
Sadasivam Vinoth
Department of Plant Science, School of Life Sciences, Bharathidasan University, 620024 Tiruchirappalli, Tamil Nadu, India
LiveDNA: 91.13024
Sundari
Department of Plant Science, School of Life Sciences, Bharathidasan University, 620024 Tiruchirappalli, Tamil Nadu, India
Packiaraj Gurusaravanan
Department of Botany, Bharathiar University, Coimbatore, India
Subiramani Sivakumar
Department of Plant Science, School of Life Sciences, Bharathidasan University, 620024 Tiruchirappalli, Tamil Nadu, India
Govindarajan Siva
Department of Plant Science, School of Life Sciences, Bharathidasan University, 620024 Tiruchirappalli, Tamil Nadu, India
Ganesan Prem Kumar
Department of Plant Science, School of Life Sciences, Bharathidasan University, 620024 Tiruchirappalli, Tamil Nadu, India
Manju
Department of Environmental Science, PSG college of Arts and Science, Civil aerodrome post, 641014 Coimbatore, Tamil Nadu, India
Kalamani Velmurugan
Department of Biotechnology, Aarupadai Veedu Institute of Technology, Vinayaka Missions University, Paiyanoor, 603104 Chennai, India
Vemuri Lakshminarayana
Department of Biotechnology, Aarupadai Veedu Institute of Technology, Vinayaka Missions University, Paiyanoor, 603104 Chennai, India
Narayanasamy Jayabalan
Department of Plant Science, School of Life Sciences, Bharathidasan University, 620024 Tiruchirappalli, Tamil Nadu, India
Asian Journal of Plant Sciences
Year: 2017 | Volume: 16 | Issue: 4 | Page No.: 172-183







ABSTRACT
Background and Objective: Seagrass extract is a natural plant biostimulant which can be utilized for enhancing plant growth at various stages. The present study was undertaken to explore the application of Zostera marina seagrass extract as foliar spray on salt stressed tomato plants under green house conditions. Methodology: The one set of green house grown tomato plants were sprayed with seagrass extract and another set with water on 15th day of salt treatment. The accumulation of reactive oxygen species, antioxidant enzyme activities and their physiological responses towards different concentration of salt stress were examined after the 3rd treatment. Data were analyzed by one-way ANOVA using SPSS software version 17. Results: The initial studies revealed that the salt stressed plants shown variation in physiology, Relative Water Content (RWC) and accumulation of hydrogen peroxide (H2O2) in the leaves. The activities of ascorbate peroxidase (APX), superoxide dismutase (SOD), catalase (CAT) were found to have increased in seagrass extract sprayed plants when compared with the plants sprayed with water. In contrast to APX, SOD, CAT, the activities of peroxidase (POX) were declined in the seagrass treated plants. Conclusion: After 30 days, the plants sprayed with seagrass extract were more efficient in controlling the damage caused by the stress and it has shown enhanced activities of tolerance to the salt stressed plants compared to the control. Further, presence of functional groups and constituents were found in seagrass extract through FTIR and GC-MS analyses.
PDF Abstract XML References Citation
Received: April 09, 2017;
Accepted: July 03, 2017;
Published: September 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Sadasivam Vinoth, Sundari, Packiaraj Gurusaravanan, Subiramani Sivakumar, Govindarajan Siva, Ganesan Prem Kumar, Manju, Kalamani Velmurugan, Vemuri Lakshminarayana and Narayanasamy Jayabalan, 2017. Evaluation of Seagrass Liquid Extract on Salt Stress Alleviation in Tomato Plants. Asian Journal of Plant Sciences, 16: 172-183.
DOI: 10.3923/ajps.2017.172.183
URL: https://scialert.net/abstract/?doi=ajps.2017.172.183
DOI: 10.3923/ajps.2017.172.183
URL: https://scialert.net/abstract/?doi=ajps.2017.172.183
INTRODUCTION
World agriculture is facing a lot of challenges like producing 70% more food for an additional 2.3 billion people by 20501. Abiotic stresses limit the production and yield to less than 50% and among them, salinity greatly affects half of the irrigated lands worldwide2. Salinity causes membrane damage, nutrient imbalance, altered levels of growth regulators, enzymatic inhibition and metabolic dysfunction, including photosynthesis which ultimately leads to the plant death3. More than 20% of irrigated land have been damaged by salt worldwide and 1.5 M ha are taken out of production each year due to the high salinity levels in the soil4.
Tomato (Lycopersicon esculentum L.) is an economically important vegetable crop and it is grown almost in all parts of the country. Tomato contains lycopene, a valuable compound possessing anti-oxidant and anticancer properties. Hence, its production and consumption are increasing every year5. Even though tomato is well adapted to all climatic conditions, increasing soil and water salinity affects the growth, yield and fruit quality of some that leads to reduction in yield6. To overcome the damage caused by salinity stresses there are few attempts have been made by the researchers by using marine resources (seaweeds) as elicitors for the crop improvement.
