
J.C. N`Zi
World Agroforestry Centre (ICRAF) Cote d`Ivoire Country Program, BP 2823 Abidjan 08, Cote d`Ivoire
A.P. Koua
Universite Felix Houphouet-Boigny, Unit of Formation and Research-Biosciences, 22 BP 582 Abidjan 22, Cote d`Ivoire
J. Kahia
World Agroforestry Centre (ICRAF) Cote d`Ivoire Country Program, BP 2823 Abidjan 08, Cote d`Ivoire
K.D. Kouassi
Societe de Caoutchouc de Grand-Bereby, BP 365 Abidjan 01, San-Pedro, Cote d`Ivoire
A.S.P. N`guetta
Universite Felix Houphouet-Boigny, Unit of Formation and Research-Biosciences, 22 BP 582 Abidjan 22, Cote d`Ivoire
C. Kouame
World Agroforestry Centre (ICRAF) Cote d`Ivoire Country Program, BP 2823 Abidjan 08, Cote d`Ivoire
Asian Journal of Plant Sciences
Year: 2016 | Volume: 15 | Issue: 1-2 | Page No.: 26-34







ABSTRACT
Recent survey in the Soubré region of Côte d’Ivoire has showed high soil fertility depletion. The cost of mineral fertilizer is high and does not always yield a positive return on investment. Soybean can use atmospheric nitrogen through biological fixation by establishing a symbiotic relationship with Rhizobium bacteria. Nitrogen (N) is subject to intense chemical and microbiological transformation processes and is, therefore a very mobile nutrient element in the soil-plant-atmosphere continuum. A study was conducted at Logboayo and Soubré to evaluate the effect of IRAT-FA3 Bradyrhizobium japonicum strain inoculum on the nodulation and some growth parameters of five soybean varieties (Glycine max) named Canarana, Doko, V3-2013, V6-2013 and IT-235. A randomized complete block design with a split plot replicated three times was used. Inoculation was the main factor and variety the subplot treatment. Data were collected on seedling emergence, growth parameters and nodulation. Results showed that lowest emergence time of 6.49 Days After Sowing (DAS) was recorded at Soubré compared to 6.91 DAS for Logboayo. Doko variety had the highest number (11) of nodules per plant in inoculated treatments. The variety V3-2013 showed ability to nodulation with indigenous rhizobia. Seeds inoculation resulted in an increase of nodules and growth of organs. The results of the present study opens up option for soybean to be grown along other crops in order to address soil fertility problems in the project area.
PDF Abstract XML References Citation
Received: November 24, 2015;
Accepted: January 30, 2016;
Published: March 15, 2016
How to cite this article
J.C. N`Zi, A.P. Koua, J. Kahia, K.D. Kouassi, A.S.P. N`guetta and C. Kouame, 2016. Evaluating Nodulation and its Effects on Some Agromorphological
Parameters of Soybean Varieties (Glycine max L.). Asian Journal of Plant Sciences, 15: 26-34.
DOI: 10.3923/ajps.2016.26.34
URL: https://scialert.net/abstract/?doi=ajps.2016.26.34
DOI: 10.3923/ajps.2016.26.34
URL: https://scialert.net/abstract/?doi=ajps.2016.26.34
INTRODUCTION
Soybean (Glycine max L.) is an annual herbaceous plant of the family of Fabaceae native of Eastern Asia. It is a legume; cultivated for its seeds, which are highly rich in protein and oil (FAO., 1995; Nyabyenda, 2005). Soybean can use atmospheric nitrogen through biological fixation by establishing a symbiotic relationship with Rhizobium bacteria (Van der Vossen et al., 2007). Nitrogen (N) is subject to intense chemical and microbiological transformation processes (hydrolysis, oxidation, reduction) and is, therefore a very mobile nutrient element in the soil-plant-atmosphere continuum (Stahl et al., 2002; Phongpan and Mosier, 2003).
