
Wei Li
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Chengdu, Sichuan 611830, China
An-Jun Liu
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Chengdu, Sichuan 611830, China
Yu-Zhen Sheng
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Chengdu, Sichuan 611830, China
Guo-Yue Cheng
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Chengdu, Sichuan 611830, China
Zi-En Pu
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Chengdu, Sichuan 611830, China
Ya-Xi Liu
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Chengdu, Sichuan 611830, China
Asian Journal of Plant Sciences
Year: 2013 | Volume: 12 | Issue: 1 | Page No.: 34-39







ABSTRACT
In order to exploiting new genetic resources for the wheat breeding, nine waxy genes (granule-bound starch synthase, GBSSI) were obtained from wild species, Aegilops tauschii (DD) by special primer PCR cloning. These sequences showed the higher similarity to the sequences of Wx-D1 locus in common wheat. The analysis of variation and alignment of sequences showed that all the sequences could be distinguished, although from the same species. Out of 35 variable sites of nucleotide in the whole sequences, twelve variable sites located on the region of exon and made six amino acid residues change. Most of variable sites located on transit peptide. Using neighbour-joining method, phylogenetic tree suggested that the sequences of waxy gene from Ae. tauschii were clustered closely with the sequences of Wx-D1 locus and far with the sequence of Wx-A1 and Waxy-B1 loci in Triticum L.. Moreover, Ae. tauschii showed more close relationship with Triticum L. and barley and had more close relationship with barley and far distance with rice, potato, pea and Arabidopsis. These results would contribute to the deep understanding of functional aspects and evolution of waxy gene and the improving of starch quality in common wheat.
PDF Abstract XML References Citation
Received: March 05, 2013;
Accepted: April 15, 2013;
Published: June 21, 2013
How to cite this article
Wei Li, An-Jun Liu, Yu-Zhen Sheng, Guo-Yue Cheng, Zi-En Pu and Ya-Xi Liu, 2013. Molecular Characterization of Waxy Gene in Aegilops tauschii. Asian Journal of Plant Sciences, 12: 34-39.
DOI: 10.3923/ajps.2013.34.39
URL: https://scialert.net/abstract/?doi=ajps.2013.34.39
DOI: 10.3923/ajps.2013.34.39
URL: https://scialert.net/abstract/?doi=ajps.2013.34.39
INTRODUCTION
Aegilops tauschii (DD, 2n = 14) is considered as not only the ancestor of D genome of hexaploid wheat (McFadden and Sears, 1946; Kihara, 1944), but also the first gene pool for the improving of common wheat (Kimber and Feldman, 1987). This wild diploid species contains some excellent potential traits and easily transfer these characters into tetraploid and hexaploid wheat (Dudnikov, 1998). The original location of this species is the west of Asian. The waxy gene on the D genome had been cloned in common wheat (Yan et al., 2000; Murai et al., 1999). Granule-bound starch synthase (GBSSI), also called WAXY protein (Echt and Schwartz, 1981), is a key enzyme in the process of amylose synthesis and play an important role in the starch synthesis of cereal (Nakamura et al., 1993). The molecular weights of three subunits of GBSSI protein are from 59-60 kDa. They are encoded by three loci of waxy gene, Waxy-A1 (Wx-A1, 2,781 bp), Waxy-B1 (Wx-B1, 2,794 bp) and Waxy-D1 (Wx-D1, 2, 862 bp), each consisting of 11 exons and 10 introns, located on chromosome 7AS, 4AL and 7DS, respectively (Nakamura et al., 1993; Murai et al., 1999). The waxy gene had been cloned from barely (Rohde et al., 1988), rice (Hirano and Sano, 1991; Wang et al., 1990), maize (Shure et al., 1983; Klosgen et al., 1986), sorghum (Hamblin et al., 2007), pea (Dry et al., 1992), potato (Van-Der-Leij et al., 1991), common wheat (Mason-Gamer et al.,1998; Murai et al., 1999).