Marine resources are the underutilized bioresources. The bioactive substances extracted from the marine algae and seagrass have a biostimulant activity when applied to the crops7. The seaweeds and seagrasses form an integral part of marine ecosystem and it has been used as a soil conditioner to increase the yield and productivity of the crops in an eco-friendly manner. Apart from the genetic modification system for the crop improvement, there are several reports which suggests that marine organic resources have an biostimulant activity that enhance the tolerance of crop plants to the abiotic and biotic stresses. Naturally, seaweed extracts tend to impart tolerance to plants under stress conditions by targeting number of pathways8. Application of Ascophyllum extracts exhibited significant improvement in salt and freezing tolerance of grape plants when compared to control plants9. Production of Reactive Oxygen Species (ROS) including superoxide radicals (O2‾), hydrogen peroxide (H2O2) and hydroxyl radicals (OH•) is induced in plants under several stress conditions10,11. The ROS is a product of altered chloroplast and mitochondrial metabolism during stress. Antioxidative enzymes are the most important components in the scavenging system of ROS. Under salt stress conditions, these antioxidant enzymes are increased in plant parts to remove H2O2 by converting it to water12. Superoxide dismutase (SOD) is a major scavenger of superoxide free radical anions in chloroplasts whereas the H2O2 produced is eliminated by catalase (CAT) in peroxisomes13 and a variety of peroxidases (POX) decomposes H2O2 by oxidation of co-substrates such as phenolic compounds and/or antioxidants14.
Application of an A. nodosum extract increased the salt tolerance in salt-sensitive genotype of Alfalfa15. The seaweed extracts enhanced the activities of chitinase, glucanase, PO, PAL and lipoxygenase enzymes in treated control plants16. The presence of marine bioactive substances in extract improves the stomata uptake efficiency in treated plants compared to the control plants9. There are numerous reports which suggest that seaweeds have promising role in abiotic stress tolerance of several crop and ornamental plants. But till now there are no reports on the effect of seagrass liquid extract on salt stress alleviation of tropical plants. This study will give experimental evidence to the positive role of seagrass extract in tolerance of salt stressed tomato plants. Moreover, the present study is the first report on evaluating the effect of seagrass Zostera marina extract in enhancing the tolerance of salt stressed tomato plants under green house conditions. Further significant finding of this study can be utilized to other crops as well for agricultural production.
MATERIALS AND METHODS
Collection and preparation of seagrass extract: Zostera marina samples were collected from the coastal area of Ramanathapuram district, Tamilnadu 9°16°49.58° N and 77°26°1.03° E. The plants were handpicked and washed thoroughly with seawater to remove unwanted impurities, adhering sand particles and epiphytes. They were transported to the testing laboratory at Department of Plant Science, Bharathidasan University using polythene bags. Samples were washed completely using tap water for 4-5 times and they were shade dried. After drying they were finely chopped and powdered for the preparation of extracts.
Preparation of seagrass suspension17: The seagrass suspension was prepared by boiling about 500 g of samples in 500 mL of sterile distilled water for about 20 min and finally diluted 1:3 dilution with water. Then the extracts were initially filtered using muslin cloth and then filtered with Whatman No. 41 filter study. One batch of suspension was used throughout the experiments and another set of suspension was stored at -20°C for further use.
Preparation of pots for the experiment and duration of salt stress induction: Seeds of tomato (PKM-I cultivar) were procured from Agricultural research station, Manaparai, Tiruchirappalli. The good quality seeds were selected for the experiment and it was soaked in water for 12 h. The seeds were surface sterilized using 0.1% mercuric chloride and then sown in pots filled with soil: Sand and organic manure in the ratio of 2:1:1. Approximately about 15 seeds were placed in each pot and seed to seed distance in pot was maintained as 3-5 cm. The salt stress was induced in 15 days old tomato plants with different concentration ranges from 0 (Control), 50, 100, 150 and 200 mM. The subsequent salt stress was given to the plants at 10 days interval and it was maintained in green house conditions with natural light illumination.
Experimental design and foliar spray of seagrass extracts: The experiment was divided into two treatments. In one set of experiment, the salt stressed tomato plants (15 days old) were foliar sprayed with seagrass extract. In another, the salt stressed tomato plants were foliar sprayed with distilled water. Both experiments and control plants (no salt stress induced) were maintained and the salt stress was given to plants at various concentrations ranges from 50, 100, 150 and 200 mM of NaCl. The extracts were sprayed on to the leaf of the salt treated and control plants after the first stress treatment (approximately 7 days after the salt stress treatment). The extract was applied to the salt stressed plants at every 10 days interval.
Random samples from each pots were analyzed for the various parameters like shoot length, root length, total fresh and dry weight, relative water content, Histochemical detection of hydrogen peroxide, ascorbate peroxidase, catalase, peroxidase. Total proline content was analyzed in the salt stressed (0-200 mM NaCl) and foliar sprayed plants (stress induced 0-200 mM NaCl). All the experiments were performed in four replicates and maintained at green house conditions.
Relative water content: Relative Water Content (RWC) of the leaf was estimated by measuring the Fresh Weight (FW) of the leaf, they were immersed in a plate containing distilled water for 7 h. Turgid leaf was quickly blotted with tissue study and then Turgid Weight (TW) was determined. Dry Weight (DW) was taken after oven drying at 80°C for 48 h, the time at which constant weight reached18.
Histochemical assay for H2O2 production in leaf: The accumulation of H2O2 production in tomato leaf was analyzed using 3,3’-diaminobenzidine (DAB) staining19. The DAB staining solution (0.1% w/v DAB) and 10 mM MES (pH: 6.5) forms a deep brown polymerization product upon reaction with H2O2 in the presence of peroxidase. The seagrass sprayed tomato plant leaves were infiltrated in 10 mL staining solution in a syringe. The leaves were incubated for 24 h under normal illumination of light and destained with 90% ethanol for 12 h20.
