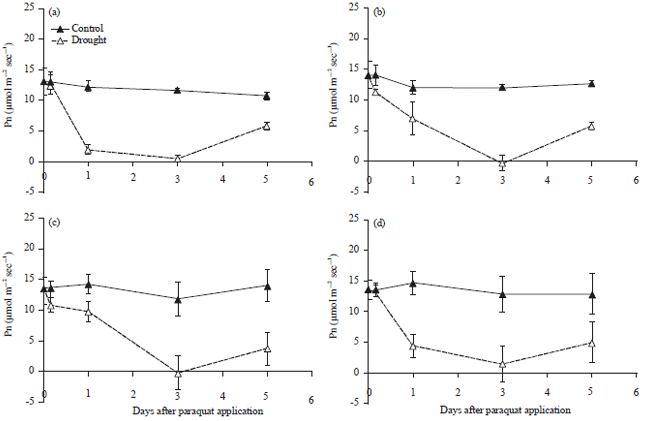 | |
| Fig. 2(a-d): | Photosynthetic rate (Pn) of cultivated soybean varieties (a) Tidar, (b) Burangrang, (c) Panderman and (d) Wild soybean Glycine tomentella during 5 days after paraquat application (Mean+SE of t-student test at α of 5%, n = 3) |
At normal conditions indicated by the control plants, the average MDA level in the leaves was 3281 nmol g–1 fresh weight. The MDA level increased in drought-stressed plants (Fig. 3). Generally, the MDA level of cultivated soybean significantly increased after 8 days of drought treatment when the plant started wilting. However, in G. tomentella MDA levels started to increase when mild drought stress was reached (4 days after drought treatment). The maximum level of MDA (5890 nmol g–1 fresh weight) or almost twice that of control plants was reached at 10 days when severe drought stress occurred. After rewatering, the MDA level decreased to that of the control plants or even lower in sensitive variety Panderman (Fig. 3).
Application with paraquat herbicide also increased MDA level in all treated plants 1 day after treatment (Fig. 4). However, the maximum level of MDA was lower in paraquat application compared to that of drought treatment.
| Table 3: | Transpiration rate (mmol m–2 sec–1) of Tidar, Burangrang, Panderman and G. tomentella in response to drought period (days) |
 | |
| Values in the same column significantly different at 5% of t-student | |
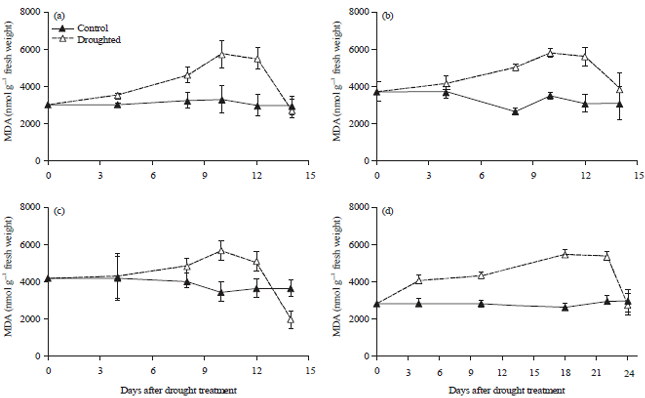 | |
| Fig. 3(a-d): | Malondialdehyde (MDA) content of soybean leaves of (a) Tidar, (b) Burangrang, (c) Panderman and (d) Wild soybean Glycine tomentella during drought period (Mean+SE of t-student test at α of 5%, n = 3) |
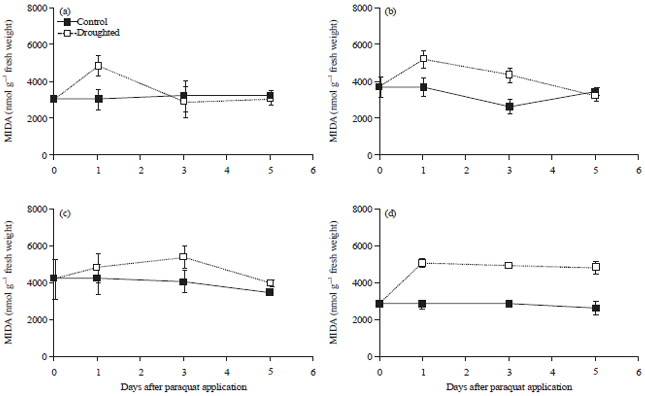 | |
| Fig. 4(a-d): | Malondialdehyde (MDA) content of soybean leaves of (a) Tidar, (b) Burangrang, (c) Panderman and (d) Wild soybean Glycine tomentella 5 days after paraquat application (Mean+SE of t-student test at α of 5%, n = 3) |
Five days after paraquat application, the MDA concentration decreased again to the level of that in the control plants, except in G. tomentella, which remained unchanged (Fig. 4).
