
K.G. Hossain
Mayville State University, Mayville, North Dakota, United States
N. Islam
University of Maryland College Park, MD, United States
D. Jacob
North Dakota State University, Fargo, North Dakota, United States
F. Ghavami
North Dakota State University, Fargo, North Dakota, United States
M. Tucker
Mayville State University, Mayville, North Dakota, United States
T. Kowalski
Mayville State University, Mayville, North Dakota, United States
A. Leilani
Mayville State University, Mayville, North Dakota, United States
J. Zacharias
Mayville State University, Mayville, North Dakota, United States
Asian Journal of Plant Sciences
Year: 2013 | Volume: 12 | Issue: 1 | Page No.: 11-20







ABSTRACT
Essential minerals are considered as key determinants of optimum health and nutritive quality of common bean seed. This study aimed to identify genetically stable essential minerals in common bean. Eleven diverse common bean genotypes were grown in three distinct growing environments and 17 essential minerals were analyzed by Inductively Coupled Plasma-Optical Emission Spectroscopy. Genetic control of mineral composition in common bean seed was demonstrated by large (p<0.01) genotypic differences in Ca and Sr contents and moderate genotypic difference was observed in Fe content. Significant influence of genotype and environments (GxE) interaction was observed in the content of all minerals. The ratios between genetic and environmental variances and between genetic and GxE variances indicated the greater influence and stability of genetic factor on the concentration of Ca and Sr in common bean seed. Significant positive correlations among important minerals such as Zn with S, P, Fe and Na and Cu with K, Mg, Ni, P were identified. The stability of genetic effects on Ca and Sr concentration in common bean has been identified in this study. Calcium is one of the most important minerals which regulates many cellular processes and has important structural roles in living organisms. Further studies to characterize Ca physiology in common bean may identify genetic or biochemical markers to expedite breeding common bean with enhanced Ca concentration.
PDF Abstract XML References Citation
Received: January 29, 2013;
Accepted: April 05, 2013;
Published: June 21, 2013
How to cite this article
K.G. Hossain, N. Islam, D. Jacob, F. Ghavami, M. Tucker, T. Kowalski, A. Leilani and J. Zacharias, 2013. Interdependence of Genotype and Growing Site on Seed Mineral Compositions in Common Bean. Asian Journal of Plant Sciences, 12: 11-20.
DOI: 10.3923/ajps.2013.11.20
URL: https://scialert.net/abstract/?doi=ajps.2013.11.20
DOI: 10.3923/ajps.2013.11.20
URL: https://scialert.net/abstract/?doi=ajps.2013.11.20
INTRODUCTION
Diets containing a diverse and well balanced mixture of both macronutrients and micronutrients determine the optimal health of human being. Macronutrients make up the bulk of foodstuff and meet the energy demand of human body while micronutrients, commonly referred as vitamins and minerals (Omenn, 1996), are essential for health management. Science-driven progress in agriculture in the last 100 years has resulted in achieving lifts in crop productivity needed to provide adequate food energy for the world. However, according to the statistics of the United Nation's World Health Organization and World Bank, the adequate food supply is not providing adequate balanced nutrition, resulting in worldwide malnutrition (Anonymous, 1992). The FAO and WHO nutritionists have identified micronutrient malnutrition as a primary health care issue and today over three billion people are afflicted with micronutrient malnutrition and the numbers are increasing (Mason and Garcia, 1993; Welch et al., 1997; WHO, 2002). Nearly two-thirds of all deaths of children are associated with nutritional deficiencies that lead to the high rate of morbidity and mortality and adverse effects on learning ability, development and growth in infants and children (Caballero, 2002). Micronutrient deficiencies have been identified as prime cause of childhood stunting of children from early fetal stages of development through the fourth year of life (Branca and Ferrari, 2002). This situation is particularly at alarming stage in many developing nations (Mason and Garcia, 1993; WHO, 2002). Minerals like aluminum (Al), boron (B), beryllium (Be), calcium (Ca), copper (Cu), iron (Fe), potassium (K), magnesium (Mg), manganese (Mn), molybdenum (Mo), sodium (Na), nickel (Ni), phosphorus (P), sulfur (S), strontium (Sr), titanium (Ti) and zinc (Zn), are essential and necessary for normal growth, reproduction and health (Phan-Thien et al., 2010). They are not strictly belonging to functional foods however; several of these minerals have preventative and therapeutic effects on chronic