As nitrogen is the most limiting nutrient for growth of leguminous plants because its absence in the soil cannot support growth (Simon et al., 2014), it is essential in plant cells for synthesis of enzymes, proteins, chlorophyll, DNA and RNA, plant growth and production of food and feed (Matiru and Dakora, 2004). On the other hand, rhizobia are very important for crop production because they form symbiotic relationship with legume the process that converts atmospheric elemental nitrogen into ammonia accounting for 65% of the nitrogen presently utilized in agriculture (Matiru and Dakora, 2004; Simon et al., 2014). Bradyrhizobium japonicum is a nitrogen-fixing soil bacterium that has a symbiotic relationship with the host legume soybean Glycine max (Lee et al., 2012). Such symbiosis makes soybean less dependent on soil nitrogen particularly in tropical regions of Africa, where soils are deficient in nitrogen (FAO., 1995). Therefore, legume-rhizobia symbiosis can provide easy and inexpensive way to enhance soil fertility and improve crop production (Roychowdhury et al., 2013). Legumes can grow in much degraded soils because they have the ability to fix nitrogen in association with rhizobia (Wong, 2003; Freitas et al., 2004). Biological nitrogen fixation has gained attention in recent years because it substitutes inorganic fertilizer and is environmental friendly farm input essential for poor resource farmers (Bekere and Hailemariam, 2012). It also limits groundwater’s pollution by nitrates (Berrada and Fikri-Benbrahim, 2014). As a nodulating legume, soybean improves soil fertility by fixing nitrogen from the atmosphere (Danso, 1992). The nitrogen fixed in the nodules by the N-fixing rhizobia is subsequently made available for plant growth. This is a major benefit in African farming systems, where soils have become exhausted by continuous cropping. This nitrogen source is important considering the scarcity and high cost of nitrogen fertilizers (Abana et al., 2015).
In Côte d’Ivoire, most soils are deficient in nitrogen and this deficiency is the most prominent constraint to food production (Bognonkpe and Becker, 2009). According to Nhamo et al. (2003) and Stajkovic et al. (2011), besides their major role in the traditional diets throughout the world, legumes provide a multiple benefits to both soil and other crops through intercropping. The present study is part of a wide program geared towards addressing food security in the Nawa region of Côte d’Ivoire (N’Zi et al., 2015). The objective of this study was to investigate the effect of IRAT-FA3 Bradyrhizobium japonicum strain inoculum on the nodulation and its effect on some growth parameters of five soybean varieties.
MATERIALS AND METHODS
Study site: Experiments were conducted at the World Agroforestry Centre (ICRAF) Stations in Logboayo and Soubré, located in the Nawa Region; South-West of Côte d’Ivoire, 5°47 08"N, 6°36 30"W, 276 m a.s.l. The mean annual rainfall is 1360 mm and varies from 968-1767 mm. The minimum and maximum temperature varies from 23-36°C. The land in the two sites was previously planted with yams. The experimental plots were located on the top of a slope gradient of 7 and 9% in Soubré and Logboayo, respectively. The soils characteristics are 5.6 for pH, 1.04% for soil organic carbon, 0.1% for total nitrogen and 7.7 mg kg–1 for available phosphorus (Diby et al., 2014). Potassium, magnesium and calcium were 0.13, 1.09 and 3.74 cmol kg–1, respectively. Sand content was 41%, while the fine particles (silt+clay) were 57%.
Plant material: Five soybean varieties named Canarana, Doko, V3-2013, V6-2013 and IT-235 were used. The seeds were provided by the Research Station of Food Crops of the Centre National de Recherche Agronomique (CNRA) in Bouaké in Côte d’Ivoire.
Bacterium strain: The IRAT-FA3 B. japonicum strain was used to produce the inoculum. The strains of this species are known for having a perfect symbiosis with soybean and are insignificant or absent in tropical soils (Cattelan and Hungria, 1995). It is described as being very competitive in acid soil and has already been used in Rwanda, Burundi, Madagascar, Cameroon and Côte d'Ivoire (Beugre et al., 2013). The inoculum was produced and provided by the Inoculum Production Unit for Legumes (UPIL) of the CNRA in Bouaké. Peat was used as inoculum substrate.
Methods
Experimental design: Two experiments were conducted. The first one was the germination test performed under a shade house. Ten seeds of each variety were sown in washed sand and lightly covered with sand. Germination was recorded when cotyledons emerged from the surface of the sand.
The second experiments were laid out in the field in a randomized complete block design with split plots arrangement replicated three times with two factors in each site. The main factor was the inoculation with two levels: Inoculated treatment and non-inoculated treatment. The second factor was the soybean variety, with five levels (Canarana, Doko, V3-2013, V6-2013 and IT-235). A block or replicate was divided into two sub-blocks or large plot. Each large plot was assigned with a treatment (Inoculated or non-inoculated). Each large plot was divided into as many elementary plots as soybean varieties.