The variation of amylose content is associated with the status of the waxy gene expression (Yamamori et al., 1994; Urbano et al., 2002). The molecular mechanism of variation was also continuously focused for waxy gene researches. There are a 19 bp deletion at an exon-intron junction and a deletion including the entire coding region of the Wx-B1 gene and a 588 bp deletion in the C-terminal region at Wx-D1 locus in Japanese bread wheat Kanto 107 and Chinese cultivar Baihuo (Vrinten and Nakamura, 2000). A 173 bp insertion of waxy gene also led to a new null Wx-A1 allele in six Turkish cultivars (Saito et al., 2004). Many SNPs (single nucleotide polymorphisms) and/or InDels (insertions or deletions) were detected in the introns of waxy gene (Huang and Brule-Babel, 2010). Consequently, although waxy is usually considered as conserved gene, the variation of sequences, including exons and introns, could be detected between Triticum and Aegilops (Mason-Gamer et al., 1998).
The variations of waxy gene in wild diploid species, T. monococcum L. ssp. monococcum and T.urartu Thum. Ex Gandil. which was considered as the A genome origin of the all polyploidy wheats, have been well characterized at the molecular level (Murai et al., 1999; Yan et al., 2000; Liu et al., 2009). But the less information was reported about the performance of waxy gene in Ae. tauschii which was considered as the ancestor of D genome of hexaploid wheat. It was blocking to understanding deeply the function and evolution of waxy gene in Triticum L. and exploiting the new genetic resources.
In present study, the variation of sequence of waxy gene was investigated in Ae. tauschii which potentially could be an important source for improving starch quality and also provide valuable information on the phylogeny of waxy gene in different species.
MATERIALS AND METHODS
A total of nine accessions of Ae. tauschii, were used in this study (Table 1). The materials were kindly provided by Dr. Harold Bockelaman, USDA-ARS, National Small Grains Collection.
All seeds were germinated under the dark at 23°C for 1 week, young leaves were harvested and crushed into powder with the liquid nitrogen and the genomic DNA extracted by the CTAB method (Wang et al., 2008). A pair of primers which were designed by the conserve sequence region of waxy genes (Yan et al., 2000), was used as the cloning primers: the forward primer: 5’-TTGCTGCAGGTAGCCACAC-3’ and the reverse primer: 5’-CTC AAGTGCTGCCTGGCAGAGAA-3’. The amplified region includes the first, second and the third exons and introns and also includes the partial fourth exon of waxy gene. PCR was performed in a 50 μL volume, containing 1.5 UTaq plus DNA polymerase, 100 ng of each template DNA, 5 μL PCR buffer (supplied with Taq plus DNApolymerase), 1.5 mM MgCl2, 100 mM of each dNTP and 150 ng each primer. The reactions were conducted in a PTC-220 (MJ Research, USA) using the following program: 95°C for 3 min, followed by 35 cycles at 94°C for 1 min, at 60°C for 1 min and at 72°C for 2 min and a final extension step of 72°C for 10 min. The PCR products were separated on 1.5% agarose gels. The expected fragments were recovered and cloned into pMD18-T vector (TaKaRa), then transformed into the competent E. coli cells (DH5α).
| Table 1: | The information of the cloning waxy genes in this study |
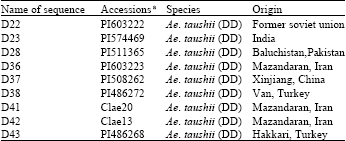 | |
| a: No. of accessions came from USDA-ARS, National Small Grains Collection (http://www.ars-grin.gov/npgs/index.html) | |
Positive clone of each accession was sequenced by commercial company (Invitrogen) in two directions.
Data analysis: The obtained sequences were compared with known waxy sequences using BLAST (http://www.ncbi.nlm.nih.gov/BLAST). Sequences were manually inspected by BioEdit ver 7.0.9 (Hall, 1999). Sequence alignment was completed by ClustalX (Larkin et al., 2007) and DNAMAN 5.2.2 (http://www.lynnon.com). Neighbour-joining trees was constructed based on the alignment of all known sequences, including six sequences from barley (Rohde et al., 1988), rice (Wang et al., 1990), Pea (Dry et al., 1992), potato (Van-Der-Leij et al., 1991) and Arabidopsis (NM103023), respectively, using Kimura 2-parameter model in MEGA4.0 (Saitou and Nei, 1987; Kimura 1980; Tamura et al., 2007).