diseases. For example, the most worldwide prevalent malnutrition is linked to Fe deficiency and is a leading risk factor for disability and death. Iron (Fe) deficiency anemia is much less prevalent in developed countries compared to developing countries but still accounts for more than two-thirds of anemia cases in general practice (Zimmermann and Hurrell, 2007). It has been found that Ca consumption is linked to the prevention of osteoporosis and identified as a major area of opportunity for functional food innovations (Tapsell et al., 2005). Diet with high K can lower blood pressure and reduce cardiovascular disease mortality. It can also delay the progression of renal disease and aid in the management of kidney stone disease (He and MacGregor, 2008). Interest in plant foods mineral composition has been restricted to fortification rather than biofortification, or the genetic manipulation of mineral content through plant breeding. There are limited crop breeding program focused on enhancement or improvement mineral composition, with the notable exception of a large CGIAR (Consultative Group on International Agricultural Research) program aiming to increase bioavailable Fe, Zn and carotenoids in a number of staple food crops (Welch and Graham, 2004). Along with the more traditional interventions like supplementation, fortification and dietary diversification, biofortification of staple food products is seen as one of the key strategies for alleviating micronutrient malnutrition afflicting poor communities (White and Broadley, 2005). Varieties enhanced with essential minerals are readily adopted by small-scale farmers because no changes in agronomic practice or extraordinary capital investments are required. Micronutrient-enriched seeds would also have the added advantage of boosting productivity in nutrient-deficient soils which are typically cultivated by the rural poor of the developing world (Welch and Graham, 2004). Strategic utilization of diverse genetic resources for developing high micronutrient content plant food either by conventional breeding or molecular techniques is essential to ensure better living of human being (Bouis, 2000; Combs, 1996).
The common bean (Phaseolus vulgaris L.) is the principal grain legume which is a valuable and cheap source for minerals or micronutrients. However, minerals contents in this crop, like any other plant species, dependent on the availability of minerals in soil environment, acquisition by roots and efficient transportation and storage mechanism. The physiological basis and controlling of absorption and accumulation mineral/micronutrients and metals in edible portion of seeds and grains are not well understood (Welch, 1986). However, mineral contents in storage tissues are influenced by the soil environments, genotypes and the genotype by environment (GxE) interaction (Phan-Thien et al., 2010; Zhe et al., 2010; Rao et al., 2002; Paredes et al., 2009). Numerous genes may be involved in controlling the amount of a mineral element that is absorbed by roots, translocate to shoots, remobilized from vegetative tissues and deposited in edible portions of seeds and grains in forms that are utilizable by the person eating the crop (Welch and House, 1984). However the absorption and utilization (bioavailability) of any micronutrient enriched (bio fortified) food is determined by the interactions of the micronutrient with various dietary factors. (Hotz and Gibson, 2007; Ortiz-Monasterio et al., 2007).
There are several studies on mineral concentrations in common bean but most of the studies either limited to genotypes from specific geographical area for assessing mineral contents and diversity among genotypes for mineral compositions (Paredes et al., 2009; Pinheiro et al., 2010) or few minerals (Pfeiffer and McClafferty, 2007; Cichy et al., 2009; Nchimbi-Msolla and Tryphone, 2010). In addition, genetic diversity of common bean in relation to mineral content (Talukder et al., 2010), minerals and anti-nutrient constituents (Akond et al., 2011a) and antioxidant activities (Akond et al., 2011b) is also reported. However, a comprehensive study based on most of the essential minerals in diverse genotype under different environment is lacking. The present study was conducted to analyze the contents of 17 essential minerals in 11 common bean genotypes selected for their diversity in mineral content. The genotypes were grown in three different environments to (1) Estimate the relative influence of genotype, environment and their interaction on the concentrations of various minerals in common bean seed, (2) Evaluate the relative influence of different components of variation on mineral concentrations in common bean and (3) Identify the relative association among the minerals in common bean seed.