Enzyme extraction for antioxidant enzymes: About 200 mg of leaves were homogenized with 1 mL of extraction buffer (50 mM potassium phosphate buffer, pH: 7.0 containing 1 mM EDTA and 1% polyvinylpyridine) using freezed mortar and pestle. In case of APX assay, 1 mM ascorbic acid was added in addition to the extraction buffer. The homogenate was centrifuged at 10,000 rpm for 5 min and supernatant was recovered for enzymes assay. The protein content was determined according to Bradford21 with bovine serum albumin as standard.
Superoxide dismutase: The treated and untreated samples were assayed by its ability to inhibit photochemical reduction of NBT (Nitro blue tetrazolium chloride) at 560 nm. Riboflavin was added to reaction mixture at last and illuminated for 15 min at light intensity of 5,000 lux. The reaction mixture without enzyme, kept in the light was treated as control and the dark served as blank. One unit of SOD (EC 1.15.1.1) activity was defined as the amount of enzymes required to cause 50% inhibition of the reduction of NBT as monitored at 560 nm22.
Peroxidase: The POX (EC 1.11.1.7) was quantified using the reaction mixture consists of 50 mM phosphate buffer (pH: 6.8), 4.6 mM phenylenediamine hydrochloride, 8.8 mM H2O2 and 100 μL enzyme extract. The changes in the absorbance were measured at 485 nm for 5 min and expressed as U mg–1 protein23.
Catalase: The CAT (EC 1.11.1.6) activity was quantified using the reaction mixture contains 50 mM potassium phosphate buffer (pH: 7.0), 20 mM H2O2 and 200 μL enzyme extract. The change in absorbance was recorded at 240 nm for 5 min and the catalase activity was calculated using the coefficient of absorbance of 43.6 mM–1 cm–1. The reaction was started by the addition of H2O2 and the enzyme activity was calculated in terms of 1 μmol H2O2 destroyed per min is defined as one unit24.
Ascorbate peroxidase: The ascorbate peroxidase (EC 1.11.1.11) activity was determined by decrease in absorbance at 290 nm (Extinction coefficient of 2.8 mM–1 cm–1) as ascorbate is oxidized25. The reaction mixture contains 50 mM phosphate buffer pH: 7.0, 0.5 mM AsA, 1 mM H2O2, 0.1 mM EDTA and 200 μL of enzyme extract. The enzyme activity was calculated in terms of μmol of ascorbate oxidized per min.
Determination of non-enzymatic antioxidants
Determination of free proline: The Proline content was estimated using ninhydrin reaction. Approximately100 mg of leaves was homogenized with 2 mL of 3% (W/V) sulphosalicylic acid centrifuged at 8000 rpm for 5 min and then 1.5 mL of glacial acetic acid was added. The mixture was incubated 100°C water bath for 30 min and the reaction was terminated in an ice bath. Proline content was calculated from known concentration of L-proline26.
Fourier transform infrared spectroscopy analysis of Zostera marina: The fine solid samples of Zostera marina was mixed with the potassium bromide (KBr) by the KBr pellet technique. The pellet was read spectrophotometrically by using the Bio Rad FTIR at the IR ranges between 400-4000 cm and the frequencies of different components present in the sample was recorded.
Preparation of extracts and GCMS conditions
Preparation of extracts: The crude Zostera marina powders were extracted (submerged extraction method) with methanol successively at room temperature for 15 days. About 10 g of each powdered material was soaked in 100 mL of methanol. The solvents were filtered and evaporated at 37°C under reduced pressure27. Crude methanol fraction of Zostera marina was subjected to routine qualitative chemical analysis to identify the nature of constituents present in them.
GC-sMS conditions: The GC-MS analysis of Zostera marina extract was performed using Thermo GC- TRACE ULTRA ver: 5.0 Thermo MS DSQ II system equipped with DB 35-MS capillary standard non polar column with dimension of 30 MTS×0.25 mm×0.25 μm. Helium was used as a carrier gas at a flow rate of 1 mL min–1. The oven temperature was initially operated at 70°C and gradually raised to 250 at 10°C min–1. About 1 μL of sample was injected in to the system and the total run time is about 38 min. The compounds were interpreted by comparing with the mass spectra of the National Institute of standard and Technology (NIST) library.
Statistical analysis: The study was carried out based on a completely randomized design with each treatment replicated three times and each experiment was performed three times. Tomato plants without salt stress served as a control at every step of the analysis. The statistical significance of the data obtained from this study was determined by one-way analysis of variance (SPSS version 17). The mean values were compared by Duncan’s multiple range test (DMRT; p<0.05) and the percentage of response was scored on the basis of DMRT analysis.
RESULTS
Effect of initial stress induction to tomato plants: The tomato seeds were germinated in the experimental pots. The stress was given at different concentration ranges from 0-200 mM NaCl. Plants treated with 100, 150 and 200 mM of NaCl had shown stunted growth in leaf area and height of the plant when compared to the control.
Experiment I: The foliar spray of Zostera marina seagrass extract was given to one set of NaCl stress induced plants with 3 sprays at an interval of 10 days and the plant was not watered for 3 days thereafter. It was observed that plants sprayed with Zostera marina extract shown increase in their shoot length.
