
Palaniappan Selvakumar
Department of Life Sciences, National Chung Hsing University, Taichung 402, Taiwan, Republic of China
Singaram Arvinth
Department of Life Sciences, National Chung Hsing University, Taichung 402, Taiwan, Republic of China
Subbiyan Maruthasalam
Department of Life Sciences, National Chung Hsing University, Taichung 402, Taiwan, Republic of China
Chin Ho Lin
Department of Life Sciences, National Chung Hsing University, Taichung 402, Taiwan, Republic of China
Asian Journal of Plant Sciences
Year: 2013 | Volume: 12 | Issue: 3 | Page No.: 128-136







ABSTRACT
Hydrogen peroxide (H2O2) production is one of the most important plant defense responses to pathogen infection. In the present study, the endogenous H2O2 level of tobacco (Nicotiana tabaccum L. cv. ‘SR1’) plants were enhanced by constitutively expressing a Glucose Oxidase (GO) gene isolated from the fungus Aspergillus niger and the level of disease resistance was observed. An elevated level of H2O2 was confirmed in the GO-transformed tobacco plant leaf tissue. Transgenic tobacco plants exhibited resistance to leaf spot fungal disease and bacterial wilt disease due to increased levels of H2O2 accumulation (ranging from 159±16.78 to 355±13.89 μmol g-1 leaf tissue) when compared to wild tobacco plants (27.37±3.91 μmol g-1 leaf tissue). The increased accumulation of H2O2 in transgenic plants significantly delayed the infection of fungal target spot disease caused by Rhizoctonia solani. The artificial inoculation of R. solani to tobacco plant leaves completely covered the leaf surface (100%) of control wild plants at 7 days post inoculation (DPI), but the transgenic lines were resistant and only 50-70% of the leaf surface was infected 7 DPI. In case of Ralstonia solanacearum (causing bacterial wilt), the control tobacco plants completely wilted (80-100%) at 15 DPI. However, transgenic tobacco plants were significantly resistant to virulence of Ralstonia solanacearum and 30-50% of wilting symptoms appeared after 15 DPI. The level of H2O2 was increased approximately one-fold in infected transgenic tobacco plants relative to uninfected transgenic plants. Thus, the activation of H2O2 level in transgenic plants represents a novel approach for engineering broad-spectrum disease resistance in plants.
PDF Abstract XML References Citation
Received: February 25, 2013;
Accepted: June 28, 2013;
Published: September 19, 2013
How to cite this article
Palaniappan Selvakumar, Singaram Arvinth, Subbiyan Maruthasalam and Chin Ho Lin, 2013. Disease Resistance Conferred by Constitutive Expression of a Fungal Glucose Oxidase Gene in Transgenic Tobacco Plants. Asian Journal of Plant Sciences, 12: 128-136.
DOI: 10.3923/ajps.2013.128.136
URL: https://scialert.net/abstract/?doi=ajps.2013.128.136
DOI: 10.3923/ajps.2013.128.136
URL: https://scialert.net/abstract/?doi=ajps.2013.128.136
INTRODUCTION
Tobacco (Nicotina tabacum L.) is one of the important agricultural products and it has become a model system for past several decades in tissue culture and genetic engineering (Ganapathi et al., 2004). Tobacco plants are sensitive and prone to numerous diseases and almost all disease will cause the some sort of leaf symptoms. Fungi, bacteria and viruses cause most tobacco plant diseases will result in leaf discoloration indirectly (Kucharek, 2001). As long as agriculture exists, farmers have to battle against plant fungal and bacterial pathogens which infect crop plants and thereby cause considerable losses of yield and quality (Honee, 1999). The control measures to the disease varies viz., chemical control with fungicides (Nel et al., 2003), modification of plants to obtain improved genetic capabilities through plant breeding and by integrating foreign DNA into plant genomes to produce transgenic plants (Zupan and Zambryski, 1995; Hwang and Ko, 2004; Barampuram and Zhang, 2011). However, the use of broad-spectrum fungicides further results in imbalances within the microbial community creating unfavorable conditions for the activity of beneficial organisms (Villajuan et al., 1996). Genetic engineering technology used to activate the inherent plant responses against a pathogen by either using single foreign disease resistant gene (Keen, 1999) or by choosing the plant genes to trigger the existing defense mechanisms (Bent and Yu, 1999; Rommens and Kishore, 2000).