MATERIALS AND METHODS
Plant materials: Samples comprised 11 common bean genotypes with different origin and background were selected from a previous study (Talukder et al., 2010). Seeds of these genotypes were grown in 18x19 cm pots filled with Sunshine mix 1 (Sun Gro Horticulture Canada Ltd., formulated with Canadian sphagnum peat moss, coarse grade perlite, gypsum and dolomitic lime) as substrate at Mayville State University in summer 2010. Two seeds for each entry were placed in each pot for germination; one plant was allowed to grow until harvest of the seeds. Pots were watered periodically with tap water to the approximate field capacity to facilitate normal plant growth. No additional fertilizer or pesticide as applied during the period of experimentation. Seeds of each genotype were planted in two different fields of North Dakota State University, Fargo, ND in June, 2010 and 2011 and harvested in October 2010 and 2011, respectively. All of these trails were conducted in three replications.
Inductively coupled plasma-optical emission spectroscopy (ICP-OES) for mineral concentration: Digestion and ICP-OES analysis were performed at the Metal Analysis Core Laboratory of ND INBRE at NDSU for 17 minerals: aluminum (Al), boron (B), beryllium (Be), calcium (Ca), copper (Cu), iron (Fe), potassium (K), magnesium (Mg), manganese (Mn), molybdenum (Mo), sodium (Na), nickel (Ni), phosphorus (P), sulfur (S), strontium (Sr), titanium (Ti) and zinc (Zn), contents in common bean seeds. After harvesting, seeds from each pod of individual plant were mixed thoroughly and 10 seeds were randomly taken for chemical analysis. The seeds were ground in liquid nitrogen with mortar and pestle until a relatively homogeneous particle size was achieved. Closed acid digestion was performed in a Mars Xpress Microwave system (CEM) and 55 mL PFA (Paraformaldehyde) venting vessels. The digestion mixture consisted of 250 mg of sample, 5 mL of concentrated nitric acid and 5 mL of water. The mixture was digested in microwave for 25 min at 185°C. Analyses of element concentration were performed on Spectro.
Genesis ICP-OES using Smart Analyzer Vision software (v. 3.013.0752). Quality control consisted of continuous calibration verification, internal standardization and simultaneous analysis of certified reference plant material.
Data analysis: Seed mineral contents data were subject to analysis of variance (ANOVA) for identifying genotype, environment and genotype x environment effect on 17 mineral contents in common bean. Analysis of variance of the phenotypic data was performed using GLM procedure of the Statistical Analysis System (version 8.0, SAS Institute Inc., Cary, NC, USA). Pearson correlation coefficients were used to evaluate correlations for pair-wise bran components traits by using the CORR procedure of SAS.
RESULTS AND DISCUSSION
Analysis of variances and ranges in mineral concentrations: Highly significant (p<0.01) influence of genotype (G) was observed on Ca and Sr content in common bean seed and significant (p<0.05) influence of genotype was observed on Fe content. In common bean seed, highly significant influence of growing sites were observed on Ca, Cu, Fe, Mg, Mo, Na, Ni and Sr and significant influence of growing environments were observed on Be, S and Zn contents. While no significant influence of genotype and growing sites s was observed on 14 and six seed mineral contents respectively, seed content of all minerals in this study were highly significantly influenced by the interaction of genotype and growing sites (GxE) (Table 1) Among the minerals, Ca and Sr contents of common bean seed were highly significantly influenced by the genotype, growing sites and genotype by growing site interactions. The highly significant influences of growing sites and genotype by growing site interactions were observed on seed Fe content when significant influence of genotype was observed for this mineral content in the common bean seed.