Experiment II: In the pot studies, plants at higher concentrations of NaCl, showed retardation in the morphological parameters (Fig. 1). In this set of experiment, the plants sprayed with seagrass extract shown increase in the number of leaves than the plants sprayed with water.
 | |
| Fig. 1: | Morphological variation of control and salt stressed tomato plants after 15 days |
a: Control plant, b: 50 mM NaCl treated plants, c: 100 mM NaCl treated plants, d: 150 mM mM NaCl treated plants and e: 200 mM NaCl treated plants | |
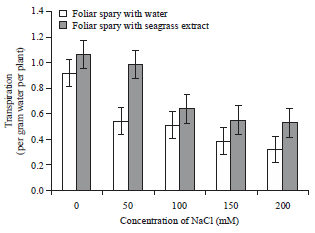 | |
| Fig. 2: | Effect of Zostera marina foliar spray treatment on relative water content of salt stressed tomato plants |
Error bars equal to standard deviation of ten replicates and experiment of repeated thrice. The data was determined by one-way analysis of variance (ANOVA) (SPSS v. 17). The mean values were compared by using Duncan’s multiple range test (p<0.05) | |
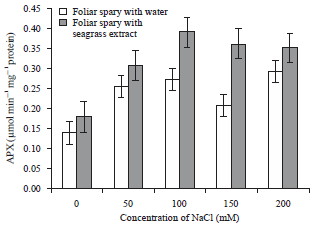 | |
| Fig. 3: | Effect of Zostera marina foliar spray treatment on ascorbate peroxidase enzyme activity of salt stressed tomato plants |
Error bars equal to standard deviation of ten replicates and experiment of repeated thrice. The data was determined by one-way analysis of variance (ANOVA) (SPSS v. 17). The mean values were compared by using Duncan’s multiple range test (p<0.05) | |
Transpirational water loss: The Relative Water Content (RWC) of the salt stressed tomato plants were declined with increase induction of salt stress from 0-200 mM. The plants sprayed with water exhibited 0.915- 0.32% in RWC. The plants sprayed with seagrass extract shown 1.58-0.52% of RWC. This indicates that the plants sprayed with seagrass extract shown an increase in RWC compared to the water sprayed plants and their respective controls. The foliar sprayed plants with extract had shown an early recovery from the NaCl stress by gradual increase in the transpirational rate.
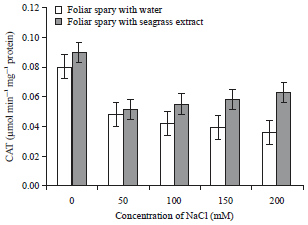 | |
| Fig. 4: | Effect of Zostera marina foliar spray treatment on catalase enzyme activity of salt stressed tomato plants |
Error bars equal to standard deviation of ten replicates and experiment of repeated thrice. The data was determined by one-way analysis of variance (ANOVA) (SPSS v. 17). The mean values were compared by using Duncan’s multiple range test (p<0.05) | |
But the tomato plants sprayed with water not show much variation in recovering the stress conditions (Fig. 2) and the plant leaves shrunk quickly as compared to the extract treated plant pots. The control plants exhibited normal transpirational rate than the stressed plants.
Effect of NaCl stress on antioxidant enzyme activities: The enzymatic activity of ascorbate peroxidase was increased significantly in control and extract treated salt stressed plants. The plants foliar sprayed with water shown increase in the APX activity at 50 and 100 mM NaCl treatment than other treatments. The activity of the extract treated plants exhibited increase in the concentration of enzymes (0.391±0.01 μmol min–1 mg–1 protein) at 100 mM NaCl stressed plants (Fig. 3). The extract treated control plants had shown slight increase in the activity (0.178±0.013 μmol min–1 mg–1 protein) than the plants sprayed with water (0.138±0.016 μmol min–1 mg–1 protein).
The Zostera marina treated tomato plants shown increase in the catalase enzymatic activity than the plants sprayed with water (Fig. 4). The Control plants exhibited 0.080±0.001 μmol min–1 mg–1 protein, more or less the foliar sprayed control plants of shown same level of CAT enzymatic activity. Similarly, the activity of catalase (0.051±0.001 to 0.090±0.001 μmol min–1 mg–1 protein) was observed to increase gradually with respect to increase in NaCl concentration. In case of water treated plants, the enzyme activity decreased to 0.036±0.014 μmol min–1 mg–1 protein, increase in the concentration of NaCl leads to the decrease in the enzyme activity. It was observed that the activity of SOD shown serious variations in recovering the stress. Increase in the concentration of stress had lead to the decrease in the SOD (Fig. 5).
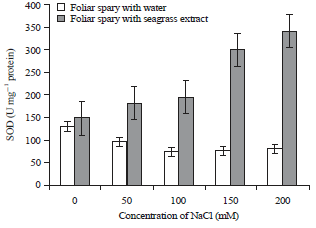 | |
| Fig. 5: | Effect of Zostera marina foliar spray treatment on superoxide dismutase enzyme activity of salt stressed tomato plants |
Error bars equal to standard deviation of ten replicates and experiment of repeated thrice. The data was determined by one-way analysis of variance (ANOVA) (SPSS v. 17). The mean values were compared by using Duncan’s multiple range test (p<0.05) | |
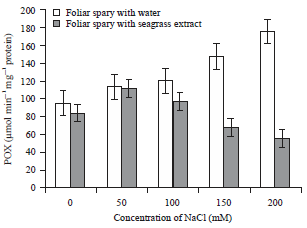 | |
| Fig. 6: | Effect of Zostera marina foliar spray treatment on peroxidase enzyme activity of salt stressed tomato plants |
Error bars equal to standard deviation of ten replicates and experiment of repeated thrice. The data was determined by one-way analysis of variance (ANOVA) (SPSS v. 17). The mean values were compared by using Duncan’s multiple range test (p<0.05) | |
In contrast to the CAT, APX and SOD, the POX activity of the enzyme increased at the stressed conditions. The stress recovered plants shown decrease in the activity than the control. The highest activity was observed in the 200 mM NaCl stressed plants of about 175.9±0.10 μmol min–1 mg–1 protein. Whereas the stress recovered plants (Foliar sprayed with extract) shown only 55.5±0.16 μmol min–1 mg–1 protein (Fig. 6).