Plant produced vast array of proteins and other organic molecules against plant pathogens prior to infection or during pathogen attack with their own defense mechanism. The formation of necrotic lesions in the infection site is associated with the coordinated induction of diverse defense responses such as cell wall reinforcement, synthesis of phytoalexins and oxidation of phenolic compounds, activation of defense-related genes and localization of cell death or the hypersensitive response (Amil-Ruiz et al., 2011). Activation of defense related genes extends systemically to the whole plant in which Salicylic Acid (SA) is a key signaling molecule, conferring broad spectrum resistance to the pathogen. This process is called Systemic Acquired Resistance (SAR) (Lee et al., 2002). One of the most peculiar responses in plant-pathogen interactions is a rapid and transient production of Active Oxygen Species (AOS), such as the superoxide anion radical (O2¯), hydroxyl radical (OH’) and hydrogen peroxide (H2O2). This process is referred to as an oxidative burst (Gechev et al., 2006; Petrov and Van Breusegem, 2012). The accumulation of H2O2 has been determined to be one of the earliest events that occur at host-pathogen recognition; it has been postulated to play an important role in plant defense (Houot et al., 2001). In addition to its oxidative potential in killing or inhibiting the growth of pathogens, H2O2 has been shown to be involved in a number of plant defense response processes. In plant cell suspension culture systems, H2O2 produced during the oxidative burst has been shown to activate the biosynthesis of phytoalexins and to induce the cross-linking of the hydroxyproline- rich cell wall glycoproteins (Miller et al., 2010; Petrov and Van Breusegem, 2012).
For controlling the plant diseases several strategies have been developed, among them, development of transgenic plants with Glucose Oxidase (GO; EC. 1.1.3.4) gene is playing an important role. GO is closely related to plant diseases resistance and well documented. It catalyzes the oxidation of β-D-glucose to D glucono-1, 5-lactone and H2O2, whose production is toxic to phytopathogens (Kim et al., 1988). A number of bacteria and fungi produce GO which has a putative antibiotic function, but GO has not been found in animals and plants (Frederick et al., 1990). This property of GO provides a new strategy for the development of disease-resistant plants (Wu et al., 1995; Kazan et al., 1998; Murray et al., 1999). Earlier reports showed GO transformed transgenic plants enhanced disease resistance to a wide range of pathogens, viz., Erwinia carotovora subsp. carotovora, Phytophthora infestans and Verticillium dahlia in transgenic potato (Wu et al., 1995), Rhizoctonia solani in transgenic tobacco (Murray et al., 1999) and Verticillium dahliae in transgenic cotton (Murray et al., 1999). However, some harmful phytotoxic effects, such as sterility, reduced germination rate, root growth inhibition, necrotic lesions and senescence under strong illumination were reported when GO from Talaromyces flavus was expressed in transgenic plants (Kazan et al., 1998; Murray et al., 1999). In contrast to these reports, transgenic potato expressing GO from Aspergillus niger showed normal growth (Wu et al., 1995). Expression of GO gene in transgenic plants with different constitutive promoter might be resulted in greater disease resistance (Felcher et al., 2003) and over expression of GO could enhance resistance to bacterial and fungal pathogens (Hilbeck et al., 2008). In addition, apart from the biotic stress, Maruthasalam et al. (2010) reported that the transgenic tobacco plants expressing GO from A. niger expressed higher amount of tolerance to low temperature than non GO transformed control plants. Based on the above information clearly understand that transgenic plants expressing GO from A. niger will be more resistant to biotic and abiotic stress.
A previous study from this laboratory proved the gene encoding GO from A. niger was expressed in tobacco plants were more resistant to chilling tolerance due to elevated production of H2O2 (Maruthasalam et al., 2010). In the present study, aim to observe the disease resistance level of transgenic tobacco plants having the gene encoding GO from A. niger. The increased level of H2O2 in transgenic tobacco plants resistance against important fungal pathogen Rhizoctonia solani causing target spot disease to tobacco and bacterial pathogen Ralstonia solanacearum causing wilt disease to tobacco plants were studied.
MATERIALS AND METHODS
Plant material and plant growth conditions: Transgenic lines of tobacco were produced with GO gene from A. niger var. niger under the control of the Cauliflower mosaic virus (CaMV) 35S constitutive promoter and nopaline synthase (NOS) terminator (Maruthasalam et al., 2010). All nontransgenic and transgenic plants were propagated through seeds and maintained in growth chambers with a 16/8-h light/dark cycles at 24°C with periodical watering and nutrient supplementation. As previously reported by Maruthasalam et al. (2010) transgenic tobacco lines viz., SR1, T1, T4-7 were taken up for the in vivo study and these transgenic lines accumulated GO protein and had increased levels of H2O2 in the leaf tissue and root.