Variations in mineral content (Table 2) indicated that except the genotypic variation of Cu, Mg, Mo and Ni content, concentrations of all other minerals varied widely among genotypes compared to the variation associated with growing environments. Among the genotypes, the Cu content ranged from 4.04 to 9.68 μg g-1, Mg content ranged from 1221.68 to 1550.70 μg g-1, Mo content ranged 6.95 to 20.55 μg g-1 and Ni content ranged from 1.49 to 2.73 μg g-1 which are comparable to the ranges of these minerals among growing sites (Table 2). In some instances, the wider genotypic variabilities are due to a single genotypes such as Al content ranged from 7.27 to 43.61 μg g-1 and a single genotype A55 contributed 43.61 μg g-1 and among the rest, the Al contents ranged from 7.27 to 18.42 μg g-1 (Table 3). The relatively wider overall variability of mineral contents compared to the variabilities of mineral contents associated with growing sites and genotypes was due to the highly significant influence of GxE on mineral compositions under this study. Nchimbi-Msolla and Tryphone (2010) reported significant genotypic and environmental influence on seed Fe and Zn concentrations when they conducted their trial in two different locations with 10 different genotypes of common bean. In Burundi, K content ranged from 4420 to 6310 μg g-1 (Barampama and Simard, 1993). Several studies has shown that both genetics and the environment influence the K content in bean seeds (Barampama and Simard, 1993; Moraghan and Grafton, 2001).
Ratios of among variance components: Variation is broadly considered to have three components, i.e., genetic/genotypic, environmental and genotype-by-environment (GxE) interaction. GxE interaction introduces uncertainty into the selection process and therefore, influences the estimation of trait heritability and response to selection. A ratio of the variances associated with the environmental effects (σ2E) to the variances associated with the genetic effects (σ2G) provides a means for examining the relative influences of genotype and environment on mineral composition in common bean. A ration larger than 1.0 indicates the greater influence of environmental factors on variability and a ratio less than 1.0 indicates the relatively greater influence of genetic factors. Ratios of variances between environmental effect and genetic effects (Table 4) indicated highest influence of environment on the variability of Mo concentration (23.30) in common bean seed followed by Cu (8.6), S (3.54), Ni (3.00), Zn (1.51), Mn (1.29) and B (1.28).
| Table 1: | Mean squares from Analysis of Variance (ANOVA) of seed mineral concentrations of 11 common bean genotypes grown in three different growing sites |
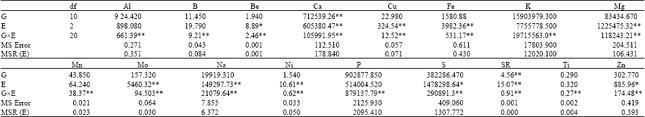 | |
| G: Genotype; E: Growing site; G×E: Genotype by growing site interaction; Df: Dgrees of freedom. *Significant at 5% level of probability; **significant at 1% level of probability | |
| Table 2: | Ranges of 17 essential mineral concentrations among growing sites (location) and genotypes and the overall ranges of mineral concentration in common bean seed |
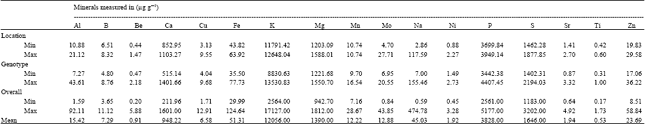 | |
| Table 3: | Mean of the 17 essential seed mineral concentrations of 11 genotypes of common bean grown in three different growing sites |
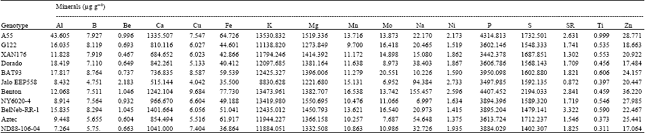 | |
| Table 4: | Variances of seed mineral concentrations common bean associated with growing site (σ2E), genotype (σ2G ) and genotype x growing site (σ2G×E) and ratios of variances between σ2E and σ2G and between σ2G×E and σ2G |
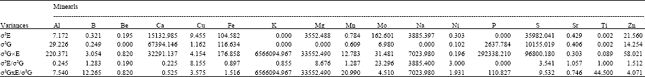 | |
The greatest influence of genetic factor on the variability of Ca concentration (0.23) was observed in common bean seed followed by Al (0.25), Fe (0.90) and almost equal influence of environment and genetic factors was observed on the variability of Ti (1.00) and Sr (1.10).