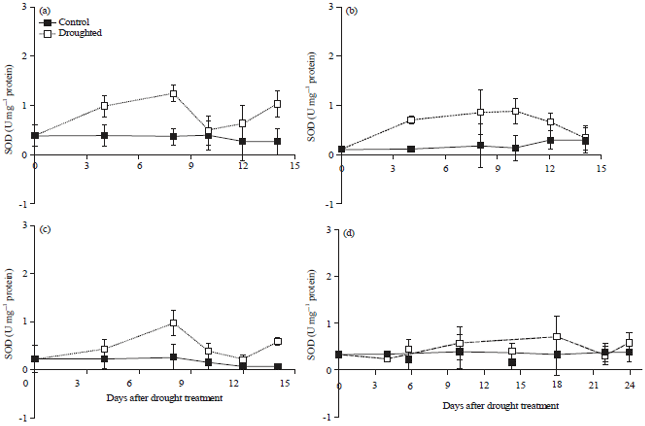
Antioxidative enzyme activity: Drought stress caused an increase of antioxidative enzymes (GR and SOD) activity in all soybean starting from mild drought. The highest activity of the enzymes was recorded after 8 days of the drought for cultivated soybean and the value decreased again at the end of drought period (12 days). There was no different pattern of enzyme activities between tolerant and sensitive soybean varieties (Fig. 5, 6). In G. tomentella there was different pattern of GR and SOD enzyme activities, where the activity of GR enzyme increased gradually and reached the maximum level at the end of drought period, while it was not occurred in SOD enzyme (Fig. 5, 6). Only wild soybean that had the highest GR activity during the last period of drought.
The response of antioxidative enzyme activities (GR and SOD) to paraquat treatment was different between GR and SOD. The GR activities tended to increase linearly in response to paraquat treatment starting from the beginning of treatment until 5 days after herbicide application. On the other hand, SOD enzyme was highly active 1 day after herbicide application and then the activity tended to decrease slowly until 5 days after application. There was almost similar pattern of SOD activity in response to paraquat application for all soybean varieties as well as wild soybean (Fig. 7, 8).
Water balance and gas exchange inside the plant during drought stress: Water deficit in plants occurs when water loss due to the transpiration of the leaves exceeds water absorption by the plant roots37. When this happens, the plant generally tends to reduce water loss by reducing transpiration rate as indicated by Table 3. Lower transpiration in response to drought stress is associated with the decrease of stomatal conductance which is sensitive to water deficit even before the water potential of the plant decreases38,39. The different responses of E and Pn to drought stress treatment between cultivated soybeans and wild soybean (G. tomentella), where those parameters dramatically reduced after 12 days in cultivated soybeans varieties, while in G. tomentella the reduction occurred after 22 days after drought period may be explained by differences in growth and canopy development. The cultivated soybeans Tidar, Burangrang and Panderman had similar growth and canopy development, while wild soybean G. tomentella grew more slowly with smaller leaves than cultivated plants. Therefore, the water was lost from the media through transpiration and consequently caused reduction of RWC of cultivated soybeans faster than wild one (G. tomentella).