Determination of specific GO activity: Specific GO activity was determined according to Maruthasalam et al. (2010). Briefly, 1.0 g of freshly collected leaves was ground with liquid nitrogen, homogenized in 0.1 M citrate phosphate buffer (pH 5.0) and the protein content of supernatant was determined. To ~100 μg of total protein, a reaction buffer containing 1.7 mM potassium iodide (KI) and 80 mM glucose was added to a final volume of 1.0 mL and incubated at 37°C for 20 min. The reaction was stopped with 200 μL of H2SO4 and 200 μL of freshly prepared 1.0% starch solution containing 0.83 mM sodium molybdate was added to facilitate color development. The absorbance was read at 540 nm after 10 min. The GO activity was calculated using a calibration curve of purified GO protein (Sigma, USA). One unit of GO activity represents the amount that can oxidize 1.0 l M of β-D-glucose into D-gluconic acid and H2O2 per minute at pH 5.1 and 35°C (Lee et al., 2002).
Qualitative detection of H2O2 in transgenic plants: Histochemical analyses for H2O2 production were studied using starch-KI (Potassium Iodide) medium (Olson and Varner, 1993; Murray et al., 1999) and diaminobenzidine (DAB) uptake method (Thordal-Christensen et al., 1997; Kang et al., 2003). In starch-KI test, leaf discs collected from the tobacco lines were incubated on a solid medium (4% starch, 0.1 M KI, 1.0 M glucose, 0.8% agarose) for 4-5 h at room temperature for colour development (Maruthasalam et al., 2010).
Colorimetric estimation of H2O2 content: H2O2 content in tobacco leaves was determined according to Maruthasalam et al. (2010). About 500 mg of leaf tissue was extracted under ice-cold conditions with 0.1% trichloroacetic acid (TCA). After centrifugation (12,000 rpm/4°C/20 min; Sigma MK-202, Germany), 0.5 mL leaf extract was mixed with 0.5 mL of 100 mM potassium phosphate buffer (pH 7.0) and 1 mL of 1 M KI. The reaction mixture was left in darkness for 1 h and the H2O2 content was calculated by measuring the absorbance at 410 nm and comparing it to a standard graph of known amounts of H2O2 (Merck, Germany).
Fungal target spot disease resistance test: For the live plant leaf infection test, seeds of five homozygous transgenic lines T1, T4, T5, T6 and T7 were germinated on MS medium with hygromycin (25 mg L-1) and maintained in pots as described earlier. Non-transformed SR1 plants were established similarly on the hygromycin-free medium. Four weeks old tobacco plants were used for live plant leaf infection test. All the R. solani inoculations were carried out with mycelial discs of 5 mM diameter, obtained from 4-day-old cultures of R. solani, grown on potato dextrose agar, at 28°C. Each selected leaf was inoculated with a single mycelial disc to the center of the abaxial leaf surface. The mycelial disc placed leaf was covered with absorbent cotton and secured with polypropylene bag. The cotton was moistened periodically with sterile distilled water, to maintain high humidity. Development of symptoms was recorded 3 Days after Inoculation (DAI) and area of infection was measured every day.
Bacterial wilt disease resistant test: The virulence of Ralstonia solanacearum strains was tested on six week old transgenic tobacco plants which were grown in 150 g (dry weight) of organic mixture (8:1:1 ratio of peat moss: vermiculite: perlite). Bacteria from freshly streaked plates were grown overnight in Casamino acid-Peptone-Glucose (CPG) broth (Casamino acid 1 g L-1, Peptone 10 g L-1 and Glucose 5 g L-1) centrifuged to pellet the cells and resuspended in sterile water. Tobacco roots were wounded by cutting vertically down across each pot 1 cm away from the base of the stem. Sixty milliliters of a water suspension of washed bacteria (each with OD600 = 1.0) was poured over the dry soil (150 g) to give initial concentrations of 3 to 5x108 CFU g-1 of soil of tobacco plant per pots. For each assay, inoculum concentrations were determined by dilution plating. Plants were inspected daily for signs of wilting and were rated on a zero-to-four disease index scale as follows: 0, no wilting (plant healthy); 1, 1 to 25% wilting (one leaf wilted); 2, 26 to 50% wilting (up to one-third of plant wilted); 3, 51 to 75% wilting (up to two-third of plant wilted) and 4, 76 to 100% wilted or dead (plant completely wilted). Each assay contained 40 (five plants per each line) plants per treatment and was repeated four times. Disease index was calculated as follows:
where, a, b, c, d, e are the number of plants in 5 categories, respectively and t is the total number of plants examined.