The magnitude of the GxE interaction in relation to genetic effects can be calculated by the variance component ration of σ2GXE/σ2G (Table 4). A ratio smaller than 1.0 indicates greater influence and stability of genetic factors relative to the variability associated with the interaction of genotype and environment. The ratio between variances associated with environment factor and genetic factors indicated a higher influence of genotype on the concentrations of Ca (0.225), Al (0.245), Fe (0.897) but the ratio between σ2GXE/σ2G indicated the greater influence and stability of genetic factor on the concentration of Ca (0.525) in common bean seed. Although the equal contribution of environment and genotype on the concentration of Sr was indicated but a ration of σ2GXE/σ2G smaller than 1.0 (0.75) indicated the greater influence and stability of genetic factor in controlling the Sr concentration in common bean seed. In a study in wheat Peterson et al. (1983) observed larger influence of genotype on Mg and P variability. They also reported higher influence of environment on Ca, Fe, Cu and Zn. The ratios of Mg showed a larger influence on the variability by genotype than GxE interaction. Ratios for Ca, Mn and Zn were less that 1.0 indicated the important influence of the GxE interaction on the concentrations of these elements. Iron ratios of larger than 1.0 indicated the larger influence of genotype. In their study, they reported a larger number of environments must be evaluated to select genotypes with higher concentration of most of the minerals with the exception of Mg and Fe.
The 11 genotypes in this study were selected for the diversity of their mineral content according to a study consisting of 29 genotypes (Talukder et al., 2010). The stability of genetic differences of several elements such as Ca, Sr and Fe was estimated by significant genotypic effect according to ANOVA (Table 1) but ratios between genotypic and environmental variances and genotypic and GxE variances (Table 4) confirmed that the stability of genetics differences for Ca and Sr in common bean seed. Common bean is very important legume and is a chief source of minerals for human consumption. However, uptake and translocation of minerals to storage tissues are dependent on the availability of minerals in the soil. In this study, the larger influence of GxE on the concentration of 15 different minerals suggests that the selection genotypes for improving the concentration of these mineral concentrations in common bean may be an unrealistic approach. However, larger-scale data analysis covering a greater number of genotypes, environments and seasons is required to fully characterize the GxE interaction affecting these mineral accumulation in common bean seed.
The stability of genetic effects for Ca and Sr concentration in common bean has been confirmed in this study. In particular the genotypes A55 and BelNeb-RR-1 contained consistently high concentration of Ca and BelNeb-RR-1 contained a high concentration of Sr (Table 3). The genotypic effect on Ca concentration has also been reported in other species such in peanut (Phan-Thien et al., 2010) and chick pea (Abbo et al., 2000). Calcium is one of the most important minerals which regulates many cellular processes and has important structural roles in living organisms (Tandogan and Usulu, 2005). Calcium is an important component of bone and its consumption is linked to the prevention of osteoporosis and identified as a major area of opportunity for functional food innovations (Tapsell et al., 2005). Calcium deficiency is a common concern worldwide and according to National Health and Nutrition Examination Survey (NHANES) that the mean dietary calcium intake among adults in the U.S. is well below the recommended intake for adults of 1, 000 to 1, 200 mg day-1 (Pasco et al., 2000). There is limited research on importance of Sr in human health. It has recently been considered Sr as dietary supplement for bone health and it is reported that it may improve bone density in osteoporosis or osteopenia (Ortolani and Vai, 2006). However, higher concentration of Ca when compared to Sr (Table 3) makes common bean seed a better choice for Ca from nutritional point of view. It is clear that there is a strong genetic component to Ca accumulation and controlled studies to better characterize Ca physiology in common bean may generate specific genetic or biochemical markers that could be used in breeding programs for selecting lines with enhanced Ca content. Traits that lead to high Ca uptake and accumulation in seeds could also reduce the need for soil amelioration as well as improve common bean nutritional quality.