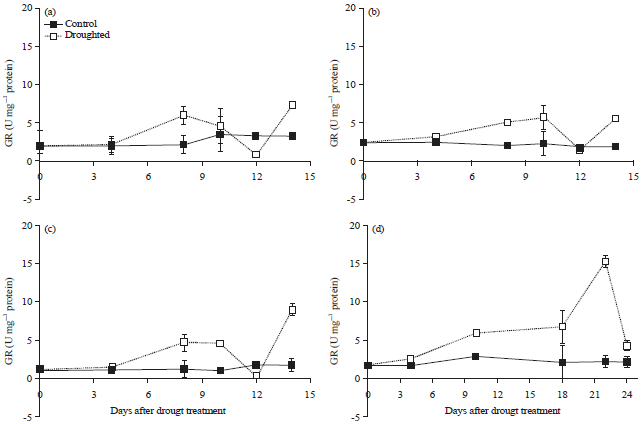 | |
| Fig. 5(a-d): | Glutathione Reductase Enzyme (GR) of soybean leaves of (a) Tidar, (b) Burangrang, (c) Panderman and (d) Wild soybean G. tomentella during drought period (Mean+SE of t-student test at α of 5%, n = 3) |
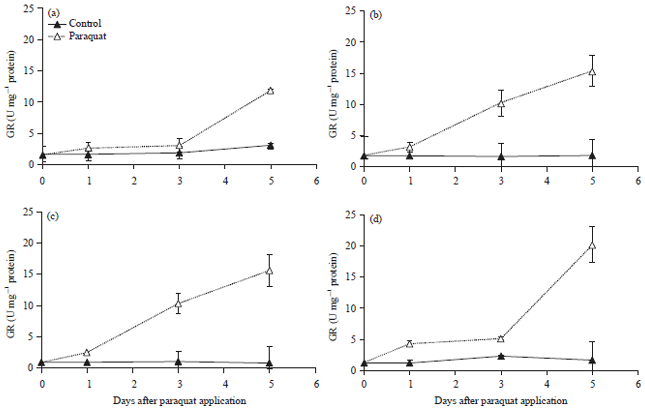 | |
| Fig. 6(a-d): | Glutathione Reductase Activity (GR) of of soybean leaves of (a) Tidar, (b) Burangrang, (c) Panderman and (d) Wild soybean G. tomentella 5 days after paraquat application (Mean+SE of t-student test at α of 5%, n = 3) |
 | |
| Fig. 7(a-d): | Superoxide dismutase (SOD) activity of soybean leaves of (a) Tidar, (b) Burangrang, (c) Panderman and (d) Wild soybean G. tomentella during drought period (Mean+SE of t-student test at α of 5%, n = 3) |
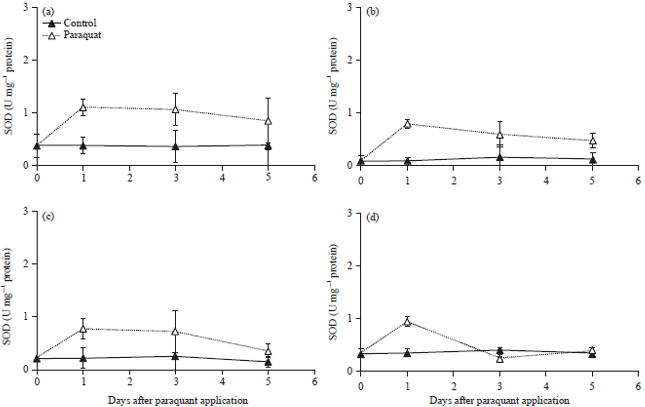 | |
| Fig. 8(a-d): | Superoxide dismutase (SOD) activity of a soybean leaves of (a) Tidar, (b) Burangrang, (c) Panderman and (d) Wild soybean G. tomentella 5 days after paraquat application (Mean+SE of t-student test at α of 5%, n = 3) |
The reduction of transpiration rate as a response to drought stress was in accordance with the decrease of photosynthetic rate that dramatically declined after 8 days of drought (Table 3, Fig. 1). The decrease of stomatal conductance reduced CO2 supply to the chloroplast and then reduced photosynthetic rate37. Stomatal conductance is the main factor that influences photosynthesis reduction in plants exposed to drought stress2.