Statistical analysis: The data were subjected to One-way Analysis of Variance (ANOVA) to determine the significance of individual differences at p<0.05 level. Significant means were compared by the Duncan’s multiple range tests. All statistical analyses were carried out using SPSS statistical software package (SPSS, Version 15, Chicago, USA).
RESULTS
Transgenic tobacco plant: A previous study from our laboratory revealed that the endogenous level of H2O2 were enhanced in the tobacco plants by the expression of gene coding for GO from A. niger.
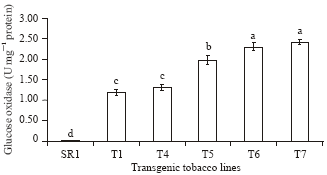 | |
| Fig. 1: | GO specific activities of transgenic tobacco plants. Bars with different letters are significantly different at p = 0.05 |
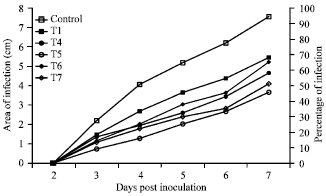 | |
| Fig. 2: | Infection rate of Rhizoctonia solani on four week old transgenic tobacco lines |
The putative transformants, as identified by GUS (β-glucuronidase) and PCR analyses, were grown on an organic mixture under controlled (16-h light/24°C) conditions (Maruthasalam et al., 2010).
GO activity in transgenic tobacco plants: The transgenic lines possessed ~1-3 units’ specific GO activity mg-1 total protein. Moreover, the specific GO activity varied significantly between the transgenic tobacco lines (Fig. 1).
Disease resistant level of transgenic tobacco plants
Fungal leaf spot disease: In the present study, transgenic tobacco expressing GO gene were tested against Rhizoctonia solani. Figure 2 reveals the results of the tobacco plant leaf infection test with the symptoms caused by R. solani. The infection rate progressed more slowly in GO transformed tobacco lines than on the wild SR1 plant (Control). The infection has started on the leaf surface of control and transgenic lines at third day after inoculation with R. solani. However infection started on transgenic line T7 at fourth day after inoculation of R. solani. The infection has completely covered the leaf surface 100% at 7 days post inoculation on control plants, but the transgenic lines were resistant to infection and 50-70% of leaf surface only infected at 7 days post inoculation (Fig. 2).
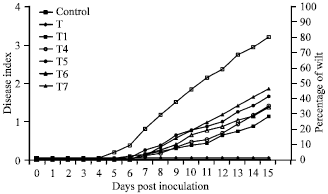 | |
| Fig. 3: | Virulence of Ralstonia solanacearum in GO transformed tobacco |
At end of the assay, percentage of infection rate on transgenic lines T1, T4-T7 were decreased by 67.78, 68.84, 76.42, 57.78 and 45.36%, respectively, compared with nontransgenic control line. As a result, reduced rates of necrotic lesions were obtained from the infected transgenic lines. All the transgenic lines were statically significant from the control lines.
Bacterial wilt disease: The wilt disease resistant ability of the GO transformed tobacco plants were tested against the bacterial pathogen Ralstonia solanacearum. Six week old transgenic tobacco plants were inoculated with wilt pathogen Ralstonia solanacearum using a natural soil-soak inoculation method. Under these conditions, wild-type tobacco (SR1) plants completely wilted almost all plants (80-100%) at 15 days post inoculation, with an average disease index of 3.8 (Fig. 3). However, GO transformed transgenic tobacco plants (T1, T4-T7) were significantly resistant in virulence to Ralstonia solanacearum, with an average disease index of 1 - 2 by the end of the assay (15 days post inoculation). All the transgenic lines were statically significant from the control lines.