Correlation among mineral contents in common bean seed: Significant positive correlations of Al concentration with Ti; Be with Mn; Ca with Sr; Cu with K, Mg, Ni, P and Sr; Fe with Na, S and Zn; K with Mg and P; Mg with Ni and P; Na with S and Zn; P with Zn; and S with Zn were observed in common bean seeds (Table 5). Significant negative correlation of Be with K and Mo with Ni was observed in common bean seed. In this study, the seed Cu content was found to be correlated with highest number of minerals (5) and Fe content was correlated with three of the minerals.
| Table 5: | Correlation coefficients and their significance among 17 seed minerals of 11 common bean genotypes grown in three growing sites |
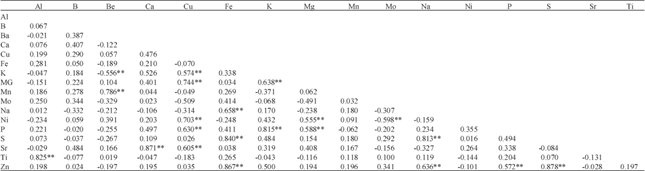 | |
| **Significant at 1% level of probability | |
Although the physiological basis of mineral deposition in the storage tissue (seed) is not well understood (Welch, 1986) but a well-designed study may be conducted by treating the common bean genotypes with increased amount of any of the correlated minerals to see if there is any positive response on other minerals. There is no report so far on the negative influence of Be on K content or Mo on Ni content or vice versa however minerals with negative correlation may have antagonistic effects on the contents of each other in common bean seed. There are several studies on the correlation among mineral composition in common bean seed. Paredes et al. (2009) analyzed mineral compositions among 246 Chilean accessions of common bean and identified that P content was positively correlated with the content of Zn, B and S; K content was positively correlated with Mg content; Cu content was positively correlated with the Zn; Fe content was positively correlated with Mn; Zn content was positively correlated with S and the B and C content were correlated positively. Hacisalihoglu et al. (2005) reported strong positive correlations of the concentrations of Fe, Cu and Mg with Zn concentration in common bean seed while the concentration P showed a moderate correlation with Zn concentration. Gelin et al. (2007) showed that the Zn content of common bean seed was positively correlated with Fe, Ca and P with the highest correlation between Fe and P. The significant positive correlation between Zn and Fe concentration in common bean seed also reported elsewhere (House et al., 2002; Pfeiffer and McClafferty, 2007). Mineral composition is also greatly influenced by the soil environment. To exclude any false positive association among mineral composition, correlations with higher R values (R = 0.55) were considered in this study. The highest correlation was observed between Zn and S (R = 0.878) followed Sr and Ca (R = 0.871) and Zn and Fe (R = 0.867). The correlation between Zn and P, K and Mg and Zn and S are inconsistent with the study of Paredes et al. (2009). The correlation between Zn with P and Fe also reported by Gelin et al. (2007) and Zn and Fe by House et al. (2002) and Pfeiffer and McClafferty (2007). Correlation of Zn and Fe content in common bean seed also reported in a study using Recombinant Inbred Lines (RILS) (Cichy et al., 2009) and identified quantitative trait loci (QTLs) of Fe and Zn co-localized in three linkage group of the common bean genome.
Knowledge on correlations is required to obtain the expected response of other traits when selection is applied to a particular trait of interest in a breeding program. It was found that the magnitude and direction of correlation coefficients of measured traits varied under different environments (Zou et al., 2011). Among the 17 minerals studied, 11 were found to be significantly correlated to one or more of the minerals in the combined effect of all three growing sites. The correlations among mineral concentrations in common bean seed are important features in understanding the behavior of traits and are of values for selecting desirable traits in a breeding program. Significant positive correlations among mineral concentrations in common bean seeds suggest that selecting bean seeds with high concentration of one mineral may increase the amount of other correlated mineral. These results suggest that seed mineral concentration values may be considered as an additional selection tools for nutrient efficiency in common bean.