The size of the sub plots was 4×3 m giving a total area of 12 m2 (Fig. 1). Two consecutive blocks were separated by a space of 2 m wide. Distance between 2 elementary plots of the same block was 1 m. A space of 2.5 m was left between the borders of the field and the blocks. Seeds were sowed at 50 cm spacing between the rows and 20 cm between the holes. The elementary plot of 12 m2 was represented by 7 lines of 4 m each. Data were collected from 5 lines on the inside of the basic plot 0.5 m from each side, on an area of 6 m2.
The site was cleared and this was followed by a manual deep plowing of 30 cm and making ridges. A fertilizer NPK 12-24-18 was applied at the rate of 100 kg ha–1. There was no pesticide application throughout the crop cycle. In addition, three manual weeding were made on the 15th, 30th and 55th Days After Sowing (DAS) to aerate the soil and reduce competition with weeds.
Seeds inoculation: The soybean seeds were inoculated with the B. japonicum bacterium using the seeds coating method as described by Vitosh (1997).
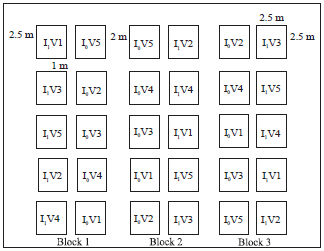 | |
| Fig. 1: | Experimental design, I0: Non-inoculated, I1: Inoculated and V: Varieties |
They were sown at a depth of 3 cm according to the protocol described by Kouame et al. (2007). The proportion was 100 g of inoculum for 15 kg of seed. The bacteria density was 109 bacteria/g of inoculum (N'Gbesso et al., 2010). The control plots were sowed first in order to avoid contamination by bacteria of the inoculum.
Data collection: Data were collected on seed germination under a shade house, seed emergence time and seed emergence percentage in the field, growth rate between 20 and 40 Days After Sowing (DAS), plant diameter at 40 DAS, plant height, number of leaves present on the main stem, number of active nodules per plant at 35 DAS, number of large nodules per plant (diameter = 2 mm) according to King and Purcell (2001), large active nodules per plant, small nodules per plant (diameter ranged from 1-2 mm) and small active nodules per plant at 50 DAS. Activity of nodules was evaluated by their coloration and the number of active or efficient nodules was counted after slicing them with a blade. According to Beuerlein (1997), only nodules with a bright red internal color are active and therefore have a maximum of nitrogen-fixing activity.
Statistical analysis: All data were subjected to analysis of variance (ANOVA) using the general linear model of Statistical Analysis System software (SAS., 2003). The significance differences between the means were assessed with Duncan method at 5%. Pearson correlation was used to examine relationships between variables.
RESULTS
Seed germination: The germination of the soybean seeds is shown in Fig. 2. The germination test showed that the first seeds sprouted 4 Days After Sowing (DAS). Doko variety had the highest germination rate of 47%. Six days after sowing, all varieties showed germination of 80%, except V6-2013 which had a germination of 60%. Eight days after sowing, the germination of varieties Canarana, Doko, V3-2013 and IT-235 were above 90%, compared to 70% recorded for V6-2013 (Fig. 3).
Seed emergence in the field: The time taken for the seeds to emerge was recorded starting from 4-11th day after sowing (Table 1). The analysis of variance showed a highly significant difference between the means of soybean varieties (p<0.0001) for this parameter. The seed inoculation showed no effect on the emergence time on both sites (p = 0.2869).
 | |
| Fig. 2(a-d): | Progression of the germination of the soybean seeds, (a) Plant at cotyledon stage, (b) Sprouted seeds, (c) Plants growing in the field and (d) Measurement of plant height |
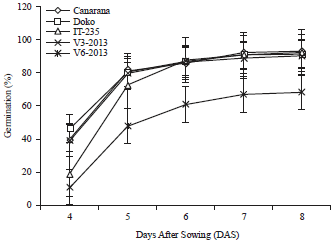 | |
| Fig. 3: | Progression of the germination of the soybean varieties |
Soubré recorded the lowest emergence time with 6.49 DAS compared to 6.91 DAS for Logboayo. The shortest emergence time was obtained with the variety IT-235 on both sites. The varieties V6-2013 and Doko took the longest time to germinate (7.04 and 7.38 DAS, respectively).
In the field the seeds emergence were observed in all varieties at 4 DAS. At 11 DAS, varieties Canarana, IT-235 and V3-2013 had more than 50% emergence while the varieties V6-2013 and Doko had lower than 50%.