Figure 4 showed the transgenic tobacco plants resistant against wilt disease caused by Ralstonia solanacearum on 13 days post inoculation. All transgenic tobacco plants (T1, T4-T7) were observed resistant to wilt disease compared to SR1b (control) tobacco plants. SR1b tobacco plant was more susceptible to wilt pathogen and appeared stunted growth. Among the transgenic tobacco plants, T1 tobacco plant showed more resistant to wilt pathogen followed by T5, T6, T4 and T7 tobacco plants. However, normal growth was observed only in uninoculated transgenic (T) and control (SR1a) tobacco plants.
Increased accumulation of H2O2 in transgenic tobacco plants: The levels of H2O2 in different leaf tissues of transgenic tobacco plants were determined.
 | |
| Fig. 4: | Resistance of GO-transformed tobacco plants to wilt disease caused by R. solanacearum on 13 days post inoculation evaluated by soil soaking method |
 | |
| Fig. 5(a-b): | Detection of elevated production of H2O2 in plant tissues of GO- transformed five week old tobacco plants on (a) Kl and (b) Starch medium |
The elevation of H2O2 levels in tissues of transgenic plants could be detected visually by placing young plants or plant tissues into a medium containing Kl and starch. H2O2 diffusing from tissues of transgenic plants was indicated by the purple-blue color in surrounding areas in the medium. Using such an assay, elevated production of H2O2 was observed in transgenic tobacco plants tissues, whereas no H2O2 could be detected in the non-transgenic control plants (Fig. 5). From the Table 1, the amount of H2O2 in leaf tissues of transgenic lines were determined to be 5-10 fold higher than control lines. Interestingly, the amount of H2O2 in transgenic tobacco plants were increased after the leaves inoculated with R. solani, whereas little difference could be detected in control lines.
| Table 1: | H2O2 elevation in five week old transgenic tobacco expressing the GO gene before and after infection with R. solani |
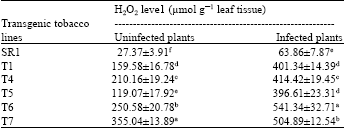 | |
| Values in the column are means of three replicate and column with same letter are statistically not significant at p = 0.05 using Duncan’s multiple range test | |
The maximum amount of H2O2 content was recorded in T7 transgenic tobacco plant (355.04±13.89 μmol g-1) and least amount was recorded in SR1 (control) tobacco plant (27.37±3.91 μmol g-1). Incase of R. solani infected tobacco plants, the maximum amount of H2O2 content was recorded in T7 transgenic tobacco plant (541.34±32.71 μmol g-1) and least amount was recorded in SR1 tobacco plant (63.86±7.87 μmol g-1). However, more difference of H2O2 content was observed before (119.07±17.92 μmol g-1) and after (396.61±23.31 μmol g-1) infection of T5 transgenic plant followed by other transgenic plants.
DISCUSSION
The transgenic tobacco plants that constitutively produce elevated sub lethal levels of H2O2 provide a unique tool to study the disease resistant level of transgenic tobacco plant against the fungal pathogen R. solani and bacterial wilt pathogen Ralstonia solanacearum. The endogenous level of H2O2 were enhanced in the tobacco plants by the expression of GO from A. niger. The earlier studies showed expression of GO from Talaromyces flavus resulted in significantly deleterious phenotypes when either constitutively expressed (Murray et al., 1999) or induced by specific signals (Kazan et al., 1998; Murray et al., 1999). Similarly Lee et al. (2002) reported constitutively expression of GO from A. niger clearly indicate the deleterious effects on cabbage. However, Lee et al. (2002) found a transgenic tobacco line with high GO activity, showing a normal germination rate, indicating that the plant could tolerate the phytotoxic effects of GO. In this study clearly indicates a normal growth pattern of tobacco plants carrying the GO gene. Similarly, Wu et al. (1995) described the normal growth of transgenic potato with constitutive expression of GO from A. niger. Frederick et al. (1990) described GO has not been found in plants and absence of the enzyme activity in non transgenic plants. In present study also the GO activity was only detected in transgenic tobacco lines and absent in control tobacco plants. Constitutive expression of GO may confer an advantage on plants so that they can tolerate the elevated level of H2O2, presumably by inducing an endogenous antioxidant system (Lee et al., 2002).