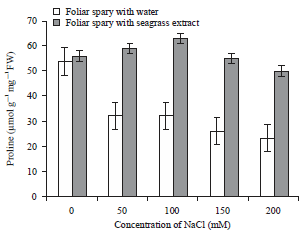 | |
| Fig. 7: | Effect of Zostera marina foliar spray treatment on proline activity of salt stressed tomato plants |
Error bars equal to standard deviation of ten replicates and experiment of repeated thrice. The data was determined by one-way analysis of variance (ANOVA) (SPSS v. 17). The mean values were compared by using Duncan’s multiple range test (p<0.05) | |
Effect of NaCl stress on non antioxidant enzyme activities: The level of proline increases proportionately faster than other amino acids in plants under stress conditions. The plants sprayed with extracts shown increase in the activity at maximum 100 mM of NaCl and it shown the maximum recovery from stress than the 150 mM and 200 mM of NaCl concentration. The proline activity also decreased at the higher concentration of stress (Fig. 7).
Histological localization of O2‾ and H2O2: The salt stressed plants showed an increase in the accumulation of H2O2 in the leaves of tomato plants. The control plants have shown less or minimal brown color patches in the leaves, but increase in the concentration of salt stress increased the brown spots in the leaves due to the reaction of DAB in the presence of peroxidases to produce a brown polymerization product. Considerable more H2O2 was detected in the tip of the leaf (Fig. 8).
Stress recovery and the effect of foliar spray: The seagrass extract sprayed plants shown the stress recovery after 30 days, whereas the plant sprayed with ordinary water, shown low survival rate and the leaves became yellowish color and the density of the plants reduced to the minimal level. Hence, the Zostera marina foliar sprayed plants showed increase in the shoot length and biomass than the control.
FTIR analysis of Zostera marina: The functional groups present in the seagrass includes alcohol (0-H), alkane (C-H), alkene (C=C), alkyl halides like (C-F and C-Cl), alkyne (-C≡C-), amine (C-N) and aromatic (C=C) groups (Fig. 9). The characteristic IR absorption frequencies and intensity of bands of organic functional groups of Zostera marina were analyzed (Table 1).
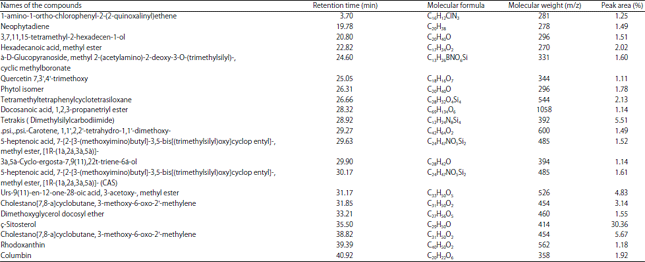
GC-MS analysis: The gas chromatography mass spectral analysis of the methanolic extract revealed the presence of 21 compounds (Table 2). The major compound present in the extract was ç-Sitosterol (30.36%) at 35.50 retention time. The cholestano[7,8-a]cyclobutane, 3-methoxy-6-oxo-2'-methylene was found to be the next compound present at higher concentration with peak area of about 5.67% at 38.82 retention time. Tetrakis (Dimethylsilylcarbodiimide) was found at 28.92 retention time and the peak area was about 5.51%. Cholestano[7,8-a]cyclobutane, 3-methoxy-6-oxo-2'-methylene an compound with unknown properties were found at retention time of 31.85 with peak area of 3.14% (Fig. 10).
 | |
| Fig. 8: | Histochemical DAB staining assay for H2O2 accumulation in leaves of salt stressed tomato plants |
The brown patches in the leaves indicate the production of hydrogen peroxide at different concentration of NaCl stress, a: Control plant, b: 50 mM NaCl treated plants, c: 100 mM NaCl treated plants, d: 150 mM mM NaCl treated plants, e: 200 mM NaCl treated plants | |
DISCUSSION
This study represents that the seagrass extract has shown a protective mechanism under saline conditions by the increased production of antioxidant enzymes. The compounds present in the seagrass extract act as an elicitors for the tolerance. Marine resources such as seaweeds and seagrass have been documented for several decades because of their innumerable applications. Besides their application as farmyard manure, liquid extracts obtained from these resources have gained importance as foliar sprays for crop plant28. It is evident that the extract contains growth promoting hormones such as auxins, cytokinins, gibberellins, abscisic acid and brassinosteroids29. The enhanced growth plants by seaweed extract is due to the presence of inorganic compounds like vitamins, micro and macro nutrients30, organic compounds like betaines31, polysaccharides32 and phenolic compounds33 in the extracts. It is well known that one of the first plant responses to salinity stress is a reduction in leaf growth rate with associated reductions in leaf area available for photosynthesis. Subsequently, excessive accumulation of salts can lead to death of tissues, organs, toxicity, osmotic stress and production of reactive oxygen species11. The ROS can be scavenged by superoxide dismutase (SOD), catalase (CAT) and variety of peroxidases (POX)12.