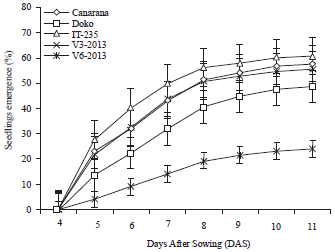 | |
| Fig. 4: | Progression of the emergence time of soybean varieties seedlings in field |
After this time, there was no seed emergence in all varieties (Fig. 4). The analysis of variance revealed a highly significant effect of soybean variety effect on seed emergence with p<0.0001, but the seed inoculation with p = 0.1949 did not influence this parameter.
| Table 1: | Evaluation of the seed emergence time (DAS) of the plants of soybean varieties |
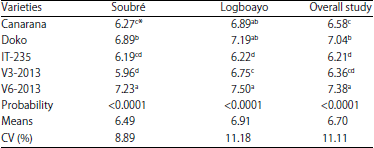 | |
| *: Numbers followed by the same letter in a column are statistically identical to the level of 5% (Duncan test) | |
| Table 2: | Evaluation of the growth parameters and the vegetative development of the plants of the varieties |
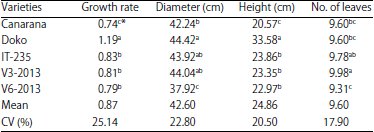 | |
| *: Numbers followed by the same letter in a column are statistically identical to the level of 5% (Duncan test) | |
Growth and plant development: Analysis of variance of plant height at 40 DAS and growth rate between 20 and 40 DAS, revealed a significant effect of the interaction treatment x variety (p<0.0012) and (p<0.0015), respectively (Table 2). Moreover, the variety significantly influenced the height and growth rate of plants with p<0.0001 for both parameters. During this period, variety Doko recorded the highest mean for the height (33.58 cm) and a growth rate of 1.19 cm day–1. The lowest height was observed in variety Canarana (20.57 cm) which also showed the lowest growth rate with 0.74 cm day–1. A highly significant effect of the variety on the plant height was observed with p<0.0001. For the plant diameter and the number of leaves on the main stem, results revealed that these parameters varied from one variety to another with the same probability p<0.0001. Varieties V3-2013 (9.98) and IT-235 (9.78) produced the highest number of leaves while the lowest values were observed with V6-2013 (9.31). Doko variety was observed to have the highest diameter (44.42 cm).
Plant nodulation and nodule activity: Figure 5 shows the effect of inoculation on nodulation and soybean development.
The results of the nodulation at 35 DAS showed that the treatment effect (p = 0.0248, p = 0.0155) and the variety effect (p = 0.0002, p = 0.0043) were significant for the total number of nodules per plant and the active nodules. The variety Doko had the highest number of nodules (Fig. 6). The interaction between variety and treatment (p = 0.0022, p = 0.0116) were highly significant for each of the parameters studied on both sites.
The varieties responded positively to inoculation with a significant increase in the number of nodules compared with non-inoculated (Fig. 6). The presence of nodules was observed in the region of 10 cm close to the neck of the varieties. Nodules of Doko variety were observed exclusively in neck area of the inoculated treatments in this study. In Soubré, non-inoculated of varieties Canarana and V3-2013 developed nodules, while that was not the case at Logboayo. As for the number of active nodules, the results showed that over 70% of nodules counted on inoculated and had a nitrogen-fixing activity. For the number of cumulative active nodules in inoculated and non-inoculated plants, the variety V3-2013 showed the best performance (Fig. 7).
The results revealed a significant increase in the number of nodule compared to non-inoculated varieties 50 DAS. Non-inoculated plants of the variety V3-2013 showed a higher number of nodules compared to the non-inoculated plants of the other varieties. Concerning the number of small nodules and their activity, significant differences were observed among inoculated varieties on one hand and on the other between the non-inoculated varieties. For the activity of nodules, 34.28 nodules per plant were active in inoculated varieties against 5.04 active nodules in non-inoculated varieties. It was also observed that variety V3-2013 and Doko had more small nodules. The variety V6-2013 had the lowest number of nodules with 20.2 small nodules and 15.4 active nodules per plant.
Variety V3-2013 produced more active nodules with 14.4 followed by IT-235 while variety V6-2013 recorded no active nodule (Table 3).