RESULTS
Variation of sequences of waxy gene: A total of nice sequences with 925 bp were obtained after cloning from all the materials. Blast in NCBI confirmed all the sequences belonged to the partial waxy gene, suggested that the waxy gene was successfully cloned from Ae. tauschii (Table 1). The average value of similarity between the sequences was 99.12% and showed 98.85% and 98.76% similar with Wx-TtD (AF110375) and Wx-D1a (AB019624). The substitution and indels of nuclear bases were detected at thirty-five base sites in the whole sequences. The substitution types included A-G, T-C, A-C, G-C and G-T. The indels types mainly focused on one or two bases deletion.
Based on the variation of bases, all the sequences could be clustered into four groups. There was difference only at one SNP site between sequences from the same group, likes group 1 (D23 and D43), group 2 (D37 and D38) and group 3 (D36 and D41), indicated that these sequences have the higher similarity each other. Furthermore, sequences from group 4 showed the highest variation, compared with other groups. Group 3 consisted of the sequences from accession PI603223 and clase 20 which came from Mazandaran, Iran. Interestingly, the accession clase 13 which also came from the Mazandaran, Iran, was clustered into group 4 (D22, D28 and D42). These results indicated although the accessions came from the same region, the variation of sequence of waxy gene could be observed in the same species, Ae. stauschii.
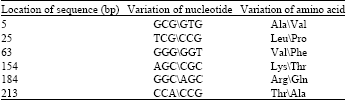
Variation of amino acid residue of waxy gene: All the sequences included three introns and four exons. The lengths of the first, second, third and fourth exon were 321, 81, 99 and 102 bp while the first, second and third intron were 90, 96 and 104 bp, respectively. Twelve variable nucleotide sites were observed in the region of exons, but only six sites lead the change of amino acid residue among the sequences waxy gene. Two types of amino acid residues was detected at 5 bp (Ala\Val), 25 bp (Leu\Pro), 63 bp (Val\Phe), 154 bp (Lys\Thr), 184 bp (Arg\Gln) and 213 bp (Thr\Ala), respectively (Table 2). Interestingly, most of variable amino acid residues located on the transit peptide of WAXY protein (Fig. 1). The change of nucleotide mainly happened on the first and third base in the codon.
| Table 2: | The distribution of variable amino acid residues of waxy gene in Ae. tauschii |
 | |
Phylogenetic analysis: In order to estimate the relationship of waxy gene between Ae. tauschii and other species, the phylogenetic tree was constructed by Neighbor-joining method (Fig. 2). A total of 21 sequences were used in this tree, including sequences from barely, rice, potato, pea and Arabidopsis, respectively. All the sequences could be clustered to six groups. All sequences from Ae. tauschii and sequence AF110375 from Ae. tauschii and sequence AF163319 from Wx-D1 locus of common wheat, were clustered into one group, suggested that waxy gene from D genome showed higher conserved characterization. Interestingly, sequence D28, D39 and D22 which were collected from different location, were clustered closely. Sequence of waxy gene from potato and rice were clustered into one group while sequences from Arabidopsis and pea were clustered into one group. Sequence from barely was more similar to sequence of waxy gene from Triticum L. and Aegilops species. Sequence from Ae. speltoides (AF110374), T. monococcum (AF110373), common wheat (EU719660 and AB019622), suggested that waxy gene at Wx-B1 locus showed more diversity in Triticum L. and Aegilops species.