The total biomass production of the plants has been inhibited by salinity stress. Increase in the dose of the salt from 0-200 mM reduces the Relative Water Content (RWC). The extract treated plants showed higher RWC than the plants sprayed with water. Chlorophyll content of the extract treated plants was observed to be higher than control plants under saline conditions. Similar observations of increased chlorophyll content were reported in Brassica chinensis and Phaseolus vulgaris after foliar application of seaweed extract34,35. In this study, chlorophyll content was reduced under stress conditions whereas extract treated plants showed higher chlorophyll content than water sprayed control plants. This observation clearly represents the positive role of seagrass extract in increased chlorophyll content under green house condition (Fig. 10).
| Table: 1 | FTIR analysis infrared absorption and structure of functional groups present in the Zostera marina |
 | |
| Table 2: | Chemical composition of Zosterna marina extract by GCMS analysis |
 | |
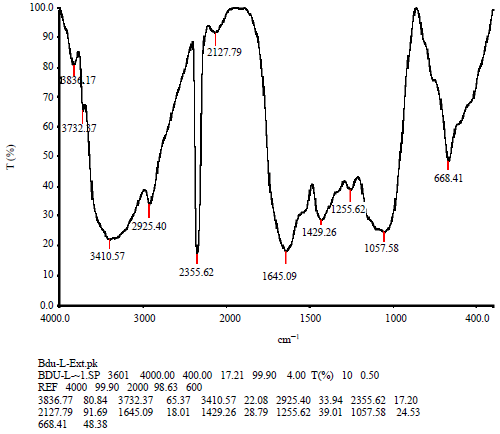 | |
| Fig. :9 | FTIR analysis of Zostera marina, the peaks represents the chemical groups present in the sample |
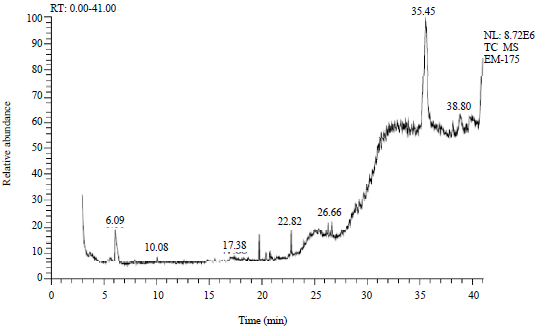 | |
| Fig. 10: | GC-MS analysis of Zostera marina |
The plants sprayed with water shown decline in chlorophyll content and the leaves became yellowish in color and the number of plants have been reduced due o the sensitivity to the NaCl. But the plants sprayed with the extract have enhanced the tolerance to the different concentrations of saline conditions. This observation was supported by the results of Mahmud et al.36 in which chlorophyll content of untreated rice plants was reduced whereas methyl jasmonate treated plants showed higher chlorophyll content under salt stress condition.
In pot studies, both the shoot length and root length were found to increase in the seagrass foliar sprayed plants when compared to control plants. This result is in accordance with the observations made by Mansori et al.34, in which seaweed treated Phaseolus plant showed increased length of shoot and root. The enzyme activity of the APX in the plants subjected to saline conditions suggests that oxidative stress is an important component of salt stress and it scavenges peroxides by converting ascorbic acid to dehydroascorbate37. In this study, three folds of higher activity of APX were seen in the extract treated plants compared to the control and plants sprayed with water. The APX of the more active ascorbate-glutathione cycle may be related to the development of relatively higher salt tolerance in plants38. Our results are in correlation with various others findings that the APX and GR activities have been increased in the saline conditions.
The SOD is responsible for the scavenging of toxic O2‾ in different cell organelles. Salinity have induced increased SOD activity as reported by various workers. The numerous number of reports indicated that the biotic and abiotic stress alters the amount and activities of the enzymes, the enhanced SOD activity39,40. In our experiment, the activity of the extract treated plants shown 4 fold increase in enzymes at 200 mM concentration of NaCl.
The CAT is another enzyme which is also involved in H2O2 detoxification converting H2O2 into water and oxygen. El-Sharkawy et al.15 reported the enhancement of CAT activity in both salt sensitive and salt tolerant cultivars of Alfalfa. Whereas, in this study, CAT activity was found to decrease with increasing concentration of salt, but the plants sprayed with extract shown increase in the CAT activity in 50, 100, 150 and 200 mM NaCl treated plants.
In contrast to the APX, SOD and CAT activity, the POX activity has been declined in the extract treated tomato plants. But the plants sprayed with water shown increase in the activity of enzymes from 96.6±0.16 to 175.9±0.10 at 200 mM concentration of NaCl. In contrast the activity of enzyme is very low and shown nearly threefold decrease in the activity at 200 mM concentration. The up-regulation and down regulation of the antioxidant activities depend on the crop varieties and the type of abiotic stress. It is reported that plants sprayed with seaweed extracts also exhibit enhanced salt and freezing tolerance10. Our study revealed that the foliar sprayed salt stressed tomato plants exhibit increased SOD, CAT and APX than control plants. It indicates that the natural level of tolerance occurs to the seagrass treated plants in-spite of the character gene transfer technology. The GC-MS and FTIR results indicated that the presence of various constituents that all together paves the way for the salt tolerance. This work suggests that the extract can be recommended as foliar spray and biofertilizer to increase the yield of the different crops in an environment friendly manner. The level of tolerance in response to the stress can be further investigated in an innovative manner for the commercial use of the extract.