GO has previously been found to be the major antimicrobial metabolite of the fungus Talaromyces flavus and A. niger used in the biological control of soil-borne pathogens (Kim et al., 1988) presumably through generation of H2O2. Fukuta et al. (2012) expressed the antimicrobial lactoferricin gene in tobacco plants and transgenic plants showed enhanced resistance to bacterial and fungal diseases. One of the most common diseases of tobacco seedlings is damping-off caused by Rhizoctonia solani (Lucas, 1975; Shew, 1991) and some isolates of R. solani cause target spot to the tobacco leaf (Shew and Main, 1990). In the present study, transgenic tobacco plants expressing GO gene were resistant against R. solani and the infection rate progressed more slowly than wild control plants. Other than the fungal disease, bacterial wilt caused by Ralstonia solanacearum, is a serious disease of tobacco plants (Robertson et al., 2004). The present results showed GO transformed transgenic tobacco plants were significantly resistant in virulence to Ralstonia solanacearum, compared to control plants.
The results demonstrate that H2O2 production can be manipulated in the plant through expression of a transgene and be linked to enhanced resistance of the plants to pathologically diverse pathogens. In tobacco, the effective control of both the bacterial wilt disease and the funga1 target spot disease occurred by virtue of elevated levels of H2O2. The previous studies showed GO transformed transgenic plants enhanced disease resistance to a wide range of pathogens such as Erwinia carotovora subsp. carotovora, Phytophthora infestans and Verticillium dahlia in transgenic potato (Wu et al., 1995), Rhizoctonia solani in transgenic tobacco (Murray et al., 1999) and Verticillium dahliae in transgenic cotton (Murray et al., 1999).
Brunner et al. (2005) reported the transgenic Trichoderma strain expressing glucose oxidase for plant pathogen control has the advantage that H2O2 accumulation occurs only in the presence of the pathogen and reduces the accumulation of H2O2 in the absence of a pathogen attack. Interestingly, the amount of H2O2 content were increased after the transgenic tobacco leaf inoculated with R. solani, whereas little difference could be detected in control lines. This result strongly suggests that elevated H2O2 levels mediate strong resistance to target spot in transgenic tobacco plants. That H2O2 elevation accounts for the increased disease resistance shown by the GO transgenic plants is confirmed by our observation. As H2O2 is a potent signal which induces plant defense responses, it has been a point of interest whether the enhanced disease resistant in GO-transformed plants is associated with the induction of plant defense genes by elevated levels of H2O2 (Lee et al., 2002). Evidently, H2O2 generated by GO is an absolute necessity for expose the transgenic plants disease resistant.
H2O2 elevation is extremely rapid and intense and is postulated to have multiple functions in the active defense of plant cells (Mehdy, 1994; Baker and Orlandi, 1995) as well as in immune responses of animal cells (Schreck et al., 1991). The enhanced resistance to both a bacterial and a fungal pathogen exhibited by GO transformed plants demonstrates the potential of obtaining broad spectrum resistance by manipulating the endogenous levels of H2O2 in tobacco plants. It is likely that increased levels of H2O2 in transgenic plants confer resistance to these diverse pathogens via a common mechanism.
H2O2 also serve as a signal in turning on the natural defense mechanisms of plants during interactions with the pathogens (Wu et al., 1995). Earlier studies used exogenously supplied O2~ or H2O2 to leaf tissue or cells in suspension cultures have shown that these AOS trigger hypersensitive cell death (Adam et al., 1989; Levine et al., 1994) which often leads to systemic acquired resistance in plants (Wu et al., 1995). In addition, H2O2 has been found to influence gene expression in a number of defense-related cellular processes (Wu et al., 1995). H2O2 was found to stimulate phytoalexin production by potato tuber slices (Chai and Doke, 1987) and to induce glyceollin biosynthesis by soybean cells in suspension cultures (Apostol et al., 1989; Degousee et al., 1994).
The transgenic tobacco plants in which the level of H2O2 is elevated endogenously now provide us with a unique system to study more directly the involvement of H2O2 in plant defense processes. The use of an AOS-generating enzyme in plants also represents a novel and effective strategy of engineering disease resistance in agricultural crops. Given the nonspecific oxidative toxicity of H2O2 to a large number of plant microbial pathogens and its critical role in plant disease resistance, the approach described here may yield broad spectrum resistance to plant pathogens.
CONCLUSION
The present study showed that expressions of a gene encoding H2O2 generating GO confers resistance to bacterial and fungal pathogens in transgenic tobacco plants. The results demonstrate that H2O2 production can be manipulated in the plant through expression of a transgene and be linked to enhanced resistance of the plants to pathologically diverse pathogens. Evidently, H2O2 generated by GO is an absolute requirement for rendering the transgenic plants disease resistant.