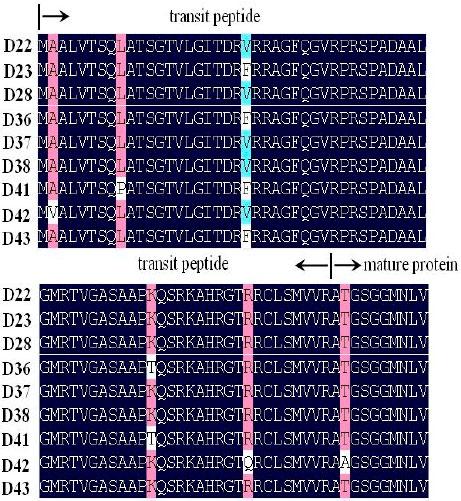 | |
| Fig. 1: | Partial alignment of amino acids residues of waxy gene. Conservative positions are indicated by dark shading. Names of sequences were referenced in Table 1 |
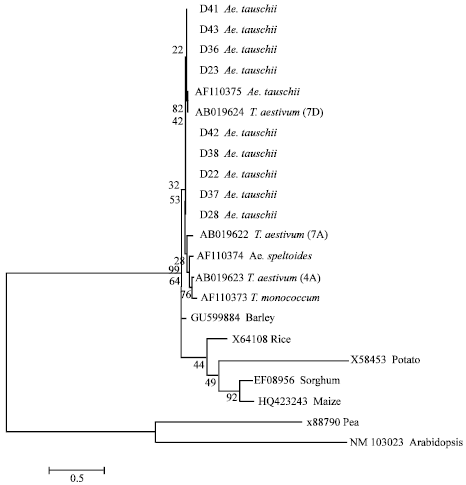 | |
| Fig. 2: | Neighbour-joining tree based on sequences of waxy gene was constructed by using Kimura 2-parameter model in MEGA 4.0. Bootstrap tree was obtained by 1, 500 repeat calculation |
Interestingly, sequence EU719608 (Wx-1Aa) was clearly distinguished with other sequences. Otherwise, the sequence of waxy gene from Ae. tauschii had the close relationship with the sequence at Wx-D1 locus and far relationship with the sequence at Wx-A1 and Wx-B1 from Triticum L.
DISCUSSION
Waxy gene encodes from 59-60 kDa protein and is a key regulate gene of the synthesizing process of amylose which is one of main components of seed endosperm (Yamamori et al., 1994). Alternatively, Ae. tauschii, the donor of D genome in common wheat, is one of the most important gene resources for the improving of common wheat, as it is easy to transform gene to common wheat by directly cross (Kimber and Feldman, 1987).Whereas, exploiting allele of waxy gene in Ae. tauschii could provide more useful genetic resources and understand the function and evolution of waxy gene. In the previous studies, null mutation at Wx-D1 locus (Wx-D1b) could lead to the distinct decreasing of amylose content in Ae. tauschii (Ainsworth et al., 1993). Yan et al. (2000) obtained the sequences of waxy gene from diploid species containing genome A, B and D (Wx-TmA, Wx-TsB and Wx-TtD), 2803, 2795 and 2862 bp and Murai et al. (1999) cloned the sequence of Wx-D1 locus (2862 bp), containing 11 exons and 10 introns. McLauchlan et al. (2001) developed five PCR-molecular markers based on the alignment of sequences from emmer wheat (2n = 14, AA), Ae. tauschii (2n = 14, DD), barely, maize, rice, potato. Briney et al. (1998) compared the gDNA sequences from rice and barely and sequences of mRNA from wheat, barley and rice and design the STS-PCR primers for Wx-B1 locus. Fujita et al. (1996) thought that there was conserve at the N terminal of Wx-D1 subunit between common wheat (AABBDD) and Ae. tauschii. Although waxy gene are considered as high conserved sequences in Triticum L., in the present study, the sequences of waxy gene from Ae. tauschii still showed variation of nucleotide bases, even changed the amino acid residues at six sites in the expressing region. The effect of variation of amino acid residues, especially on transit peptide region, for the function of waxy gene, or amylose content and starch quality, should be further investigated. It could help to exploit unique gene resources from Ae. tauschii for the improving of common wheat.
Although, the alignment of waxy gene from diploid wheats showed very less variation, the phylogenetic tree can distinguished the sequences of T. urartu, T. boeoticum and T. monococcum (Liu et al., 2009). In the present study, the sequences of waxy gene from Ae. tauschii were clustered closely with sequences at Wx-D1 locus in common wheat and were far with the sequences at Wx-A1 and Wx-B1 loci in Triticum L.. Thereby, although amino acid sequences and structural motifs are highly conserved, the sequences of waxy gene can showed the specificity of genomes not only at Wx-A1, Wx-B1 and Wx-D1 loci of hexaploid wheat, but also in diploid wheats which is usually considered as the ancestor of A genome and Ae. speltoides which is usually considered as the ancestor of B genome and Ae. tauschii which is usually considered as the donor of D genome. The results showed that the variation of exon was enough to provide extensive resolution within very closely related species. It is also similar to the results reported by Mason-Gamer et al. (1998). Many SNPs and/or InDels (insertions or deletions) have been observed in the regions of intron (Huang and Brule-Babel, 2010), also provide an approach to analysis the genetic diversity and exploit the molecular markers of waxy gene. The new variant sites of amino acid residuces of waxy gene, compared with the previous report (Fujita et al., 1996), suggested that exploiting the variation of waxy gene in new genetic resources is necessary to understanding the genetic diversity and function of waxy gene.