CONCLUSION
The common bean is the most important grain legume consumed worldwide and it is considered as a cheap source of almost all essential mineral for human health. This study was conducted with 11 common bean genotypes grown in three different growing sites and 17 essential minerals were analyzed to understand the influence of different variation components on the contents of these minerals in common bean seed. The consistent and stable genotypic influence was observed on Ca and Sr and also strong positive correlation was observed between these mineral in common bean seed. Calcium regulates many cellular processes in human, an important component of bone and its consumption is linked to the prevention of osteoporosis. The importance of Sr in human health management is not well documents and the Ca concentration in common bean is much higher compared to Sr. The finding of this study suggests that the common bean genotypes with higher content Ca could be incorporated in the breeding to develop common bean cultivar with enhanced Ca content. The strong genetic component in Ca content will help characterize the physiology of Ca accumulation in common bean seed and generate markers that could expedite the breeding programs in selecting lines with higher Ca content. The significant positive correlations among mineral concentration in common bean seeds specially between Zn and S and Zn and Fe suggest that selecting bean seeds with high concentration of Zn may also have increase amounts of S and Fe and vice versa. These results suggest that seed mineral concentration values may be considered as an additional selection tools for improving the nutritive quality of common bean. Among the 17 minerals, 11 were found to be correlated with one or more of the minerals in common bean seed. Further studies may be suggested by treating the common bean genotype with the increased amount of correlated minerals. The positive response of the increased mineral (s) on other mineral contents will ensure the identified correlations among the minerals in this study.
ACKNOWLEDGMENTS
Research reported in this article was supported by an Institutional Development Award (IDeA) from the National Institute of General Medical Sciences of the National Institutes of Health under grant number P20GM12345.
Coauthor contributions: K.G. Hossain is the main and corresponding author and was the supervisor of this study; K. G. Hossain and N. Islam developed the concept and design the experiment; D. Jacob conducted the mineral analysis; J. Zacharias, A. Leilani, M. Tucker and T. Kowalski conducted the field and pot experiments and F. Ghavami conducted the statistical analysis.
REFERENCES
- Abbo, S., M.A. Grusak, T. Tzuk and R. Reifen, 2000. Genetic control of seed weight and calcium concentration in chickpea seed. Plant Breed., 119: 427-431.
CrossRef - Akond, A.S.M.G.M., H. Crawford, J. Berthold, Z.I. Talukder and K. Hossain, 2011. Minerals (Zn, Fe, Ca and Mg) and antinutrient (phytic acid) constituents in common bean. Am. J. Food Technol., 6: 235-243.
CrossRefDirect Link - Anonymous, 1992. World declaration and plan of action for nutrition. Proceedings of the International Conference on Nutrition, December, 1992, Agricultural Organization of the United Nations and World Health Organization, Italy, Rome, pp: 1-50.
Direct Link - Barampama, Z. and R.E. Simard, 1993. Nutrient composition, protein quality and antinutritional factors of some varieties of dry beans (Phaseolus vulgaris L.) grown in Burundi. Food Chem., 47: 159-167.
CrossRefDirect Link - Bouis, H.E., 2000. Enrichment of food staples through plant breeding: a new strategy for fighting micronutrient malnutrition. Nutrition, 16: 701-704.
PubMedDirect Link - Caballero, B., 2002. Global patterns of child health: The role of nutrition. Ann. Nutr. Metab., 46: 3-7.
CrossRefDirect Link - Cichy, K.A., G.V. Caldas, S.S. Snapp and M.W. Blair, 2009. QTL Analysis of seed iron, zinc and phosphorus levels in an andean bean population. Crop. Sci., 49: 1742-1750.
CrossRefDirect Link - Gelin, J.R., S. Forster, K.F. Grafton, P.E. McClean and G.A. Kooas-Cifuentes, 2007. Analysis of seed zinc and other minerals in a recombinant inbred population of navy bean (Phaseolus vulgaris L.). Crop Sci., 47: 1361-1366.
CrossRefDirect Link - He, F.J. and G.A. MacGregor, 2008. Beneficial effects of potassium on human health. Physiol. Plant., 133: 725-735.
CrossRefDirect Link - Hotz, C. and R.S. Gibson, 2007. Traditional food-processing and preparation practices to enhance the bioavailability of micronutrients in plant-based diets. J. Nutr., 137: 1097-1100.