ACKNOWLEDGMENT
This study was supported by the National Science Council (NSC), Taiwan, Republic of China.
REFERENCES
- Adam, A., T. Farkas, G. Somlyai, M. Hevesi and Z. Kiraly, 1989. Consequence of O2·- generation during a bacterially induced hypersensitive reaction in tobacco: Deterioration of membrane lipids. Physiol. Mol. Plant Pathol., 34: 13-26.
CrossRef - Amil-Ruiz, F., R. Blanco-Portales, J. Munoz-Blanco and J.L. Caballero, 2011. The strawberry plant defense mechanism: A molecular review. Plant Cell Physiol., 52: 1873-1903.
CrossRefDirect Link - Baker, C.J. and E.W. Orlandi, 1995. Active oxygen in plant pathogenesis. Annu. Rev. Phytopathol., 33: 299-321.
CrossRefDirect Link - Bent, A.F. and I.C. Yu, 1999. Applications of molecular biology to plant disease and insect resistance. Adv. Agron., 66: 251-298.
CrossRef - Brunner, K., S. Zeilinger, R. Ciliento, S.L. Woo, M. Lorito, C.P. Kubicek and R.L. Mach, 2005. Improvement of the fungal biocontrol agent Trichoderma atroviride to enhance both antagonism and induction of plant systemic disease resistance. Applied Environ. Microbiol., 71: 3959-3965.
CrossRefDirect Link - Degousee, N., C. Triantaphylides and J.L. Montillet, 1994. Involvement of oxidative processes in the signaling mechanisms leading to the activation of glyceollin synthesis in soybean (Glycine max). Plant Physiol., 104: 945-952.
Direct Link - Felcher, K.J., D.S. Douches, W.W. Kirk, R. Hammerschmidt and W. Li, 2003. Expression of a fungal glucose oxidase gene in three potato cultivars with different susceptibility to late blight (Phytophthora infestans Mont. deBary). J. Am. Soc. Hortic. Sci., 128: 238-245.
Direct Link - Frederick, K.R., J. Tung, R.S. Emerick, F.R. Masiarz and S.H. Chamberlain et al., 1990. Glucose oxidase from Aspergillus niger. Cloning, gene sequence, secretion from Saccharomyces cerevisiae and kinetic analysis of a yeast-derived enzyme. Biol. Chem., 265: 3793-3802.
Direct Link - Fukuta, S., K. Kawamoto, Y. Mizukami, Y. Yoshimura, J.I. Ueda and M. Kanbe, 2012. Transgenic tobacco plants expressing antimicrobial peptide bovine lactoferricin show enhanced resistance to phytopathogens. Plant Biotechnol., 29: 383-389.
Direct Link - Ganapathi, T.R., P. Suprasanna, P.S. Rao and V.A. Bapat, 2004. Tobacco (Nicotiana tabacum L.): A model system for tissue culture interventions and genetic engineering. Indian J. Biotechnol., 3: 171-184.
Direct Link - Gechev, T.S., F. van Breusegem, J.M. Stone, I. Denev and C. Laloi, 2006. Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. BioEssays, 28: 1091-1101.
CrossRefPubMedDirect Link - Hilbeck, A., S. Jansch, M. Meier and J. Rombke, 2008. Analysis and validation of present ecotoxicological test methods and strategies for the risk assessment of genetically modified plants. Bundesamt fur Naturschutz (BfN)-Skripten 236, Federal Agency for Nature Conservation, Bonn, Germany, pp: 1-287.
- Honee, G., 1999. Engineered resistance against fungal plant pathogens. Eur. J. Plant Pathol., 105: 319-326.
CrossRef - Houot, V., P. Etienne, A.S. Petitot, S. Barbier, J.P. Blein and L. Suty, 2001. Hydrogen peroxide induces programmed cell death features in cultured tobacco BY-2 cells, in a dose-dependent manner. J. Exp. Bot., 52: 1721-1730.
CrossRef - Hwang, S.C. and W.H. Ko, 2004. Cavendish banana cultivars resistant to fusarium wilt acquired through somaclonal variation in Taiwan. Plant Dis., 88: 580-588.
CrossRefDirect Link - Kang, G., C. Wang, G. Sun and Z. Wang, 2003. Salicylic acid changes activities of H2O2-metabolizing enzymes and increase the chilling tolerance of banana seedlings. Environ. Exp. Bot., 50: 9-15.