CONCLUSION
It is concluded that the present study provides the possibility of increased salt tolerance in tomato plants through foliar application of seagrass liquid extract. To the nest of our knowledge this is the first report on the application of seagrass liquid extract in salt stress alleviation of tomato. Further this work can be extended to other economically important crops as well to create tolerance for abiotic and biotic stress conditions.
SIGNIFICANCE STATEMENTS
This study discovers the positive role of Zostera marina extract to give tolerance to tomato plants under salt stress condition. The findings of this study can be beneficial to enhance tolerance under other abiotic stresses as well. This study will help the researcher to increase tolerance in salt sensitive genotypes of economically important crop plants by using Zostera marina extract as a stress alleviator. Thus, a new theory on the effect of this seagrass on stress alleviation may be arrived at.
ACKNOWLEDGMENTS
The authors are thankful for the financial support assisted by the Council of Scientific and Industrial Research (CSIR), New Delhi, India through CSIR_SRF (Order No. 09/475(0189)/2012-EMR-I Dated:19.03.13) and University Grants Commission (UGC), New Delhi India. The authors also thank SITRA, Coimbatore College Tamilnadu for GCMS instrumentation and support in analyzing the samples.
REFERENCES
- Hasanuzzaman, M., K. Nahar and M. Fujita, 2013. Plant Response to Salt Stress and Role of Exogenous Protectants to Mitigate Salt-Induced Damages. In: Ecophysiology and Responses of Plants under Salt Stress, Ahmad, P., M.M. Azooz and M.N.V. Prasad (Eds.), Springer, New York, United State, ISBN-13: 9781461447467, pp: 25-87.
CrossRefDirect Link - Moghadam, A.A., E. Ebrahimie, S.M. Taghavi, A. Niazi and M.Z. Babgohari et al., 2013. How the nucleus and mitochondria communicate in energy production during stress: Nuclear MtATP6, an early-stress responsive gene, regulates the mitochondrial F1F0-ATP synthase complex. Mol. Biotechnol., 54: 756-769.
Direct Link - Hasanuzzaman, M., M.A. Hossain, J.A.T. da Silva and M. Fujita, 2012. Plant Responses and Tolerance to Abiotic Oxidative Stress: Antioxidant Defenses is a Key Factors. In: Crop Stress and its Management: Perspectives and Strategies, Bandi, V., A.K. Shanker, C. Shanker and M. Mandapaka (Eds.). Springer, Germany, pp: 261-316.
- Munns, R. and M. Tester, 2008. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol., 59: 651-681.
CrossRefDirect Link - Raiola, A., M.M. Rigano, R. Calafiore, L. Frusciante and A. Barone, 2014. Enhancing the health-promoting effects of tomato fruit for biofortified food. Mediators Inflammation, Vol. 2014.
CrossRef - Gerszberg, A. and K. Hnatuszko-Konka, 2017. Tomato tolerance to abiotic stress: A review of most often engineered target sequences. Plant Growth Regul.
Direct Link - Stamatiadis, S., L. Evangelou, J.C. Yvin, C. Tsadilas, J.M.G. Mina and F. Cruz, 2015. Responses of winter wheat to Ascophyllum nodosum (L.) Le Jol. extract application under the effect of N fertilization and water supply. J. Applied Phycol., 27: 589-600.
CrossRefDirect Link - Van Oosten, M.J., O. Pepe, S. Pascale, S. Silletti and A. Maggio, 2017. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric., Vol. 4.
CrossRef - Mancuso, S., E. Azzarello, S. Mugnai and X. Briand, 2006. Marine bioactive substances (IPA extract) improve foliar ion uptake and water stress tolerance in potted Vitis vinifera plants. Adv. Hortic. Sci., 20: 156-161.
Direct Link - Sharma, P., A.B. Jha, R.S. Dubey and M. Pessarakli, 2012. Reactive oxygen species, oxidative damage and antioxidative defense mechanism in plants under stressful conditions. J. Bot., Vol. 2012.
CrossRefDirect Link - Das, K. and A. Roychoudhury, 2014. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci., Vol. 2.
CrossRefDirect Link - Singh, R., S. Singh, P. Parihar, R.K. Mishra and D.K. Tripathi et al., 2016. Reactive Oxygen Species (ROS): Beneficial companions of plant's developmental processes. Front. Plant Sci., Vol. 7.
CrossRefDirect Link - Kim, Y.H., A.L. Khan, M. Waqas and I.J. Lee, 2017. Silicon regulates antioxidant activities of crop plants under abiotic-induced oxidative stress: A review. Front. Plant Sci., Vol. 8.
CrossRefDirect Link - Racchi, M.L., 2013. Antioxidant defenses in plants with attention to Prunus and Citrus spp. Antioxidants, 2: 340-369.
CrossRefDirect Link - El-Sharkawy, M., T. El-Beshsbeshy, R. Al-Shal and A. Missaoui, 2017. Effect of plant growth stimulants on alfalfa response to salt stress. Agric. Sci., 8: 267-291.