PubMed - Moraghan, J.T. and K. Grafton, 2001. Genetic diversity and mineral composition of common bean seed. J. Sci. Food Agric., 81: 404-408.
CrossRefDirect Link - Nchimbi-Msolla, S. and G.M. Tryphone, 2010. The effects of the environment on iron and zinc concentrations and performance of common bean (Phaseolus vulgaris L.) genotypes. Asian J. Plant Sci., 9: 455-462.
CrossRefDirect Link - Omenn, G.S., 1996. Micronutrients (vitamins and minerals) as cancer-preventive agents. IARC Sci. Publ., 139: 33-45.
PubMedDirect Link - Ortiz-Monasterio, I.J., N. Palacios-Rojas, E. Meng, K. Pixley, R. Trethowan and R.J. Pena, 2007. Enhancing the mineral and vitamin content of wheat and maize through plant breeding. J. Cereal Sci., 46: 293-307.
CrossRef - Ortolani, S. and S. Vai, 2006. Strontium ranelate: An increased bone quality leading to vertebral antifracture efficacy at all stages. Bone, 38: 19-22.
CrossRefPubMedDirect Link - Paredes, M.C., V.V. Becerra and J.U. Tay, 2009. Inorganic nutritional composition of common bean (Phaseolus vulgaris L.) genotypes race chile. Chil. J. Agric. Res., 69: 486-495.
Direct Link - Pasco, J.A., K.M. Sanders, M.J. Henry, G.C. Nicholson, E. Seeman and M.A. Kotowicz, 2000. Calcium intakes among Australian women: Geelong Osteoporosis Study. Aust. N. Z. J. Med., 30: 21-27.
CrossRefDirect Link - Pfeiffer, W.H. and B. McClafferty, 2007. HarvestPlus: Breeding crops for better nutrition. Crop Sci., 47: S88-S105.
CrossRefDirect Link - Phan-Thien, K.Y., G.C. Wright and N.A. Lee, 2010. Genotype-by-environment interaction affects the essential mineral composition of peanut (Arachis hypogaea L.) Kernels. J. Agric. Food Chem., 58: 9204-9213.
CrossRefDirect Link - Pinheiro, C., J.P. Baeta, A.M. Pereira, H. Domingues and C.P. Ricardo, 2010. Diversity of seed mineral composition of Phaseolus vulgaris L. germplasm. J. Food Compos. Anal., 23: 319-325.
CrossRefDirect Link - Rao, M.S.S., B.G. Mullinix, M. Rangappa, E. Cebert and A.S. Bhagsari et al., 2002. Genotype x environment interactions and yield stability of food-grade soybean genotypes. Agron. J., 94: 72-80.
Direct Link - Talukder, Z.I., E. Anderson, P.N. Miklas, M.W. Blair, J. Osorno, M. Dilwari and K.G. Hossain, 2010. Genetic diversity and selection of genotypes to enhance Zn and Fe content in common bean. Can. J. Plant Sci., 90: 49-60.
Direct Link - Welch, R.M. and R.D. Graham, 2004. Breeding for micronutrients in staple food crops from a human nutrition perspective. J. Exp. Bot., 55: 353-364.
CrossRefDirect Link - White, P.J. and M.R. Broadley, 2005. Biofortifying crops with essential mineral elements. Trends Plant Sci., 10: 586-593.
CrossRefDirect Link - Zhe, Y., J.G. Lauer, R. Borges and N. de Leon, 2010. Effects of genotype x environment interaction on agronomic traits in soybean. Crop Sci., 50: 696-702.
CrossRefDirect Link - Zimmermann, M.B. and R.F. Hurrell, 2007. Nutritional iron deficiency. Lancet, 370: 511-520.
CrossRefPubMedDirect Link - Zou, G., Y. Song, G. Zhai, Z. Zhang, J. Zou and Y. Tao, 2011. Genetic variability and correlation of stalk yield related traits and sugar concentration of stalk juice in a sweet sorghum (Sorghum bicolor L. Moench) population. Aust. J. Crop. Sci., 5: 1232-1238.
Direct Link