CONCLUSION
In this study, nine waxy genes were cloned from the donor of D genome of common wheat, Aegilops tauschii (DD), by special primer PCR cloning. These sequences were not only higher similar to the gene of Wx-D1 locus in common wheat, but also showed the different variation each other, although in the same species. Out of 35 variable sites of nucleotide in the whole sequences, twelve variable sites located on the region of exon and made six amino acid residues change. Most of variable sites located on transit peptide. Based on the neighbour-joining method, phylogenetic tree also showed that the sequences of waxy gene from Ae. tauschii were clustered closely with the sequences of Wx-D1 locus and far with the sequence of Wx-A1 and Waxy-B1 loci in Triticum L. Moreover, Ae. tauschii showed more close relationship with Triticum L. and barley and had more close relationship with barley and far distance with rice, potato, pea and Arabidopsis. The present results would contribute to the deep understanding of functional aspects and evolution of waxy gene in Triticum L. In the future, the further research for the quality effect of these genes could help to improving starch quality of common wheat.
ACKNOWLEDGMENT
This study was supported by the National Basic Research Program of China (973 Program and 2011CB100100) and the Key Project of National Natural Science Foundation of China (31230053).
REFERENCES
- Ainsworth, C.C., J. Clark and J. Balsdon, 1993. Expression, organization and structure of the genes encoding the waxy protein (granule-bound starch synthase) in wheat. Plant Mol. Biol., 22: 67-82.
PubMedDirect Link - Briney, A., R. Wilson, R.H. Potter, I. Barclay, G. Crosbie, R. Appels and M.G.K. Jones, 1998. A PCR-based marker for selection of starch and potential noodle quality in wheat. Mol. Breeding, 4: 427-433.
CrossRefDirect Link - Dry, I., A. Smith, A. Edwards, M. Bhattacharyya, P. Dunn and C. Martin, 1992. Characterization of cDNAs encoding two isoforms of granule-bound starch synthase with differential expression in developing storage organs of pea and potato. Plant J., 2: 193-202.
PubMedDirect Link - Dudnikov, A.J., 1998. Allozyme variation in Transcaucasian populations of Aegilops squarrosa. Heredity, 80: 248-258.
CrossRefDirect Link - Echt, C.S. and D. Schwartz, 1981. Evidence for the inclusion of controlling elements within the structural gene at the Waxy locus in maize. Genetics, 99: 275-284.
Direct Link - Fujita, N., A. Wadnao, S. Kozaki, K. Takaoka, S. Okabe and T. Taira, 1996. Comparison of the primary structure of waxy proteins (granule-bound starch synthase) between polyploid wheats and related diploid species. Biochem. Genet., 34: 403-413.
CrossRefDirect Link - Hall, T.A., 1999. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acid Symp. Ser., 41: 95-98.
Direct Link - Hamblin, M.T., M.G.S. Fernandez, M.R. Tuinstra, W.L. Rooney and S. Kresovich, 2007. Sequence variation at candidate loci in the starch metabolism pathway in Sorghum bicolor: Prospects for LD mapping. Crop Sci., 47: 125-134.
Direct Link - Hirano, H. and Y. Sano, 1991. Molecular characterization of the waxy locus of rice (Oryza sativa). Plant Cell Physiol., 32: 989-997.
Direct Link - Huang, X.Q. and A. Brule-Babel,. Development of genome specific primers for homoeologous genes in allopolyploid species: The waxy and starch synthase II genes in allo-hexaploid wheat (Triticum aestivum L.) as examples. BMC Res. Notes, Vol. 3.
CrossRefDirect Link - Kimura, M., 1980. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol., 16: 111-120.
CrossRefDirect Link - Klosgen, R.B., A. Gierl, Z.S. Schwarz-Sommer and H. Saedler, 1986. Molecular analysis of the waxy locus of Zea mays. Mol. Gen. Genet., 203: 237-244.
CrossRef - Larkin, M.A., G. Blackshields, N.P. Brown, R. Chenna and P.A. McGettigan et al., 2007. Clustal W and clustal X version 2.0. Bioinformatics, 23: 2947-2948.