The application of paraquat herbicide also reduced transpiration as well as photosynthetic rate, however, the reduction of the Pn was faster than the reduction of E (Fig. 2), even though the maximum reduction of Pn occurred as the same time as that of E (3 days after paraquat application). Paraquat is an active compound that accepts electrons from the early acceptors of photosystem I and then reacts with oxygen to form superoxide, a free radical40. High accumulation of ROS inside the leaf may cause damage to chloroplast components, especially lipids, which consequently can reduce photosynthetic rate. This herbicide is also known to induce oxidative stress specified by the increase of hydrogen peroxide and malondialdehyde inside the cell and induce ultrastructural changes41,42. The evidence of chlorophyll content reduction in Amaranthus caudatus, Celocia argentea and Corchorus olitorius due to paraquat exposure has been obtained by Akinloye et al.43.
The increase of E and Pn back to the level of control plants 2 days after drought-stressed plants were rewatered, indicated that all the plants were capable of recovery after drought (Fig. 2). On the other hand, the E and Pn of paraquat treated plants did not recover well until 5 days after paraquat application (Fig. 2). This treatment may cause damage to photosynthetic and other cellular apparatus, causing malfunction of these components and the need to be rebuilt by the growth of new shoots. Paraquat is a compound that can induce the accumulation of free radicals that cause cellular damage44,45.
Oxidative stress induced by severe drought stress as well as paraquat: The increase of MDA in all of the soybean plants during severe drought stress indicated that severe drought stress can induce oxidative stress in both cultivated and wild soybean plants. The MDA increased dramatically 8 days after the induction of drought stress and reached the maximum level on the 10th and 12th days of drought when the stress was most severe (Fig. 3). The MDA is a compound resulting from lipid peroxidation at the cellular level and is frequently used as an indicator of lipid peroxidation level due to oxidative stress31.
The increase of MDA also occurred in the plants treated by paraquat herbicide (Fig. 4). This was also in agreement to the result presented by Akinloye et al.43 that the increase of MDA was associated to the level of paraquat treatments in A. caudatus, C. argentea and C. olitorius. This herbicide is a compound that can be reduced by photosynthetic reaction to become an unstable free radical. This radical compound can be oxidized back by oxygen to form the original ions and hydrogen peroxide (H2O2), which significantly inhibits photosynthesis especially photosystem I46 and also destroys cells and tissues44. When this happens, the plant undergoes lipid peroxidation, protein degradation, DNA denaturation and pigment damage45. At the cellular level, it causes damage to the cellular membrane and chloroplast44. The fact that the increase of MDA levels after 10 days of drought stress was approximately the same as that of the paraquat application, indicated that oxidative stress may be induced by severe drought. The level of MDA decreased to that of the control plant 2 days after the drought-stressed plants were rewatered. The same result has also been demonstrated by Zhang and Kirkham47 on wheat and Wang and Huang48 on bluegrass. This seems to indicate that the plants had recovered from oxidative stress after 2 days of rewatering.
The increase of oxidative stress due to drought stress was also confirmed by the dramatic escalation of antioxidative enzymes GR and SOD in all soybean by approximately 2-3 times in response to drought as well as to paraquat application (Fig. 5-8). The SOD is widely known as antioxidative enzyme in many species in response to environmental stress especially drought stress such as in oilseed rape49, chickpea50, rice51 and wheat52. Drought stress promotes oxidative stress due to over-generation of ROS and the modulation of the antioxidant defense system is one of the important strategies responsible for drought resistance52. Antioxidative enzymes have been confirmed to become an efficient scavenger of ROS such as H2O2 produced during drought, which may result in better protection especially for tolerant plant51.
CONCLUSION
Drought stress caused gradual decrease of relative water content and photosynthesis of three cultivars and one wild line of soybean. Photosynthesis reduction was also occurred due to paraquat treatment with more rapid response, even though there was no correlation with the decrease of water status. The ROS was dramatically accumulated in response to drought as well as paraquat application showed by the increase of malondialdehyde and antioxidative enzyme GR and SOD by approximately 2-3 folds during severe drought and 1 day after paraquat application suggesting that plants underwent oxidative stress due to severe drought stress. There was no particular distinct level of GR and SOD among tolerant and sensitive cultivars during the period of drought but GR level was almost 3 times higher in wild soybean.