Parameters correlations: The correlation coefficients (r) between different parameters show that nodulation parameters are correlated positively (Table 4). The number of active nodules at 35 DAS is correlated to the number of large nodules with r = 0.896 and to the number of large active nodules (r = 0.902) harvested at 50 DAS. The number of plant leaves is positively correlated with the number of active nodules at 35 DAS (r = 0.939), as well as the number of large active nodules at 50 DAS (r = 0.940). The diameter of the plants correlates with the number of small nodules and small active nodules at 50 DAS, r = 0.879 and r = 0.899, respectively. The growth rate of the plants between 20 and 40 DAS is correlated with plant height (r = 0.999).
 | |
| Fig. 5(a-e): | Effect of inoculation on nodulation and leaf coloration, (a) Root system without nodules of non-inoculated soybean, (b) Root system with nodules of inoculated soybean, (c) Small and active (large) nodules, (d) Green foliage (see yellowing) of on-inoculated soybean plants and (e) Dark green foliage of inoculated soybean plants |
| Table 3: | Number of active nodules at 35 DAS and nodules per plant at 50 DAS |
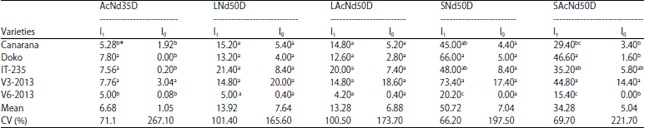 | |
| *: Numbers followed by the same letter in a column are statistically identical to the level of 5% (Duncan test) AcNd35D: No. of active nodules per plant at 35 DAS, LNd50D: No. of large nodules per plant at 50 DAS, LAcNd50D: No. of large active nodules per plant, SNd50D: No. of small nodules per plant, SAcNd50D: No. of active small nodules per plant, I0: Non-inoculated and I1: Inoculated | |
| Table 4: | Correlation coefficients (r) between nodulation and agromorphological parameters in soybean varieties |
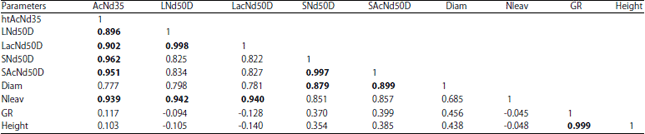 | |
| Bold values are different from 0 at a level of alpha = 0.05 significance AcNd35: No. of active nodules per plant at 35 DAS, LNd50D: No. of large nodules per plant at 50 DAS, LAcNd50D: No. of large active nodules per plant, SNd50D: No. of small nodules per plant, SAcNd50D: No. of active small nodules per plant, Diam: Plant diameter at 40 DAS, Nleav: No. of leaves, GR: Growth rate between 20 and 40 DAS and Height: Plant height at 40 DAS | |
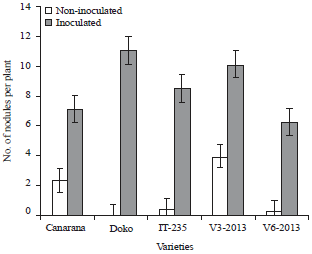 | |
| Fig. 6: | Effect of seed inoculation on nodulation of soybean varieties |
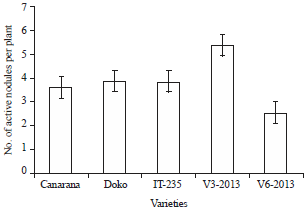 | |
| Fig. 7: | Evaluation of the number of active nodules per plant |
DISCUSSION
The different soybean varieties evaluated performed differently in the shade house and in field for some of the parameters evaluated. There was a higher germination percentage in the shade house than the seed emergence in the field. This difference could be explained by the poor seed quality and the crop environment. Indeed, the quality of a seed is determined by its ability to germinate and its ability to produce a seedling in the field. The decrease in germination seed vigor may be due to the duration and conditions of storage from production to the following growing season. According to Gazzoni (1995), the duration and seed storage conditions affect seed germination and germination vigor. This is due to sudden temperature changes which the seeds could have been exposed to. The acidic pH of the soil from both plots could also have affected seed germination. The process of seed germination and revegetation of saline environments were studied by Maas and Hoffman (1977) who found that the salinity level affected the germination of peas (Cicer arietum). According to Garcia (1995), the formation of regularly distributed plants is dependent on ambient conditions and characteristics of the area sown and that the seed vigor. The seeds of Doko were classified as good quality seeds after the test. However, the seed vigor was found to deteriorate rapidly compared to its germination capacity. In addition, the evaluation of the growth rate showed differences in plant height in the different varieties, which was less than 50 cm.