CrossRefPubMedDirect Link - Liu, Y.X., W. Li, Y.M. Wei, G.Y. Chen and Y.L. Zheng, 2009. Molecular characterization of the Waxy gene in einkorn wheat. J. Plant Sciences, 4: 114-121.
CrossRefDirect Link - Mason-Gamer, R.J., R.J. Weil and E.A. Kellogg, 1998. Granulebound starch synthase: Structure, function and phylogenetic utility. Mol. Biol. Evol., 15: 1658-1673.
Direct Link - McFadden, E.S. and E.R. Sears, 1946. The origin of Triticum spelta and its free threshing hexaploid relatives. J. Hered, 37: 81-89.
Direct Link - McLauchlan, A., F.C. Ogbonnaya, B. Hollingsworth, M. Carter and K.R. Gale et al., 2001. Development of robust PCR-based DNA markers for each homoeoallele of granule-bound starch synthase and their application in wheat breeding programs. Aust. J. Agric. Res., 52: 1409-1416.
Direct Link - Murai, J., T. Taira and D. Ohta, 1999. Isolation and characterization of the three waxy genes encoding the granule-bound starch synthase in hexaploid wheat. Genet, 234: 71-79.
CrossRef - Nakamura, T., M. Yamamori, H. Hirano and S. Hidaka, 1993. Identification of three wX proteins in wheat (Triticum aestivum L.). Biochem. Genet, 31: 75-86.
CrossRef - Rohde, W., D. Becker and F. Salamini, 1988. Structural analysis of the waxy locus from Hordeum vulgare. Nucleic Acids Res., 16: 7185-7186.
Direct Link - Saito, M., M. Konda, P. Vrinten, K. Nakamura and T. Nakamura, 2004. Molecular comparison of waxy null alleles in common wheat and identification of a unique null allele. Theor. Applied Genet., 108: 1205-1211.
CrossRefPubMedDirect Link - Saitou, N. and M. Nei, 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol., 4: 406-425.
CrossRefPubMedDirect Link - Shure, M., S. Wessle and N. Fedoroff, 1983. Molecular identification and isolation of the waxy locus in maize. Cell, 35: 225-233.
CrossRefDirect Link - Tamura, K., J. Dudley, M. Nei and S. Kumar, 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol., 24: 1596-1599.
CrossRefPubMedDirect Link - Urbano, M., B. Margiotta, G. Colaprico and D. Lafiandra, 2002. Waxy proteins in diploid, tetraploid and hexaploid wheat. Plant Breeding, 121: 465-469.
CrossRefDirect Link - Van-Der-Leij, F.R., R.G.F. Visser, A.S. Ponstein, E. Jacobsen and W.J. Feenstra, 1991. Sequence of structural gene for granulebound starch synthase of potato (Solanum tuberosum L.). Mol. Gen. Genet., 228: 240-248.
Direct Link - Vrinten, P.L. and T. Nakamura, 2000. Wheat granule-bound starch synthase I and II are encoded by separate genes that are expressed in different tissues. Plant Physiol., 122: 255-264.
PubMedDirect Link - Wang, Z., Z. Wu, Y. Xing, F. Zheng, X. Guo, W. Zhang and M. Hong, 1990. Nucleotide sequence of rice waxy gene. Nucleic Acids Res., 18: 5898-5898.
CrossRefDirect Link - Wang, J.R., Y.M. Wei, X.Y. Long, Z.H. Yan, E. Nevo, B.R. Baum and Y.L. Zheng, 2008. Molecular evolution of dimeric α-amylase inhibitor genes in wild emmer wheat and its ecological association. BMC Evol. Biol., Vol. 8.
CrossRefDirect Link - Yamamori, M., T. Nakamura, T.R. Endo and T. Nagamine, 1994. Waxy protein deficiency and chromosomal location of coding genes in common wheat. Theor. Applied Genet., 89: 179-184.
Direct Link - Yan, L.L., M. Bhave, R. Fairclough, C. Konik, S. Rahman and R. Appels, 2000. The genes encoding granule-bound starch synthases at the waxy loci of the A, B and D progenitors of common wheat. Genome, 43: 264-272.
CrossRefDirect Link