SIGNIFICANCE STATEMENT
| • | Drought stress and paraquat caused Reactive Oxygen Species (ROS) development in soybean cultivar (Glycine max L.) and wild soybean (Glycine tomentella L.) |
| • | Photosynthesis (Pn) decreased dramatically due to severe drought but recovered completely 2 days after rewatering |
| • | Paraquat application also decreased Pn soon after application but did not recovered completely after several days |
| • | Drought and paraquat induced of antioxidative enzyme activity (GR and SOD) dramatically in cultivar as well as wild soybean |
ACKNOWLEDGMENTS
This study have been funded by the Directorate General of Higher Education, the Ministry of National Education, Republic of Indonesia through the Program of Competitional Grant Research for Higher Education Institution in Indonesia.
REFERENCES
- Farooq, M., A. Wahid, N. Kobayashi, D. Fujita and S.M.A. Basra, 2009. Plant drought stress: Effects, mechanisms and management. Agron. Sustainable Dev., 29: 185-212.
CrossRefDirect Link - Cornic, G., 2000. Drought stress inhibits photosynthesis by decreasing stomatal aperture-not by affecting ATP synthesis. Trends. Plant Sci., 5: 187-188.
CrossRefDirect Link - Hamim, 2005. Photosynthesis of C3 and C4 species in response to increased CO2 concentration and drought stress. Hayati J. Biosci., 12: 131-138.
Direct Link - Hamim, S. Banon and D. Dorly, 2016. Comparison of physiological and anatomical changes of C3 (Oryza sativa [L.]) and C4 (Echinochloa crusgalli [L.]) leaves in response to drought stress. IOP Conf. Ser.: Earth Environ. Sci., Vol. 31.
CrossRefDirect Link - Genty, B., J.M. Briantais and J.B.V. da Silva, 1987. Effects of drought on primary photosynthetic processes of cotton leaves. Plant Physiol., 83: 360-364.
CrossRefDirect Link - Chastain, D.R., J.L. Snider, G.D. Collins, C.D. Perry, J. Whitaker and S.A. Byrd, 2014. Water deficit in field-grown Gossypium hirsutum primarily limits net photosynthesis by decreasing stomatal conductance, increasing photorespiration and increasing the ratio of dark respiration to gross photosynthesis. J. Plant Physiol., 171: 1576-1585.
CrossRefDirect Link - Smirnoff, N., 1993. The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol., 125: 27-58.
CrossRefDirect Link - Bartoli, C.G., M. Simontacchi, E. Tambussi, J. Beltrano, E. Montaldi and S. Puntarulo, 1999. Drought and watering-dependent oxidative stress: Effect on antioxidant content in Triticum aestivum L. leaves. J. Exp. Bot., 50: 375-383.
CrossRefDirect Link - Loggini, B., A. Scartazza, E. Brugnoli and F. Navari-Izzo, 1999. Antioxidative defense system, pigment composition, and photosynthetic efficiency in two wheat cultivars subjected to drought. Plant Physiol., 119: 1091-1099.
CrossRefDirect Link - Noctor, G. and C.H. Foyer, 1998. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Mol. Biol., 49: 249-279.
CrossRefPubMedDirect Link - Noctor, G., S. Veljovic-Jovanovic, S. Driscoll, L. Novitskaya and C.H. Foyer, 2002. Drought and oxidative load in the leaves of C3 plants: A predominant role for photorespiration? Ann. Bot., 89: 841-850.
CrossRefDirect Link - Hossain, M.A., S. Bhattacharjee, S.M. Armin, P. Qian and W. Xin et al., 2015. Hydrogen peroxide priming modulates abiotic oxidative stress tolerance: Insights from ROS detoxification and scavenging. Front. Plant Sci., Vol. 6.
CrossRefDirect Link - Sgherri, C.L.M. and F. Navari-Izzo, 1995. Sunflower seedlings subjected to increasing water deficit stress: Oxidative stress and defence mechanisms. Physiologia Plantarum, 93: 25-30.
CrossRefDirect Link - Scandalios, J.G., 1997. Oxidative stress and defense mechanisms in plants: Introduction. Free Radic. Biol. Med., 23: 471-472.