The soybean varieties were found to produce differences in nodulation. The presence of nodules on the plants of inoculated plots shows that varieties responded positively to inoculation. At 35 DAS, there was no difference in the color of leaves of inoculated and non-inoculated plants. The color differences between the leaves inoculated and non-inoculated plants were observed about 70 DAS. This difference was observed at 60 DAS for Doko variety (N'Gbesso et al., 2010). This color differences could be explained by the inoculation effect, which increases nitrogen fixation by the exhibition of dark green color of the leaves of inoculated plants.
The presence of nodules in non-inoculated plants of V3-2013 variety could indicate the presence of indigenous B. japonicum strains. Another possible explanation could be the result of a contamination of non-inoculated plots by the bacteria as mentioned by Beugre et al. (2013). The low presence of nodule on the non-inoculated treatments of Doko shows that this variety does not spontaneously nodulate. It is described as a variety with strict nodulation (N'Gbesso et al., 2010). It is known that the absence or low amount of efficient strains would be a limiting factor for the establishment of symbiotic bacteria-legume.
According to Streeter (1994) and Slattery et al. (2004), the approach of using effective or superior exotic rhizobia strains as inoculants has failed in various environments due to various reasons including the use of ineffective and non-competitive rhizobia strains as inoculants. The varieties Doko, IT-235 and V6-2013 are varieties that responded only to inoculation. The legume-rhizobial symbiosis has a large impact on the success of legumes hence the atmospheric nitrogen fixed can be more than the fertilizer nitrogen that farmer can afford to buy and apply (Kahindi et al., 1997). Significant correlations observed between nodulation and growth parameters revealed ability to improve the vegetative development and the nodulation of soybean varieties.
CONCLUSION
The results of this study showed that inoculating the seeds contributed to the increasing of the number of nodules per plant of the soybean varieties. However, there is need to repeat this trial to confirm the findings and to determine the best time in the year to plant soybean. Evaluation of other promising varieties for the Nawa region of Côte d’Ivoire should be considered in the future works.
ACKNOWLEDGMENTS
The authors are grateful to ICRAF Côte d’Ivoire Country Program and Cocoa Livelihood Program (CLP2) for the financial support.
REFERENCES
- Abana, P.C., I.J. Ogoke and M.O. Offor, 2015. Evaluation of selected soybean varieties on growth and yield in a humid tropical environment. Res. J. Agric., 2: 1-11.
Direct Link - Berrada, H. and K. Fikri-Benbrahim, 2014. Taxonomy of the Rhizobia: Current perspectives. Br. Microbiol. Res. J., 4: 616-639.
CrossRefDirect Link - Beugre, F.G., P.D.F.M. N'Gbesso, J.T. Ama-Abina and R.G. Yoro, 2013. Influence d'un herbicide et de l'inoculation des semences sur la croissance du soja [Glycine max (L.) Merrill] cultive sur un sol gravillonnaire de plateau. Agronomie Africaine, 25: 221-229.
Direct Link - Bognonkpe, J.P. and M. Becker, 2009. Native soil N mineralization in major rice based cropping systems. J. Anim. Plant Sci., 4: 384-398.
Direct Link - Danso, S.K.A., 1992. Biological Nitrogen Fixation in Tropical Agrosystems: Twenty Years of Biological Nitrogen Fixation Research in Africa. In: Biological Nitrogen Fixation and Sustainability of Tropical Agriculture, Mulongoy, K., M. Gueye and D.S.C. Spencer (Eds.). IITA, Ibadan, Nigeria, ISBN: 9780471935605, pp: 3-13.
- Freitas, H., M.N.V. Prasad and J. Pratas, 2004. Plant community tolerant to trace elements growing on the degraded soils of Sao Domingos mine in the south east of Portugal: Environmental implications. Environ. Int., 30: 65-72.
CrossRefDirect Link - Kahindi, J.H.P., P. Woomer, T. George, F.M. de Souza Moreira, N.K. Karanja and K.E. Giller, 1997. Agricultural intensification, soil biodiversity and ecosystem function in the tropics: The role of nitrogen-fixing bacteria. Applied Soil Ecol., 6: 55-76.
CrossRefDirect Link - King, C.A. and L.C. Purcell, 2001. Soybean nodule size and relationship to nitrogen fixation response to water deficit. Crop Sci., 41: 1099-1107.