CrossRef - Pan, Y., L.J. Wu and Z.L. Yu, 2006. Effect of salt and drought stress on antioxidant enzymes activities and SOD isoenzymes of liquorice (Glycyrrhiza uralensis Fisch). Plant Growth Regul., 49: 157-165.
CrossRefDirect Link - Kumar, G. and D. Sahoo, 2011. Effect of seaweed liquid extract on growth and yield of Triticum aestivum var. Pusa Gold. J. Applied Phycol., 23: 251-255.
CrossRefDirect Link - Smart, R.E. and G.E. Bingham, 1974. Rapid estimates of relative water content. Plant Physiol., 53: 258-260.
CrossRefDirect Link - Thordal-Christensen, H., Z. Zhang, Y. Wei and D.B. Collinge, 1997. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley-powdery mildew interaction. Plant J., 11: 1187-1194.
CrossRefDirect Link - Folhe, L. and F. Otting, 1984. Superoxide dismutase assays. Methods Enzymol., 105: 93-104.
PubMedDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Beauchamp, C. and I. Fridovich, 1971. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem., 44: 276-287.
CrossRefPubMedDirect Link - Kokkinakis, D.M. and J.L. Brooks, 1979. Tomato peroxidase: Purification, characterization and catalytic properties. Plant Physiol., 63: 93-99.
CrossRefDirect Link - Patterson, B.D., L.A. Payne, Y.Z. Chen and D. Graham, 1984. An inhibitor of catalase induced by cold in chilling-sensitive plants. Plant Physiol., 76: 1014-1018.
CrossRefDirect Link - Nakano, Y. and K. Asada, 1981. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in Spinach chloroplasts. Plant Cell Physiol., 22: 867-880.
CrossRefDirect Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Vinoth, S. , P.R. Kanna, P. Gurusaravanan and N. Jayabalan, 2011. Evaluation of phytochemical, antimicrobial and GC-MS analysis of extracts of Indigofera trita L.F. spp. subulata (vahl ex poir). Int. J. Agric. Res., 6: 358-367.
CrossRefDirect Link - Battacharyya, D., M.Z. Babgohari, P. Rathor and B. Prithiviraj, 2015. Seaweed extracts as biostimulants in horticulture. Sci. Hortic., 196: 39-48.
CrossRefDirect Link - Stirk, W.A., D. Tarkowska, V. Turecova, M. Strnad and J. van Staden, 2014. Abscisic acid, gibberellins and brassinosteroids in Kelpak®, a commercial seaweed extract made from Ecklonia maxima. J. Applied Phycol., 26: 561-567.
Direct Link - Rayirath, P., B. Benkel, D.M. Hodges, P. Allan-Wojtas, S. MacKinnon, A.T. Critchley and B. Prithiviraj, 2009. Lipophilic components of the brown seaweed, Ascophyllum nodosum, enhance freezing tolerance in Arabidopsis thaliana. Planta, 230: 135-147.
CrossRefDirect Link - MacKinnon, S.L., D. Hiltz, R. Ugarte and C.A. Craft, 2010. Improved methods of analysis for betaines in Ascophyllum nodosum and its commercial seaweed extracts. J. Applied Phycol., 22: 489-494.
CrossRefDirect Link - Khan, W., U.P. Rayirath, S. Subramanian, M.N. Jithesh and P. Rayorath et al., 2009. Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth Regul., 28: 386-399.
CrossRefDirect Link - Wang, T., R. Jonsdottir and G. Olafsdottir, 2009. Total phenolic compounds, radical scavenging and metal chelation of extracts from Icelandic seaweeds. Food Chem., 116: 240-248.
CrossRefDirect Link - Mansori, M., H. Chernane, S. Latique, A. Benaliat, D. Hsissou and M. El Kaoua, 2015. Seaweed extract effect on water deficit and antioxidative mechanisms in bean plants (Phaseolus vulgaris L.). J. Applied Phycol., 27: 1689-1698.
CrossRefDirect Link - Zheng, S., J. Jiang, M. He, S. Zou and C. Wang, 2016. Effect of kelp waste extracts on the growth and development of Pakchoi (Brassica chinensis L.). Sci. Rep., Vol. 6.
CrossRef - Mahmud, S., S. Sharmin, B.L. das Chowdhury and M.A. Hossain, 2017. Effect of salinity and alleviating role of methyl jasmonate in some rice varieties. Asian. J. Plant Sci., 16: 87-93.
CrossRefDirect Link - Ozkur, O., F. Ozdemir, M. Bor and I. Turkan, 2009. Physiochemical and antioxidant responses of the perennial xerophyte Capparis ovata Desf. to drought. Environ. Exp. Bot., 66: 487-492.
CrossRefDirect Link - Pandey, S., D. Fartyal, A. Agarwal, T. Shukla and D. James et al., 2017. Abiotic stress tolerance in plants: Myriad roles of ascorbate peroxidase. Frontiers Plant Sci., Vol. 8.
CrossRef - Hossain, M., M.R. Ismail, M.K. Uddin, M.Z. Islam and M. Ashrafuzzaman, 2013. Efficacy of ascorbate-glutathione cycle for scavenging H2O2 in two contrasting rice genotypes during salinity stress. Aust. J. Crop Sci., 7: 1801-1808.
Direct Link - Lu, F., X. Liang, H. Lu, Q. Li and Q. Chen et al., 2017. Overproduction of superoxide dismutase and catalase confers cassava resistance to Tetranychus cinnabarinus. Sci. Rep., Vol. 7.
CrossRef