PubMed - Sairam, R.K., P.S. Deshmukh and D.C. Saxena, 1998. Role of antioxidant systems in wheat genotypes tolerance to water stress. Biologia Plantarum, 41: 387-394.
CrossRefDirect Link - Qian, B., X. Li, X. Liu, P. Chen, C. Ren and C. Dai, 2015. Enhanced drought tolerance in transgenic rice over-expressing of maize C4 phosphoenolpyruvate carboxylase gene via NO and Ca2+. J. Plant Physiol., 175: 9-20.
CrossRefDirect Link - Talaat, N.B., B.T. Shawky and A.S. Ibrahim, 2015. Alleviation of drought-induced oxidative stress in maize (Zea mays L.) plants by dual application of 24-epibrassinolide and spermine. Environ. Exp. Bot., 113: 47-58.
CrossRefDirect Link - Uzilday, B., I. Turkan, A.H. Sekmen, R. Ozgur and H.C. Karakaya, 2012. Comparison of ROS formation and antioxidant enzymes in Cleome gynandra (C4) and Cleome spinosa (C3) under drought stress. Plant Sci., 182: 59-70.
CrossRefDirect Link - Lima, A.L.S., F.M. DaMatta, H.A. Pinheiro, M.R. Totola and M.E. Loureiro, 2002. Photochemical responses and oxidative stress in two clones of Coffea canephora under water deficit conditions. Environ. Exp. Bot., 3: 239-247.
CrossRefDirect Link - Wang, L.F., 2014. Physiological and molecular responses to drought stress in rubber tree (Hevea brasiliensis Muell. Arg.). Plant Physiol. Biochem., 83: 243-249.
CrossRefDirect Link - Porcel, R., J.M. Barea and J.M. Ruiz-Lozano, 2003. Antioxidant activities in mycorrhizal soybean plants under drought stress and their possible relationship to the process of nodule senescence. New Phytol., 157: 135-143.
CrossRefDirect Link - Jiang, M. and J. Zhang, 2002. Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J. Exp. Bot., 53: 2401-2410.
CrossRefDirect Link - Doulis, A.G., J.L. Donahue and R.G. Alscher, 1998. Differential reponses to paraquat-induced oxidative injury in a pea (Pisum sativum) protoplast system. Physiologia Plantarum, 102: 461-471.
CrossRefDirect Link - Tan, W., Q. Li and H. Zhai, 2012. Photosynthesis and growth responses of grapevine to acetochlor and fluoroglycofen. Pestic. Biochem. Physiol., 103: 210-218.
CrossRefDirect Link - Lukatkin, A.S., A.N. Gar'kova A.S. Bochkarjova, O.V. Nushtaeva and J.A.T. da Silva, 2013. Treatment with the herbicide TOPIK induces oxidative stress in cereal leaves. Pestic. Biochem. Physiol., 105: 44-49.
CrossRefDirect Link - Darwish, M., V. Vidal, F. Lopez-Lauri, O. Alnaser, S. Junglee, M. El Maataoui and H. Sallanon, 2015. Tolerance to clomazone herbicide is linked to the state of LHC, PQ-pool and ROS detoxification in tobacco (Nicotiana tabacum L.). J. Plant Physiol., 175: 122-130.
CrossRefDirect Link - Xu, L., W. Zhang, B. Ali, F. Islam, J. Zhu and W. Zhou, 2015. Synergism of herbicide toxicity by 5-aminolevulinic acid is related to physiological and ultra-structural disorders in crickweed (Malachium aquaticum L.). Pestic. Biochem. Physiol., 125: 53-61.
CrossRefDirect Link - Iturbe-Ormaetxe, I., P.R. Escuredo, C. Arrese-Igor and M. Becana, 1998. Oxidative damage in pea plants exposed to water deficit or paraquat. Plant Physiol., 116: 173-181.
CrossRefDirect Link - Jiang, Y. and B. Huang, 2001. Drought and heat stress injury to two cool-season turfgrasses in relation to antioxidant metabolism and lipid peroxidation. Crop Sci., 41: 436-442.