CrossRefDirect Link - Lee, H.I., J.H. Lee, K.H. Park, D. Sangurdekar and W.S. Chang, 2012. Effect of soybean coumestrol on Bradyrhizobium japonicum nodulation ability, biofilm formation and transcriptional profile. Applied Environ. Microbiol., 78: 2896-2903.
CrossRefDirect Link - Maas, E.V. and G.J. Hoffman, 1977. Crop salt tolerance-current assessment. J. Irrig. Drainage Div., 103: 115-134.
Direct Link - Matiru, V.N. and F.D. Dakora, 2004. Potential use of rhizobial bacteria as promoters of plant growth for increased yield in landraces of African cereal crops. Afr. J. Biotechnol., 3: 1-7.
Direct Link - Nhamo, N., W. Mupangwa, S. Siziba, T. Gatsi and D. Chikazunga, 2003. The Role of Cowpea (Vigna Unguiculata) and other Grain Legumes in the Management of Soil Fertility in the Smallholder Farming Sector of Zimbabwe. In: Grain Legumes and Green Manures for Soil Fertility in Southern Africa: Taking Stock of Progress, Waddington, S.R. (Ed.). CIMMYT, Mexico, ISBN: 9789706481139, pp: 119-127.
- N'Gbesso, M.F.P., A.S.P. N'Guetta, N.C. Kouame and K. Foua Bi, 2010. [Monitoring the effectiveness of the inoculation with bacteria (Bradyrhizobium japonicum) for 11 genotypes of soybean (Glycine max L. Merrill) in savannah area of Côte d'Ivoire]. Sci. Nat., 7: 59-67, (In French).
Direct Link - N'Zi, J.C., A.P. Koua, K.D. Kouassi, J. Kahia, J.L. Kouassi, A.S.P. N'Guetta and C. Kouame, 2015. Effect of inoculating seeds with Bradyrhizobium japonicum on the agronomic performance of five varieties of soybean (Glycine max) in Côte d'Ivoire. Afr. J. Agric. Res., 10: 3671-3677.
CrossRefDirect Link - Phongpan, S. and A.R. Mosier, 2003. Effect of crop residue management on nitrogen dynamics and balance in a lowland rice cropping system. Nutr. Cycling Agroecosyst., 66: 133-142.
CrossRefDirect Link - Roychowdhury, R., U. Banerjee., S. Sofkova and J. Tah, 2013. Organic farming for crop improvement and sustainable agriculture in the era of climate change. OnLine J. Biol. Sci., 13: 50-65.
CrossRefDirect Link - Simon, Z., K. Mtei, A. Gessesse and P.A. Ndakidemi, 2014. Isolation and characterization of nitrogen fixing Rhizobia from cultivated and uncultivated soils of Northern Tanzania. Am. J. Plant Sci., 5: 4050-4067.
CrossRefDirect Link - Slattery, J.F., D.J. Pearce and W.J. Slattery, 2004. Effects of resident rhizobial communities and soil type on the effective nodulation of pulse legumes. Soil Biol. Biochem., 36: 1339-1346.
CrossRefDirect Link - Stahl, L., G. Nyberg, P. Hogberg and R.J. Buresh, 2002. Effects of planted tree fallows on soil nitrogen dynamics, above-ground and root biomass, N2-fixation and subsequent maize crop productivity in Kenya. Plant Soil, 243: 103-117.
CrossRefDirect Link - Stajkovic, O., D. Delic, D. Josic, D. Kuzmanovic, N. Rasulic and J. Knezevic-Vukcevic, 2011. Improvement of common bean growth by co-inoculation with Rhizobium and plant growth-promoting bacteria. Romanian Biotechnol. Lett., 16: 5919-5926.
Direct Link - Streeter, J.G., 1994. Failure of inoculant rhizobia to overcome the dominance of indigenous strains for nodule formation. Can. J. Microbiol., 40: 513-522.
CrossRefDirect Link - Wong, M.H., 2003. Ecological restoration of mine degraded soils, with emphasis on metal contaminated soils. Chemosphere, 50: 775-780.
CrossRefDirect Link - Bekere, W. and A. Hailemariam, 2012. Influences of inoculation methods and phosphorus levels on nitrogen fixation attributes and yield of soybean (Glycine max L.) at Haru, Western Ethiopia. Am. J. Plant Nutr. Fertilizat. Technol., 2: 45-55.
CrossRefDirect Link