Direct Link - Kazan, K., F.R. Murray, K.C. Goulter, D.J. Llewellyn and J.M. Manners, 1998. Induction of cell death in transgenic plants expressing a fungal glucose oxidase. Mol. Plant-Microbe Interact., 11: 555-562.
CrossRefDirect Link - Keen, N.T., 1999. Plant disease resistance: Progress in basic understanding and practical application. Adv. Bot. Res., 30: 292-328.
CrossRef - Kim, K.K., D.R. Flavel and G.C. Papavizas, 1988. Identification of a metabolite produced by Talaromyces flavus as glucose oxidase and its role in the biocontrol of Verticillium dahliae. Phytopathology, 78: 488-492.
CrossRefDirect Link - Lee, Y.H., I.S. Yoon, S.C. Suh and H.I. Kim, 2002. Enhanced disease resistance in transgenic cabbage and tobacco expressing a glucose oxidase gene from Aspergillus niger. Plant Cell Rep., 20: 857-863.
CrossRef - Levine, A., R. Tenhaken, R. Dixon and C. Lamb, 1994. H2O2 from the oxidative burst orchestrates the plant hypersensitive disease resistance response. Cell, 79: 583-593.
CrossRefPubMedDirect Link - Maruthasalam, S., Y.L. Liu, C.M. Sun, P.Y. Chen, C.W. Yu, P.F. Lee and C.H. Lin, 2010. Constitutive expression of a fungal glucose oxidase gene in transgenic tobacco confers chilling tolerance through the activation of antioxidative defence system. Plant Cell Rep., 29: 1035-1048.
CrossRef - Mehdy, M.C., 1994. Active oxygen species in plant defense against pathogens. Plant Physiol., 105: 467-472.
Direct Link - Miller, G., N. Suzuki, S. Ciftci-Yilmaz and R. Mittler, 2010. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ., 33: 453-467.
CrossRef - Murray, F., D. Llewellyn, H. McFadden, D. Last, E.S. Dennis and W.J. Peacock, 1999. Expression of the Talaromyces flavus glucose oxidase gene in cotton and tobacco reduces fungal infection, but is also phytotoxic. Mol. Breed., 5: 219-232.
CrossRef - Olson, P.D. and J.E. Varner, 1993. Hydrogen peroxide and lignication. Plant J., 4: 887-892.
Direct Link - Petrov, V.D. and F. Van Breusegem, 2012. Hydrogen peroxide: A central hub for information flow in plant cells. AoB Plants.
CrossRef - Robertson, A.E., W.P. Wechter, T.P. Denny, B.A. Fortnum and D.A. Kluepfel, 2004. Relationship between avirulence gene (avrA) diversity in Ralstonia solanacearum and bacterial wilt incidence. Mol. Plant-Microbe Interact., 17: 1376-1384.
CrossRefDirect Link - Rommens, C.M. and G.M. Kishore, 2000. Exploiting the full potential of disease-resistance genes for agricultural use. Curr. Opin. Biotechnol., 11: 120-125.
CrossRef - Schreck, R., P. Rieber and P.A. Baeuerle, 1991. Reactive oxygen intermediates as apparently widely used messengers in the activation of the NF-κB transcription factor and HIV-1. EMBO J., 10: 2247-2258.
PubMedDirect Link - Shew, H.D. and C.E. Main, 1990. Infection and development of target spot of flue-cured tobacco caused by Thanatephorus cucumeris. Plant Dis., 74: 1009-1013.
Direct Link - Thordal-Christensen, H., Z. Zhang, Y. Wei and D.B. Collinge, 1997. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley-powdery mildew interaction. Plant J., 11: 1187-1194.
CrossRefDirect Link - Villajuan, A.R., K. Kageyama and M. Hyakumachi, 1996. Biocontrol of Rhizoctonia damping-off of cucumber by non-pathogenic binucleate Rhizoctonia. Eur. J. Plant Pathol., 102: 227-235.
CrossRefDirect Link - Wu, G., B. Shortt, E. Lawrence, E. Levine, K. Fitzsimmons and D. Shah, 1995. Disease resistance conferred by expression of a gene encoding H2O2 generating glucose oxidase in transgenic potato plants. Plant Cell, 7: 1357-1368.
Direct Link - Zupan, J.R. and P. Zambryski, 1995. Transfer of T-DNA from Agrobacterium to the plant cell. Plant Physiol., 107: 1041-1047.
Direct Link