CrossRefDirect Link - Cakmak, I., D. Strbac and H. Marschner 1993. Activities of hydrogen peroxide scavenging enzymes in germinating wheat seeds. J. Exp. Bot., 44: 127-132.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Beauchamp, C. and I. Fridovich, 1971. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem., 44: 276-287.
CrossRefPubMedDirect Link - Ono, K., Y. Yamamoto, A. Hachiya and H. Matsumoto, 1995. Synergistic inhibition of growth by aluminum and iron of tobacco (Nicotiana tabacum L.) cells in suspension culture. Plant Cell Physiol., 36: 115-125.
CrossRefDirect Link - Davies, W.J. and J. Zhang, 1991. Root signals and the regulation of growth and development of plants in drying soil. Annu. Rev. Plant Physiol. Plant Mol. Biol., 42: 55-76.
CrossRefDirect Link - Davies, W.J., F. Tardieu and C.L. Trejo, 1994. How do chemical signals work in plants that grow in drying soil? Plant Physiol., 104: 309-314.
Direct Link - Chagas, R.M., J.A.G. Silveira, R.V. Ribeiro, V.A. Vitorello and H. Carrer, 2008. Photochemical damage and comparative performance of superoxide dismutase and ascorbate peroxidase in sugarcane leaves exposed to paraquat-induced oxidative stress. Pestic. Biochem. Physiol., 90: 181-188.
CrossRefDirect Link - Yoon, J.Y., J.S. Shin, D.Y. Shin, K.H. Hyun, N.R. Burgos, S. Lee and Y.I. Kuk, 2011. Tolerance to paraquat-mediated oxidative and environmental stresses in squash (Cucurbita spp.) leaves of various ages. Pestic. Biochem. Physiol., 99: 65-76.
CrossRefDirect Link - Akinloye, O.A., I. Adamson, O. Ademuyiwa and T.A. Arowolo, 2011. Paraquat toxicity and its mode of action in some commonly consumed vegetables in Abeokuta, Nigeria. Int. J. Plant Physiol. Biochem., 3: 75-82.
Direct Link - Chia, L.S., D.G. McRae and J.E. Thompson, 1982. Light-dependence of paraquat-initiated membrane deterioration in bean plants. Evidence for the involvement of superoxide. Physiologia Plantarum, 56: 492-499.
CrossRefDirect Link - Scandalios, J.G., 1993. Oxygen stress and superoxide dismutases. Plant Physiol., 101: 7-12.
CrossRefPubMedDirect Link - Dayan, F.E. and M.L.D.M. Zaccaro, 2012. Chlorophyll fluorescence as a marker for herbicide mechanisms of action. Pestic. Biochem. Physiol., 102: 189-197.
CrossRefDirect Link - Zhang, J. and M.B. Kirkham, 1994. Drought-stress-induced changes in activities of superoxide dismutase, catalase and peroxidase in wheat species. Plant Cell Physiol., 35: 785-791.
CrossRefDirect Link - Wang, Z. and B. Huang, 2004. Physiological recovery of Kentucky bluegrass from simultaneous drought and heat stress. Crop Sci., 44: 1729-1736.
CrossRefDirect Link - Abedi, T. and H. Pakniyat, 2010. Antioxidant enzyme changes in response to drought stress in ten cultivars of oilseed rape (Brassica napus L.). Czech J. Genet. Plant Breed., 46: 27-34.
Direct Link - Patel, P K., A. Hemantaranjan, B.K. Sarma and R. Singh, 2011. Growth and antioxidant system under drought stress in Chickpea (Cicer arietinum L.) as sustained by salicylic acid. J. Stress Physiol. Biochem., 7: 130-144.
Direct Link - Lum, M.S., M.M. Hanafi, Y.M. Rafii and A.S.N. Akmar, 2014. Effect of drought stress on growth, proline and antioxidant enzyme activities of upland rice. J. Anim. Plant Sci., 24: 1487-1493.
Direct Link - Farooq, M., M. Hussain and K.H.M. Siddique, 2014. Drought stress in wheat during flowering and grain-filling periods. Crit. Rev. Plant Sci., 33: 331-349.
CrossRefDirect Link